Article Text

Abstract
Aims: To investigate SOCS-2 (suppressor of cytokine signalling 2) protein expression in breast carcinoma samples in relation to biopathological parameters and survival.
Methods: A polyclonal antibody against SOCS-2 was used to study 50 archival breast carcinoma samples, collected from 1993 to 1995. The presence of SOCS-2 protein was investigated in relation to clinical and biological parameters used in breast cancer pathology. Fluorescence in situ hybridisation (FISH) was used to study whether SOCS-2 expression was related to SOCS-2 gene copy number.
Results: SOCS-2 protein was expressed in 34 of 50 breast carcinoma samples and was positively associated with low grade, low nuclear grade, and p27 protein. SOCS-2 expression was inversely related to Ki-67, cyclin A, retinoblastoma protein (pRb), and the epidermal growth factor receptor (EGFR). No relation with overall survival was demonstrated. SOCS-2 amplification was found in three samples. No relation between the number of FISH signals and SOCS-2 expression was found.
Conclusions: The significant correlation seen between SOCS-2 expression, grade, nuclear grade, p27, Ki-67, cyclin A, pRb, and EGFR labelling strongly supports the hypothesis that SOCS-2 loss might be related to cell proliferation and tumour growth in breast carcinoma. Gene copy number changes did not seem to play a role in SOCS-2 regulation and expression; other mechanisms might be involved and deserve further study.
- CIS, cytokine induced SRC homology 2 SH2 protein
- EGFR, epidermal growth factor receptor
- ER, oestrogen receptor
- FISH, fluorescence in situ hybridisation
- IHC, immunohistochemistry
- %LIa, percentage of the labelled nuclear area over the total neoplastic nuclear area
- OS, overall survival
- PR, progesterone receptor
- pRb, retinoblastoma protein
- SOCS, suppressor of cytokine signalling
- breast carcinoma
- fluorescence in situ hybridisation
- suppressor of cytokine signalling 2
- biopathological variables
- prognosis
Statistics from Altmetric.com
- CIS, cytokine induced SRC homology 2 SH2 protein
- EGFR, epidermal growth factor receptor
- ER, oestrogen receptor
- FISH, fluorescence in situ hybridisation
- IHC, immunohistochemistry
- %LIa, percentage of the labelled nuclear area over the total neoplastic nuclear area
- OS, overall survival
- PR, progesterone receptor
- pRb, retinoblastoma protein
- SOCS, suppressor of cytokine signalling
- breast carcinoma
- fluorescence in situ hybridisation
- suppressor of cytokine signalling 2
- biopathological variables
- prognosis
SOCS (suppressor of cytokine signalling) is a family of proteins involved in the regulation of cytokine responses to feedback inhibition of Janus kinase/signal transducer and activator of transcription signalling pathways.1–3 SOCS proteins act as a negative feedback loop: once produced, they bind to various components of the signalling apparatus and inhibit further signal transduction.1–3 There are eight SOCS proteins (SOCS-1–7 and the cytokine induced SRC homology 2 SH2 protein; CIS) that are thought to regulate cell proliferation and differentiation. SOCS proteins are variably expressed in different tissues, and are encoded by genes induced by exposure to cytokines, hormones, and growth factors.4 In addition to the negative feedback on cytokine signalling, the SOCS box motif (the C-terminal homology region of SOCS proteins) regulates protein turnover by targeting proteins for polyubiquitination and proteasome mediated degradation. Thus, the inhibitory actions of SOCS proteins seem to combine specific inhibitory interactions with a general mechanism of targeting associated signalling molecules for degradation.4 Various effects have been demonstrated in the immune system5 and immune pathology,6 whereas the role of SOCS proteins in other diseases is less well known. Several studies have shown that SOCS protein loss or inactivation has a growth promoting effect.7,8,9,10,11,12,13,14 Only two studies have been published about SOCS and breast carcinoma, and the results were conflicting. Raccurt and co-workers reported increased expression of SOCS-1–3 and CIS proteins in 17 in situ and infiltrating ductal carcinomas relative to normal breast tissue, but they found no consistent increases in SOCS transcripts in 10 breast carcinoma cell lines.15 In contrast, Sutherland and co-workers found promoter hypermethylation and silencing of SOCS-1 (but not of SOCS-2 and SOCS-3) in 9% of breast cancers and various degrees of SOCS transcription in several breast carcinoma cell lines.16
The conflicting data reported on SOCS proteins in breast cancer are the base of our present study, which was focused on SOCS-2 only. We consider SOCS-2 to be more interesting than other SOCS proteins because it is unique in having an oestrogen responsive sequence17 and can inhibit growth hormone under oestrogen stimulation.18 The importance and role of oestrogens in breast carcinoma development is well recognised: oestrogen receptor positive and negative breast cancers express different genes,19 show different characteristics, respond differently to treatment, and have a different clinical evolution.20
“The inhibitory actions of SOCS proteins seem to combine specific inhibitory interactions with a general mechanism of targeting associated signalling molecules for degradation”
We first studied whether SOCS-2 protein was detectable in 50 archival cases of breast carcinoma, because a limited number of breast carcinomas have been studied. We then compared the results with biopathological parameters and overall survival (OS). Because SOCS-2 protein loss was found to have a growth promoting effect,14 we studied proteins involved in cell cycle regulation or used as proliferation markers (cyclin D and A, retinoblastoma protein (pRb), p53, p21, p27, and Ki-67).21 We also used fluorescence in situ hybridisation (FISH) to study whether SOCS-2 presence/loss could be related to SOCS-2 gene copy number changes.
