Article Text

Abstract
Objective Gastric cancer (GC) remains difficult to cure due to heterogeneity in a clinical challenge and the molecular mechanisms underlying this disease are complex and not completely understood. Accumulating evidence suggests that microRNAs (miRNAs) play an important role in GC, but the role of specific miRNAs involved in this disease remains elusive. We performed next generation sequencing (NGS)-based whole-transcriptome profiling to discover GC-specific miRNAs, followed by functional validation of results.
Design NGS-based miRNA profiles were generated in matched pairs of GCs and adjacent normal mucosa (NM). Quantitative RT-PCR validation of miR-29c expression was performed in 274 gastric tissues, which included two cohorts of matched GC and NM specimens. Functional validation of miR-29c and its gene targets was undertaken in cell lines, as well as K19-C2mE and K19-Wnt1/C2mE transgenic mice.
Results NGS analysis revealed four GC-specific miRNAs. Among these, miR-29c expression was significantly decreased in GC versus NM tissues (p<0.001). Ectopic expression of miR-29c mimics in GC cell lines resulted in reduced proliferation, adhesion, invasion and migration. High miR-29c expression suppressed xenograft tumour growth in nude mice. Direct interaction between miR-29c and its newly discovered target, ITGB1, was identified in cell lines and transgenic mice. MiR-29c expression demonstrated a stepwise decrease in wild type hyperplasia-dysplasia cascade in transgenic mice models of GC.
Conclusions MiR-29c acts as a tumour suppressor in GC by directly targeting ITGB1. Loss of miR-29c expression is an early event in the initiation of gastric carcinogenesis and may serve as a diagnostic and therapeutic biomarker for patients with GC.
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
-
Gastric cancer (GC) is a biologically heterogeneous disease accompanying various genetic and epigenetic alterations.
-
Several microRNAs (miRNAs) are frequently dysregulated in human cancers.
-
Most previous miRNA profiling studies were conducted using miRNA expression microarray platforms; however, conventional hybridisation-based microarray methodologies have several limitations including lack of sensitivity, narrow dynamic range and non-specific hybridisation.
What are the new findings?
-
This is the first study that performed next generation sequencing (NGS)-based whole-transcriptome profiling to identify GC-specific miRNA signatures.
-
We discovered the most promising GC-specific miRNAs, miR-29c, by comparison with three different high-throughput miRNA profilings generated from independent GC cohorts.
-
We elucidate a direct experimental evidence for the tumour suppressor role of miR-29c by regulating target gene (ITGB1) in GC through a series of in vitro and in vivo experiments.
-
We firstly demonstrated that suppression of miR-29c is an early event in gastric carcinogenesis using transgenic mouse models for gastritis (K19-C2mE) and GC (K19-Wnt1/C2mE).
How might it impact on clinical practice in the foreseeable future?
-
Our study provides GC-specific miRNA signatures generated by state-of-the-art NGS technique. Moreover, we highlight a potent tumour suppressor role of miR-29c through its downstream target gene, ITGB1, which may be an essential event in gastric carcinogenesis. Taken together, these results underscore the potential early diagnostic and therapeutic biomarker for patients with GC.
Introduction
Gastric cancer (GC) is the second leading cause of cancer-related death worldwide.1 Although clinical outcome of GC has gradually improved through earlier diagnosis, surgical resection and chemotherapy, 5-year survival rates of patients with GC are only 20%–30%.2 GC is a biologically heterogeneous disease that evolves in the background of various genetic and epigenetic alterations. Therefore, it is essential to have a more comprehensive understanding of molecular variables that affect GC disease pathways in order to develop appropriate approaches for its diagnosis and treatment.
Although a number of molecular drivers of GC have been described over the years,3 only very recently, microRNAs (miRNAs) have emerged as key players in the pathogenesis of this disease.4 miRNAs are short, non-coding RNA molecules that regulate gene expression by directly binding to the 3′UTR region of their target gene mRNA. miRNAs have been found to regulate a variety of cellular processes such as cell proliferation, differentiation, invasion, migration and epithelial–mesenchymal transition. Accumulating evidence indicates that miRNAs are frequently dysregulated in human cancers including GC,5–8 and that these non-coding RNAs play oncogenic or tumour suppressive roles in cancer cells. Thus, establishment of a miRNA expression profiles is important for investigating the underlying functional mechanisms for a specific cancer, and a better knowledge of their expression patterns could reveal molecular signatures that can be developed as prognostic/predictive biomarkers as well.
Most previous miRNA profiling studies were conducted using miRNA expression microarray platforms. Using this technology, an aberrant miRNA expression signature in GC tissues has been described previously.9 ,10 More recently, the emergence of next generation sequencing (NGS) platforms has revolutionised the field of genomic medicine, and helped in the identification of comprehensive, previously unrecognised and specific DNA and RNA targets in human cancers. In comparison with microarray platforms, NGS-based sequencing technologies have several advantages, including massive parallel analysis of widely expressed miRNAs in the genome, quantification of absolute abundance of miRNAs, identification of miRNA sequence variations and discovery of novel miRNAs. Although few recent studies have attempted transcriptomic profiling in GC,11 ,12 to the best of our knowledge, none of the previous studies have used NGS-based platform for the discovery and validation of GC-specific miRNAs by analysing matched tumour and non-cancerous gastric tissues.