MATERIAL AND METHODS
Samples
Our study was conducted on a set of archival breast carcinoma samples obtained from 50 women (age range, 28–83 years) who underwent surgery for primary breast carcinoma at the S. Orsola University Hospital of Bologna, Italy, from 1993 to 1995. None of the patients received preoperative adjuvant chemotherapy. Each sample was divided into two fragments: one was fixed in buffered 4% formalin and embedded in paraffin wax. Serial sections of fixed tissues were cut and stained with haematoxylin and eosin for routine examination. Cases were classified according to World Health Organisation criteria22 and graded according to Elston and Ellis’s criteria.23 Nuclear grade was evaluated separately. All cases were staged according to the TNM criteria (UICC). A small part of each sample was also used for nuclear extraction and subsequent FISH analysis.
Immunohistochemistry (IHC) and image cytometry
The SOCS-2 protein was detected by a rabbit polyclonal antibody (H-74; Santa Cruz Biotechnology Inc, Santa Cruz, California, USA), diluted 1/400.
The following monoclonal antibodies were used: anti-oestrogen receptor (ER) clone 1D5, diluted 1/120; anti-progesterone receptor (PR) clone 1A6, diluted 1/65; anti-Ki-67 antigen clone MIB-1, diluted 1/100; anti-pRb clone G3-245, diluted 1/160; anti-p53 protein clone BP53.12.1, diluted 1/3000; anti-Her-2 clone C11, diluted 1/250 (all from BioGenex Laboratories, San Ramon, California, USA); anti-cyclin A clone 6E6, diluted 1/300; anti-cyclin D1 clone P2D11F11, diluted 1/200 (both from NovoCastra Laboratories, Newcastle upon Tyne, UK); anti-p21 protein clone EA10, diluted 1/450 (Oncogene, Boston, Massachusetts, USA); anti-p27 protein clone DCS-72.F6, diluted 1/190 (Neomarkers, Freemont, California, USA); and monoclonal anti-EGFR (epidermal growth factor receptor) clone 31G7, diluted 1/120, (Zymed, South San Francisco, California, USA).
Serial sections from formalin fixed, paraffin wax embedded tissue blocks were collected on 3-aminopropyltriethoxy-silane (Sigma Co, St Louis, USA) coated slides, dried overnight at 37°C, and processed for IHC according to a streptavidin–biotin–peroxidase preformed complex technique. Microwave pretreatment was performed with a citrate buffer solution (pH 6.0) for the antibodies to SOCS-2, ER, PR, Ki-67, cyclin A, p53, and p27, or with EDTA buffer solution (pH 8.0) for antibodies to Her-2, pRb, and cyclin D1. Sections were digested using proteinase K solution at room temperature before staining with anti-EGFR antibody.
Nuclear immunostaining was quantified by image cytometry with Cytometrica software (C & V, Bologna, Italy), as described previously.24 A labelling index was obtained for all the biopathological parameters evaluated by image cytometry and expressed as the percentage of the labelled nuclear area over the total neoplastic nuclear area (%LIa).