In the present study, we performed comprehensive miRNA profiling using a NGS platform, identified several GC-specific miRNAs and discovered miR-29c expression to be significantly downregulated in GC tissues vis-à-vis matched normal tissues. We systematically validated the tumour suppressive role of miRNA-29c in a series of experiments performed in cell lines and transgenic mice models. Moreover, using cell lines and animal models, we identified that integrin β1 (ITGB1) is a novel, downstream gene target of miR-29c which plays an important role in cell signalling, differentiation, migration and apoptosis—all processes that are essential for the evolution and development of gastric carcinogenesis.
Materials and methods
Cell lines
Four human GC cell lines, SNU-601, SNU-668, AGS and MKN28, and one human cervical cancer cell line, HeLa, were obtained from the Korean Cell Line Bank (Seoul, Korea), and were cultured and maintained in appropriate culture conditions.
Tissue specimens
This study used 286 tissue specimens including 143 matched pairs of GC and corresponding normal mucosa (NM) tissues from three different GC patient cohorts, as described in online supplementary table S1. For NGS analysis, four matched pairs of frozen GCs and adjacent NM, and two additional NM specimens were obtained from Mie University Medical Hospital, Japan. For validation, 24 pairs of frozen GC and adjacent NM were obtained from Seoul National University Hospital, Korea. In addition, 113 pairs of formalin-fixed, paraffin-embedded (FFPE) GC tissues and matched corresponding normal gastric mucosa tissues from the Mie University Medical Hospital, Japan, were analysed. These studies were approved by the Institutional Review Boards of all involved institutions, and written informed consent was obtained from all patients.
Discovery of miR-29c using NGS
TruSeq miRNA libraries generated from GC and NM tissues were sequenced using an Illumina HiSeq 2000 sequencer with single end read length of 50 bases, following the manufacturer's instructions. The miRNA sequencing results were also compared with small RNA-seq data sets from the National Center for Biotechnology Information (NCBI) Sequence Read Archive (GSE36968)11 and miRNA microarray data sets from the GEO database (GSE28700).13
For the computational analysis of Illumina's small RNA-seq data, raw sequencing reads were subjected to quality filters as described previously.14 Before alignment, raw reads were initially filtered for (1) quality, (2) presence of the 3′ adapter, to ensure a small RNA was ligated and sequenced completely and (3) size of small RNA reads (17–27 nt). Alignment of reads was compared against human miRNA hairpin sequences in the miRBase V19 using Novoalign V2.08.01 (http://www.novocraft.com) with the following parameters: -m -r All 1 -l 18 -t 30 -h 90 -o SAM, default options. After alignment, the reads were further separated into two categories of mapped reads versus unmapped reads. For the mapped reads, we filtered out reads containing more than two mismatches.
For SOLiD small RNA reanalysis of the Profile#1,11 colour-space read alignments were performed using LifeScope Genomic Analysis Software V2.5 with the following parameters: workflow=small.rna, reference=hg19, smallRNA.genome.mapping=false and smallRNA.mirBase.mapping.scheme=18.2.0. After alignment, the filtering procedures, normalisation, log transformation and differential analyses were performed like the Illumina datasets.
After raw reads were mapped and filtered, an expression value (abundance) for each miRNA across all samples was calculated. For all differential expression analyses, the number of reads for a given miRNA was normalised by dividing the total number of mapped reads in that sample. This resulted in the per cent normalised read count for each miRNA, which was then multiplied by 106 to scale the expression value to an appropriate range. Principal component analysis was performed to identify possible outlier samples. Log transformation was applied to further scale the range of expression values. One-way analysis of variance (ANOVA) tests were performed between case and control groups in our cohort. For the Profile#1,11 tumours were compared by stage using normal tissues as control.
miRNA expression analysis
Expression of miR-29c was analysed using TaqMan miRNA assays (Applied Biosystems, Foster City, California, USA). The average expression level of miR-29c was normalised against U6 and RNU48 as described previously.6
For in situ hybridisation (ISH) analysis, 5 μm thick FFPE tissue sections were hybridised with the miR-29c probe (LNA-modified and 5′- and 3′-DIG-labelled oligonucleotide, Exiqon, Woburn, Massachusetts, USA) as described previously.6 Positive (U6 snRNA, Exiqon) and negative controls (scrambled miRNA control, Exiqon) were included in each hybridisation procedure as described previously.6
Gene expression analysis
Total RNAs were reverse transcribed to cDNA and quantitative real-time PCR (qRT-PCR) was performed as previously described.6 ,15 ,16 Primer sequences are described in online supplementary table S2.
Protein expression analysis
Proteins were isolated and western immunoblotting was performed using anti-ITGB1 (Abcam, Cambridge, UK), anti-α-tubulin (Sigma-Aldrich, St. Louis, Missouri, USA) and anti-β-actin (Cell Signaling Technology, Danvers, Massachusetts, USA) antibodies, as described previously.6 ,15
For immunocytochemistry, cells were fixed in 4% paraformaldehyde and incubated with 10% blocking buffer. Following blocking, cells were incubated with anti-ITGB1 (Abcam, Cambridge, UK) and Alexa Fluor 555 goat anti-rabbit IgG (H+L; Invitrogen, Carlsbad, California, USA).
Transfection experiments using miR-29c mimic, miR-29c inhibitor, stable miR-29c-expressing vector and ITGB1 siRNA
In order to transiently induce or inhibit miR-29c expression, hsa-miR-29c mimics (Applied Biosystems) or anti hsa-miR-29c inhibitor (Applied Biosystems) was used to transfect GC cells, as described previously.6 Verification of transfection efficiency was conducted using the Pre-miR miRNA Precursor Molecules Negative Control (Applied Biosystems) and Anti-miR miRNA Inhibitor Negative Control (Applied Biosystems), respectively.
To establish cell lines stably expressing miR-29c, a fragment containing the full-length coding region of miR-29c cDNA was amplified and cloned into the pcDNA3.1 vector.16 Plasmids were transfected into each cell line, and miRNA-expressing clones were selected as described previously.16 Primers for these steps are described in online supplementary table S2. To suppress ITGB1 expression, cells were transfected with either ITGB1 siRNA (Bioneer, Korea) or control scrambled siRNA (Bioneer) using Lipofectamine-2000 (Invitrogen), following the manufacturer's instructions.
Cell proliferation, adhesion, invasion and wound healing assays
Cell proliferation was measured using Cell Counting Kit-8 (Dojindo Laboratories, Kumamoto, Japan) following manufacturer's instructions. For the cell adhesion assay, 96-well plates were coated with fibronectin (10 μg/mL) at 4°C for 18 h and cells were allowed to adhere for 1.5 h at 37°C. At the end of this time period, adherent cells were quantified using the Cell Counting Kit-8 (Dojindo Laboratories, Kumamoto, Japan) following the manufacturer's instructions. Cell invasion and wound healing assays were performed as previously described.6
3′UTR luciferase reporter assays
ITGB1 3′UTR was amplified from human cDNA using primers. The PCR product was cloned into pGL3 luciferase reporter vector as described previously.17 Primers are shown in online supplementary table S2. Luciferase reporter vectors were transfected into the cells and luciferase activity was measured as described previously.6
Xenograft and transgenic mice models
To establish a tumour xenograft mice model, cancer cells stably expressing miR-29c were implanted into the flanks of 6-week-old female athymic nude mice (Balb/c nu; Orient Bio Inc., Seoul, Korea).16 Tumour size was measured and the volume was calculated at specified time intervals.16
Gastritis and GC animal models were developed using K19-C2mE [Tg(Krt19-Ptgs2,Krt19-Ptges)8Tko] and Gan (K19-Wnt1/C2mE) [Tg(Krt19-Wnt1)2Maos/Tg(Krt19-Ptgs2,Krt19-Ptges)8Tko] transgenic mice, as described previously.18 ,19 All transgenic mice models were based on C57BL/6 mouse. Additional experimental details are provided in online supplementary materials and methods.
All animal experiment protocols were approved by the Ethics Committees on Animal Experimentation of the Seoul National University and Kanazawa University.
Histology and immunostaining
Stomach tissues from mice were fixed in 4% paraformaldehyde, embedded and sectioned at 4 μm thickness. These sections were stained with H&E and anti-F4/80 (1:100, AbD Serotec, Oxford, UK), as described previously.18 ,19
Statistical analysis
Paired t test, Student t test, and Kruskal–Wallis tests were used to analyse miRNA and gene expression. The Spearman's correlation test was used to examine correlation between miRNA and target gene expression. Data are presented as mean±S.D and all statistical analyses were conducted using the Medcalc V12.3 (Broekstraat, Belgium) and the GraphPad Prism V5.0 (GraphPad Software, San Diego, California, USA).
Results
NGS-based discovery of miR-29c and validation of its expression pattern in GC tissues
To discover miRNA transcriptomes that distinguish GC from non-cancerous or normal gastric tissues, NGS of small RNAs was performed on four pairs of frozen GCs and adjacent normal tissues, plus two additional normal gastric mucosal specimens. NGS generated a total 22×106 reads (see online supplementary table S3). For all samples, the number of filtered reads for a given miRNA was normalised to the total mapped reads by scaling the expression values through logarithmic transformation. After filtering the low quality reads and 3′ adapter sequences, a total of 3,555,838 effective miRNA reads in GC tissues and 6,076,342 miRNA reads in the corresponding adjacent normal gastric mucosa tissues were obtained (see online supplementary table S3). Next, using one-way ANOVA test, 26 miRNAs were found to be differentially expressed between GC and NM tissues (p<0.05 with >2 fold change; figure 1A and see online supplementary table S4).

Discovery and validation of miR-29c transcriptome in gastric cancer (GC) tissues by next generation sequencing (NGS). (A) Differential expression of 26 GC-specific miRNAs between paired GC and normal mucosa (NM) tissues in NGS analysis. (B) Identification of four shared miRNAs (miR-29c, miR-135b, miR-148a and miR-204) between our data set (NGS profile) and two other data sets (Profiles#1 and #2). (C) Expression status of miR-29c in an independent validation cohort of 24 pairs of matching GC and NM tissues. ***p<0.0001, paired t test.