All the above mentioned parameters were categorised using the following cutoff values. SOCS-2 %LIa: < 5%, negative; ⩾ 5%, positive. ER, PR, and p21 %LIa: < 10%, negative; ⩾ 10%, positive. Ki-67, cyclin A, and EGFR %LIa: < 20%, negative, ⩾ 20%, positive. Her-2: < 25%, negative; ⩾ 25%, positive. p53 %LIa: < 4%, negative; ⩾ 4% to < 30%, intermediate; ⩾ 30%, positive. p27 %LIa: < 50%, negative; ⩾ 50%, positive. Cyclin D1 %LIa: < 5%, negative; ⩾ 5%, positive. pRb loss of expression was recorded as described previously25 and reported using two categories: 0, loss of function; 1, functional pRb protein.
FISH analysis
A bacterial artificial chromosome clone containing the genomic DNA sequence of the SOCS-2 gene (Homo sapiens chromosome 12 clone RP11-768A13) was obtained from BACPAC Resources (http://bacpac.chori.org/).
SOCS-2 DNA (1 μg) was labelled with biotin-16-dUTP (Roche Diagnostic, Basel, Switzerland) by nick translation (Gibco BRL Life Science Technologies, Gaithersburg, Maryland, USA), according to the supplier’s instructions.
Thirty seven of 50 samples collected from 1993 to 1995 and fixed in methanol/acetic acid were available for FISH analysis. Isolated nuclei had been treated with 0.075M KCl at 37°C for 10 minutes, fixed in 3/1 methanol/acetic acid, and stored at −20°C.
FISH was carried out as described previously,26 and the samples were evaluated on a Leitz microscope (Wetzlar, Germany).
Lymphocyte nuclei from a healthy donor were used as a control in all of the hybridisation experiments. Signals were counted in 100–200 nuclei, both in normal and in breast cancer samples, according to the criteria established by Hopman et al.27 To distinguish an abnormal number of signals from the background, cutoff values were set at 3 SD above the mean percentage of control nuclei showing 1 and 3 spots, as described previously.28
Statistical analysis
Differences in the distribution of categorical variables between groups were tested by Fisher’s exact test. OS was computed using the Kaplan–Meier test. Prognostic significance was evaluated using the Cox proportional hazards model. The analysis was conducted using StatView 5.0 statistical software (SAS Institute Inc, Cary, North Carolina, USA) and performed in October 2004.
RESULTS
SOCS-2 protein expression as detected by IHC
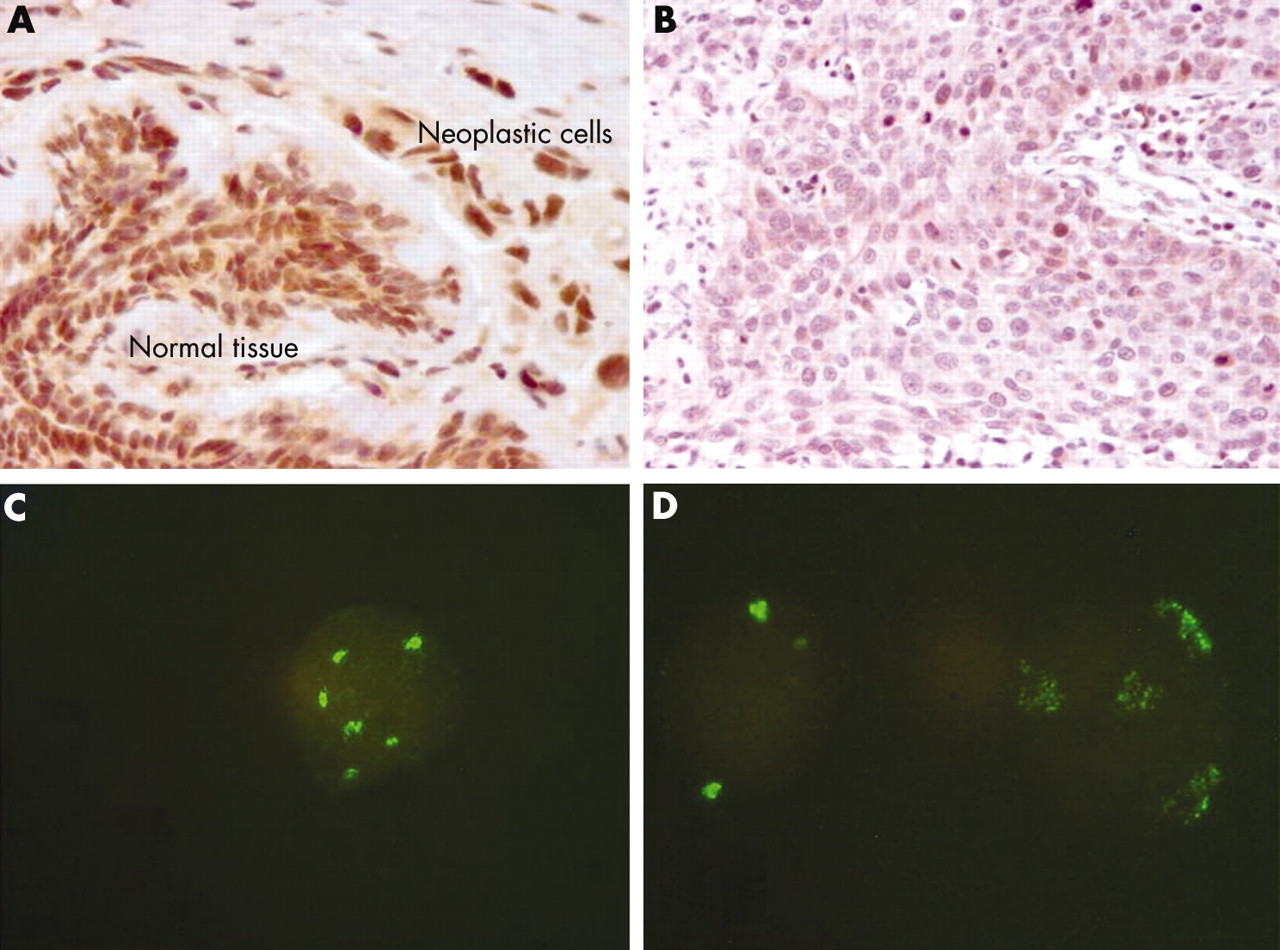
We analysed 50 breast carcinoma samples using a polyclonal antibody against a recombinant protein corresponding to amino acids 89–162, which maps within an internal region of the human SOCS-2 protein. Normal breast tissue was present in each section investigated by IHC and was compared with the tumour cells. Both the nuclei and cytoplasm were stained in normal mammary epithelial cells and in cancer cells. Endothelial and stromal cells were also stained. Weak to intense staining was detected in 34 breast carcinoma samples (fig 1A). In contrast, no or only a few stained tumour cells (below the 5% cutoff value described earlier) were found in 16 samples, whereas the adjacent normal breast tissue was positive (fig 1B).
{kind=link}
(A) Normal and tumour breast tissue showing intense suppressor of cytokine signalling 2 (SOCS-2) expression as detected by rabbit polyclonal antibody H-74 against the SOCS-2 protein. The nuclei are more intensively stained than the cytoplasm. (B) The SOCS-2 protein was detectable in only a few breast carcinoma cells, whereas normal tissue was positive for SOCS-2 protein. (C) In most samples investigated by immunohistochemistry, fluorescent in situ hybridisation analysis detected an increased number of signals for the SOCS-2 gene. A high copy number (gene amplification) was found in three samples: (D) shows a nucleus with a normal number of signals together with one showing SOCS-2 amplification.
Positive and negative samples were investigated with respect to the following pathological and biopathological parameters (table 1): size (T), grade and nuclear grade, lymph node invasion (N0/N+), ER, PR, Her-2, Ki-67, cyclin A, cyclin D1, p53, p21, p27, pRb, and EGFR. SOCS-2 expression was associated with low grade (p = 0.024), nuclear grade (p = 0.006), p27 expression (p = 0.002), and pRb expression (p = 0.0002), and was inversely correlated with Ki-67 labelling index (p = 0.009), cyclin A expression (p = 0.002), and EGFR (p = 0.049).
Relation between SOCS-2 expression as detected by IHC and biopathological variables
No relation to OS was found.
SOCS-2 alterations as detected by FISH
To investigate whether the SOCS-2 gene copy number was related to SOCS-2 protein expression we performed FISH using a bacterial artificial chromosome clone containing the whole sequence of the SOCS-2 gene. We used nuclei isolated at the time of surgery (1993–95), fixed in methanol/acetic acid, and stored at −20°C from 37 patients investigated for SOCS-2 expression by IHC in our present study. In each experiment control human lymphocyte nuclei were used. We counted two spots in a mean of 86.6 (SD, 4.8) control nuclei. One and three signals were counted in a mean of 12.3 (SD, 4.57) and 0.8 (SD, 1.75) control nuclei, respectively. FISH analysis was carried out successfully in 28 of 37 samples. Twenty two samples showed an increased number of signals (fig 1C) and 16 of the 22 also expressed the SOCS-2 protein. In three samples we detected clusters of high signal numbers, characteristic of high gene amplification (fig 1D). Samples expressing SOCS-2 had a greater number of nuclei with an abnormal number of signals (range, 12–81% v 6–36%) but the difference was not significant.