In order to further confirm the robustness of our GC-specific miRNA signature discovered by NGS, we compared our miRNA sequencing results (NGS profile) with two independent miRNA expression profiles that were generated from GCs using two different methods: a small RNA sequencing platform and a miRNA microarray. Kim et al11 reported small RNA-seq data using the SOLiD sequencing platform in 19 gastric tumour samples and six non-cancerous gastric tissues (Profile#1; see online supplementary table S5). In addition, Tseng et al13 generated miRNA expression profiles (Profile#2) using miRNA microarray in 22 paired tumour and non-tumour tissue specimens of GC patients. By comparing our NGS-based miRNA sequencing results with these two independent miRNA profiles, we identified four miRNAs that were shared between these datasets and were differentially expressed in GCs (miR-29c, miR-135b, miR-148a and miR-204; figure 1B and see online supplementary table S6).
Previously, ultra-deep miRNA sequencing profiles of normal stomach tissues revealed that miR-29c was one of most highly expressed miRNAs in normal tissues.20 Since miR-29c was among the four miRNAs that was significantly downregulated in GCs versus NMs, and was shared between our NGS profile and Profiles#1 and #2, we selected this miRNA for further validation and functional analyses in GC cell lines and animal models. In terms of validation, we quantified the expression of miR-29c in 24 pairs of frozen GCs and matched NMs, as well as in five GC cell lines. The qRT-PCR analyses revealed that miR-29c expression was significantly downregulated in GC tissues (p=0.0003; figure 1C) and cell lines, compared with NM tissues (figure 2A). These results suggest that miR-29c is consistently downregulated in GCs, and may serve as tumour suppressor in this disease.

In vitro functional analysis of miR-29c. (A) Expression of miR-29c in gastric cancer (GC) cell lines was analysed by real-time TaqMan PCR. Cell proliferation assay following transfection with either (B) miR-29c mimic or (C) anti miR-29c inhibitor. (D) Cell adhesion assay. Adherence ability of GC cell to fibronectin was analysed. NC, transfection of negative control; miR-29c, transfection of miR-29c mimic. (E) Cell invasion assays using Matrigel-coated transwell membrane. NC, transfection of negative control; miR-29c, transfection of miR-29c mimic. (F) Wound-healing assay. Cell monolayers were scratched with a pipette tip and images were taken 0 and 48 h after wound formation. NC, transfection of negative control; miR-29c, transfection of miR-29c mimic. *p<0.05, **p<0.01, ***p<0.001, t test.
Restoration of miR-29c expression inhibits cell proliferation, adhesion, invasion and migration in GC cells
To better understand the mechanistic role of miR-29c in gastric carcinogenesis, GC cell lines were transfected with either a miR-29c mimic or a miR-29c inhibitor. Since all five GC cell lines expressed miR-29c at very low levels, we selected SNU-601 for transfection experiments, as this cell line has been well characterised (figure 2A). Cell proliferation in miR-29c transfected cell lines was assayed by CCK-8 assay, each day, for up to 4 days. Restoration of miR-29c expression in GC cells resulted in decreased cell proliferation (figure 2B), whereas inhibition of miR-29c expression significantly increased GC cell proliferation compared with the negative controls (figure 2C).
Next, we analysed the effect of ectopic miR-29c expression on cellular adhesion, invasion and migration potential of SNU-601 cells. High miR-29c expression significantly reduced the adherence of GC cells to fibronectin, which is a major component of the extracellular matrix (p=0.0033; figure 2D). Similarly, overexpression of miR-29c significantly inhibited cell invasion into Matrigel-coated transwell membranes (p<0.001; figure 2E). In addition, high miR-29c expression significantly suppressed the ability of cells to migrate in a scratch wound healing assay (p<0.01; figure 2F). Collectively, these data suggest the tumour suppressive role of miR-29c in gastric carcinogenesis.
Overexpression of miR-29c reduces tumour growth in the xenograft nude mouse model, and therapeutic miR-29c delivery suppresses gastric tumourigenesis in nude mice
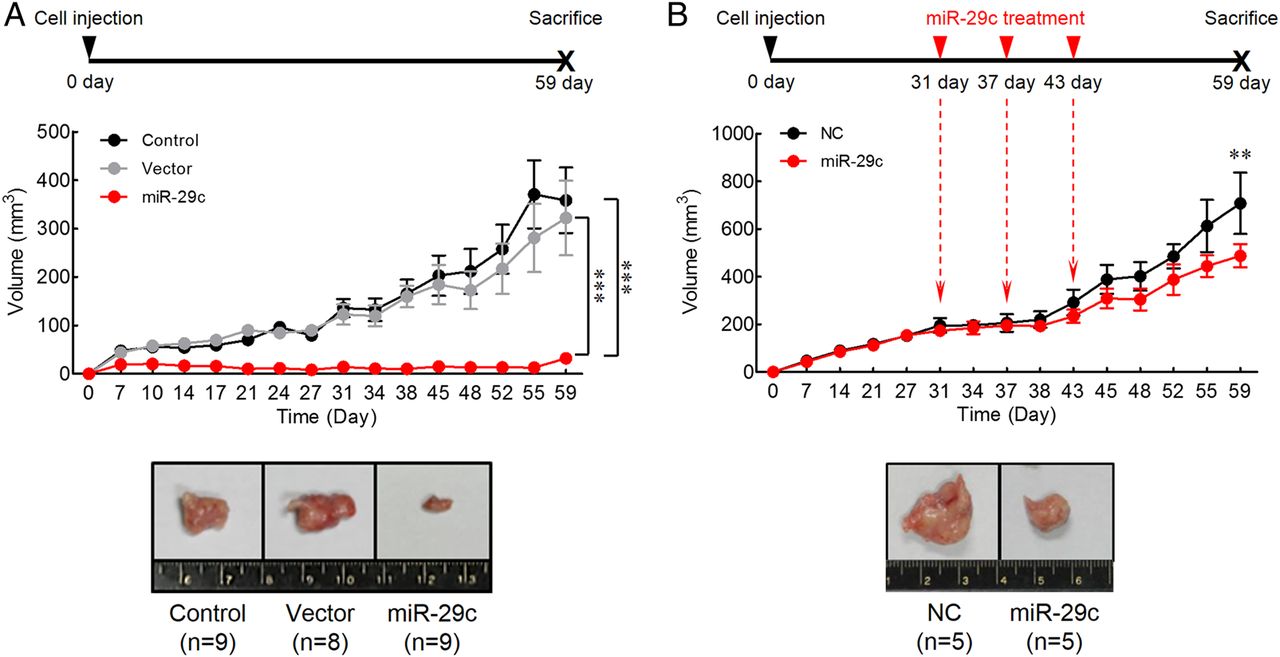
To ascertain the cellular mechanisms underlying miR-29c-mediated tumour suppression, we established a clone of SNU-601 cell line with stable overexpression of miR-29c, which was used for the xenograft nude mouse model experiments (figure 3A). The untreated parental SNU-601 cells and a stable clone of SNU-601 cells bearing the empty vector were used as controls. The mice implanted with miR-29c overexpressing cells revealed a significantly slower tumour growth compared with animals injected with either controls at the end of 8 weeks (miR-29c vs control and vector, respectively; p<0.0001).

In vivo gastric cancer tumourigenesis analysis in the xenograft nude mouse models. (A) Upregulation of miR-29c expression inhibited tumour growth in the xenograft nude mouse model. Control, SNU-601 parental cell line; Vector, SNU-601 cell line transfected with empty vector; miR-29c, SNU-601 cell line transfected with miR-29c expression vector. (B) Treatment of miR-29c suppressed xenograft nude mouse tumour. Either miR-29c mimic (miR-29c) or negative control (NC) was intratumourally injected using liposome at 31, 37 and 43 days after SNU-601 cell implantation. Tumour volumes represented as means±SD. **p<0.01, ***p<0.001, two-way ANOVA test.
To further examine the therapeutic effect of miR-29c in gastric tumourigenesis, we used a liposome-based delivery system for the delivery of miR-29c into gastric tumours (figure 3B). To establish gastric tumours in vivo, SNU-601 cells were implanted in 6-week-old Balb/c nude mice. At 31, 37 and 43 days after implantation, miR-29c mimic or negative control constructs were injected intratumourally using liposome-based vehicle. Mice treated with miR-29c mimics demonstrated a significant suppression of tumour growth, which started as early as after third treatment (43 day) compared with controls (p<0.01). Taken together, these results demonstrate that miR-29c expression strongly suppresses gastric tumour growth in vivo. Moreover, our miR-29c liposome-mediated delivery data also highlight the therapeutic potential of this non-coding RNA in gastric neoplasia.
MiR-29c acquires tumour suppressor abilities by directly targeting ITGB1 in GC
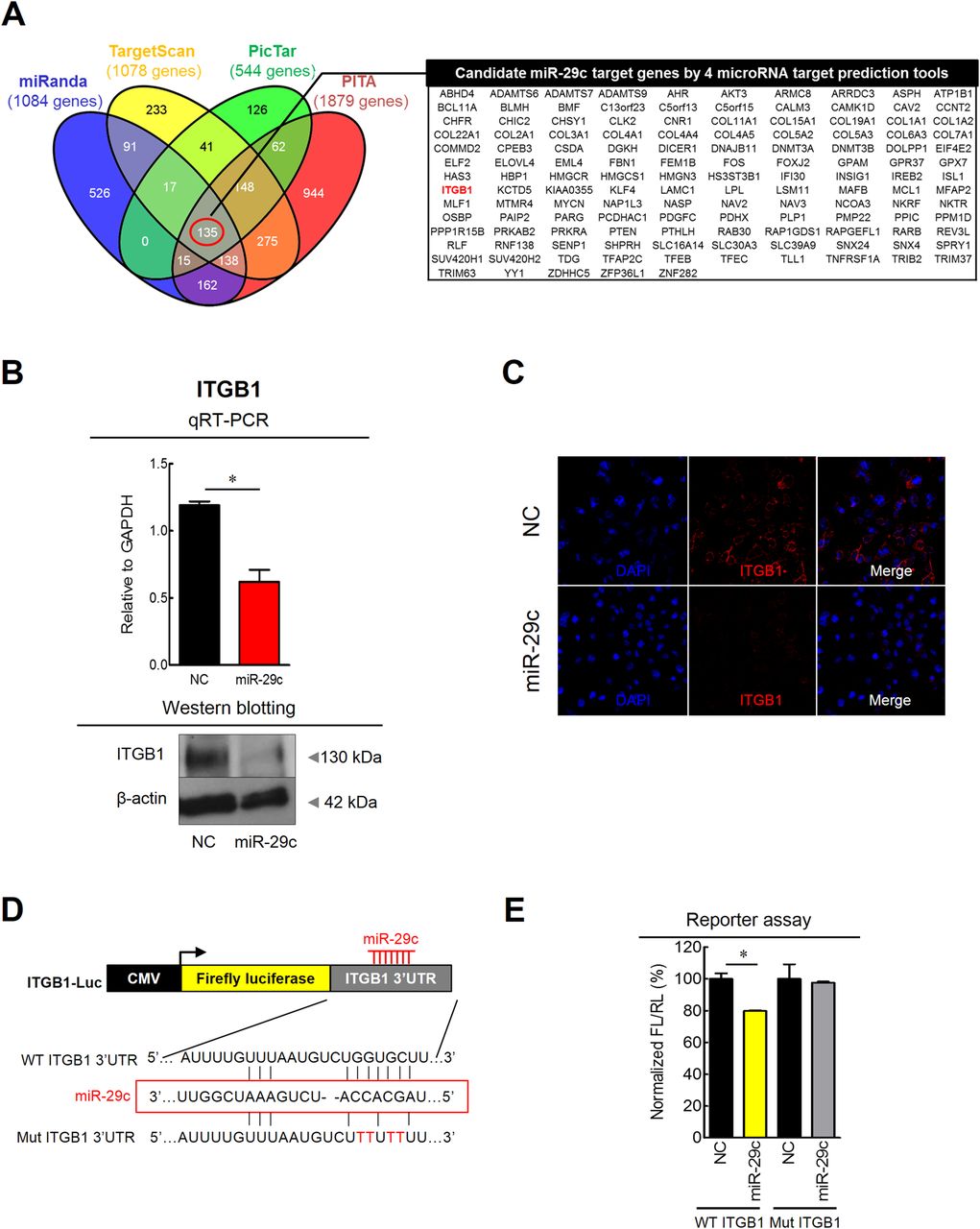
To gain further insight into the molecular mechanism(s) of miR-29c tumour suppressive activity, we sought out to determine its gene targets in the gastric mucosa by interrogating the interaction between miR-29c and its target mRNA transcripts. Candidate targets were first determined using target prediction engine including miRanda, TargetScan, PicTar and PITA (figure 4A). Although two miR-29c target genes were suggested previously, RCC221 and MCL1,22 we identified a novel target gene, ITGB1, as it seemed more germane to this study and has been shown to associate with prognosis and metastasis in patients with GC.23 ,24