DISCUSSION
In our present study, we detected SOCS-2 protein expression in normal breast tissue and in most breast carcinoma samples. SOCS-2 expression was associated with high differentiation and a low proliferation rate, but not with OS. The SOCS-2 gene copy number was not related to SOCS-2 gene expression.
These results support the hypothesis that loss of SOCS-2 protein, and not its overexpression,15 may be a crucial step towards deregulation of the cell cycle, resulting in a growth promoting effect, as demonstrated by the inverse correlation between SOCS-2 and both cyclin A and Ki-67. Cyclin A is considered a marker of S phase: it is expressed in highly proliferating cancer and is related to poor outcome,29 as is Ki-67, another validated proliferation marker.21 We also found that SOCS-2 was directly correlated with p27 and functional pRb, and was inversely correlated with EGFR. These correlations suggest a loss of control of S phase entry, possibly related to SOCS-2 loss. p27 is thought to be a tumour suppressor gene, the loss of which promotes pRb phosphorylation and release of cyclin A/E. As a result, the cell can progress into S phase and duplicate. In mammary carcinoma cell lines, p27 concentrations increase when EGFR is reduced.30 Interestingly, both p27 and EGFR are regulated by the E3 ubiquitin ligase system, and are degraded after associating with the SOCS box. SOCS-1 and SOCS-3 can negatively regulate EGFR31 and the degradation of p27 occurs via the ubiquitin–ligase complex.4 We speculated that loss of SOCS-2 might deregulate both EGFR and p27 degradation, ultimately driving the cell to increased proliferation. Thus, SOCS-2 may act in a similar way to other SOCS proteins (1 and 3), which would explain the growth promoting effect as a consequence of SOCS inactivation/loss.
“Suppressor of cytokine signalling 2 expression was associated with high differentiation and a low proliferation rate, but not with overall survival”
Despite the fact that SOCS-2 contains an oestrogen responsive element, ER and SOCS-2 were not associated in our samples, and the hypothetical anti-proliferation effect of SOCS-2 on breast cancer cell did not appear to be oestrogen related. We can only conclude that SOCS-2 expression was not dependent on the presence of oestrogen, although we cannot exclude the possibility that SOCS-2 regulation could be different in ER positive and ER negative breast carcinomas.
We used IHC to investigate SOCS-2 protein because this protein has been investigated in only a limited number of invasive and in situ breast carcinoma samples, and has not been related to biopathological parameters,15 which are fundamental to define differences that may be important in breast cancer treatment and prognosis. Our results suggest that SOCS-2 has an effect on proliferation in breast carcinoma.
Using FISH, we showed that the SOCS-2 gene was amplified in breast carcinoma, although it was not correlated with protein expression as detected by IHC. These two techniques have often been used together to study genes known to be amplified.32 Both of them enable single cell analysis and the results can be compared. In agreement with other studies, which failed to find SOCS-2 regulation at the genetic level (promoter methylation, loss of heterozygosity), we suggest that transcriptional and post-transcriptional mechanisms may play a role in SOCS-2 regulation.
In conclusion, we suggest that, depending on its level of expression, the SOCS-2 protein may play a role in breast carcinoma cell proliferation and growth. SOCS-2 loss occurred in association with a high proliferation rate: SOCS-2 loss might represent one of the steps towards breast cancer progression, and deserves further attention.
Take home messages
-
The significant correlation between SOCS-2 (suppressor of cytokine signalling 2) expression, grade, nuclear grade, p27, Ki-67, cyclin A, retinoblastoma protein, and epidermal growth factor receptor labelling suggests that SOCS-2 loss is related to cell proliferation and tumour growth in breast carcinoma
-
Gene copy number changes did not play a role in SOCS-2 regulation and expression, so that other mechanisms might be involved and deserve further study
Acknowledgments
This work was supported by MURST, University of Bologna (funds for selected topics) and Pallotti Legacy for Cancer Research.