Prediction and validation of miR-29c target gene in gastric cancer cell line. (A) 135 target genes which have miR-29c seed sites were predicted via four different miRNA target prediction tools (miRanda, TargetScan, PicTar and PITA). (B) Overexpression of miR-29c suppressed mRNA (qRT-PCR) and protein expression (western blotting) of ITGB1. (C) Immunocytochemistry for ITGB1 showed low ITGB1 fluorescence intensity following miR-29c treatment. (D and E) Luciferase reporter assays revealed direct binding of miR-29c to the wild type (WT), but not the mutant (Mut) sequences within the 3′UTR regions of TGB1. NC, transfection of negative control; miR-29c, transfection of miR-29c mimic; FL, firefly luciferase; RL, Renilla luciferase. *p<0.05, t test.
To further confirm the functional interaction between miR-29c and ITGB1 generated by the target prediction algorithms, we performed a series of assays to determine the relationship between the ITGB1 and miR-29c in GC cell lines. Overexpression of miR-29c by transfecting miR mimics in the SNU-601 cells resulted in significant reduction in ITGB1 mRNA transcription as well as protein expression by western blotting (figure 4B). Moreover, immunocytochemical analysis revealed decreased ITGB1 fluorescence intensity following transfection with miR-29c mimics (figure 4C). Since miRNAs are known to affect translation of gene transcripts, we performed luciferase reporter assays to determine whether miR-29c directly interacts with the 3'UTR of ITGB1. GC cells were cotransfected with various combinations of pGL3 luciferase reporter vectors. The plasmids were transfected with empty luciferase vector, luciferase vector containing wild type ITGB1-3′UTR or luciferase vector containing mutant-type ITGB1-3′UTR and miR-29c mimic or a negative control. In cells transfected with the wild type ITGB1-3′UTR and the miR-29c mimic, a significant decrease in luciferase activity was observed compared with wild type ITGB1-3′UTR vector and negative controls (figure 4D, E). However, the luciferase activity of mutant-type ITGB1-3′UTR vector did not change following cotransfection with the miR-29c mimic, indicating miR-29c directly interacts with the 3′UTR of ITGB1 to mediate its tumour suppressive function.
Since the functional role of the ITGB1 gene has not been previously studied in the pathogenesis of GC, we examined whether miR-29c regulation of ITGB1 translation actually influences the phenotype of gastric mucosa. To investigate the potential role of ITGB1 in GC pathogenesis, we performed ITGB1 knockdown in SNU-601 cells by transfecting them with either siRNA or the negative control. The knockdown effect by ITGB1 siRNA was confirmed by qRT-PCR as well as western blotting (figure 5A). Suppression of ITGB1 significantly reduced the ability of GC cells to adhere to fibronectin (figure 5B), and reduced ITGB1 expression led to significant decrease in their invasive and migratory abilities (figure 5C, D).

In vitro functional analysis and expression of ITGB1 in gastric cancer (GC). (A) Treatment of si-ITGB1 diminished mRNA (qRT-PCR) and protein expression (western blotting) of ITGB1 in GC cell line. Downregulation of ITGB1 by transfection with si-ITGB1 suppressed GC cell; NC, transfection of negative control, (B) adherence ability to fibronectin; NC, transfection of negative control, (C) invasion ability into matrigel-coated transwell membrane, NC, transfection of negative control, and (D) cell migration ability in a scratch wound healing assay; NC, transfection of negative control. (E) Expression status of ITGB1 in human GC tissues (GC) and corresponding normal mucosa (NM) tissues. GAPDH, glyceraldehyde-3-phosphate dehydrogenase. (F) Correlation analysis between miR-29c and target gene (ITGB1) expression in GC tissues. Red line represents linear regression line. *p<0.05, **p<0.01, ***p<0.001, t test.
Next, ITGB1 expression was measured in 24 pairs of GC and NM tissues using qRT-PCR. The expression of ITGB1 was significantly upregulated in GC compared with corresponding NM tissues (p=0.0024; figure 5E). More interestingly, ITGB1 expression was significantly and inversely correlated with miR-29c expression in the same patient cohort (r=−0.43, p=0.002; figure 5F). Collectively, these findings highlight that miR-29c directly inhibits ITGB1 gene expression, which in turn prevents the growth proliferation programme in the gastric cells.
Suppression of miR-29c is an early event in gastric carcinogenesis
To investigate the clinical relevance of miR-29c expression in GC, we analysed miR-29c expression in an independent cohort of 113 matched pairs of GC and corresponding NM tissues (the Clinical validation cohort). Consistent with our NGS validation cohort specimens, miR-29c expression was significantly lower in GC compared with NM (p<0.0001; figure 6A). These results were further confirmed by ISH studies, which also revealed marked suppression of miR-29c expression in GC compared with NM tissues (figure 6B).
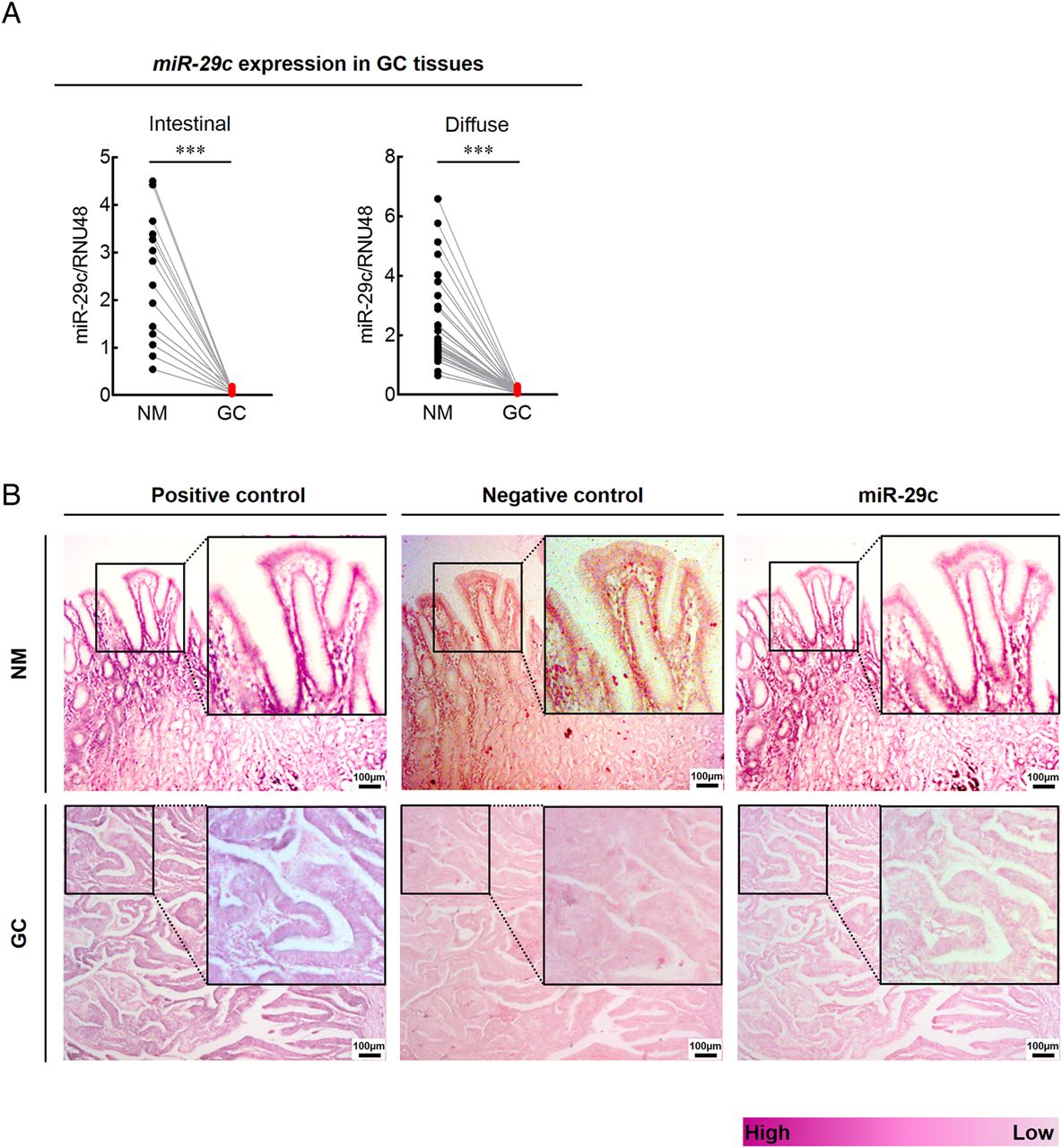
Clinical relevance and in situ hybridisation expression of miR-29c in gastric cancer (GC) tissues. (A) Expression status of miR-29c in pairs of matching GC and normal mucosa (NM) tissues (left panel, intestinal type GC; right panel, diffuse type GC; ***p<0.001, t test). (B) In situ hybridisation analysis of miR-29c in matching GC and NM tissues (positive control, U6 snRNA, purple colour; negative control, scrambled miRNA control, pink colour; inserts show higher magnifications).
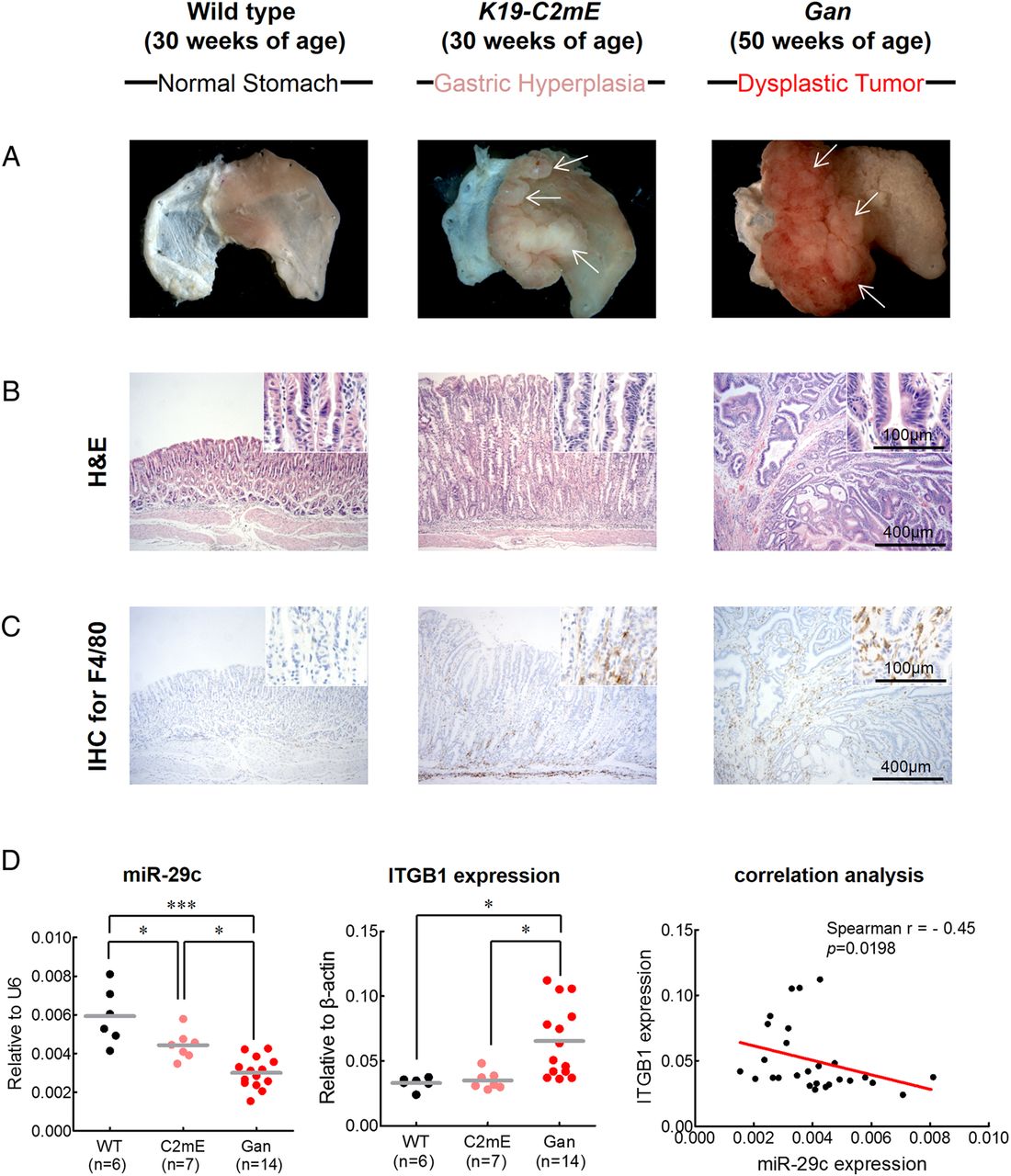
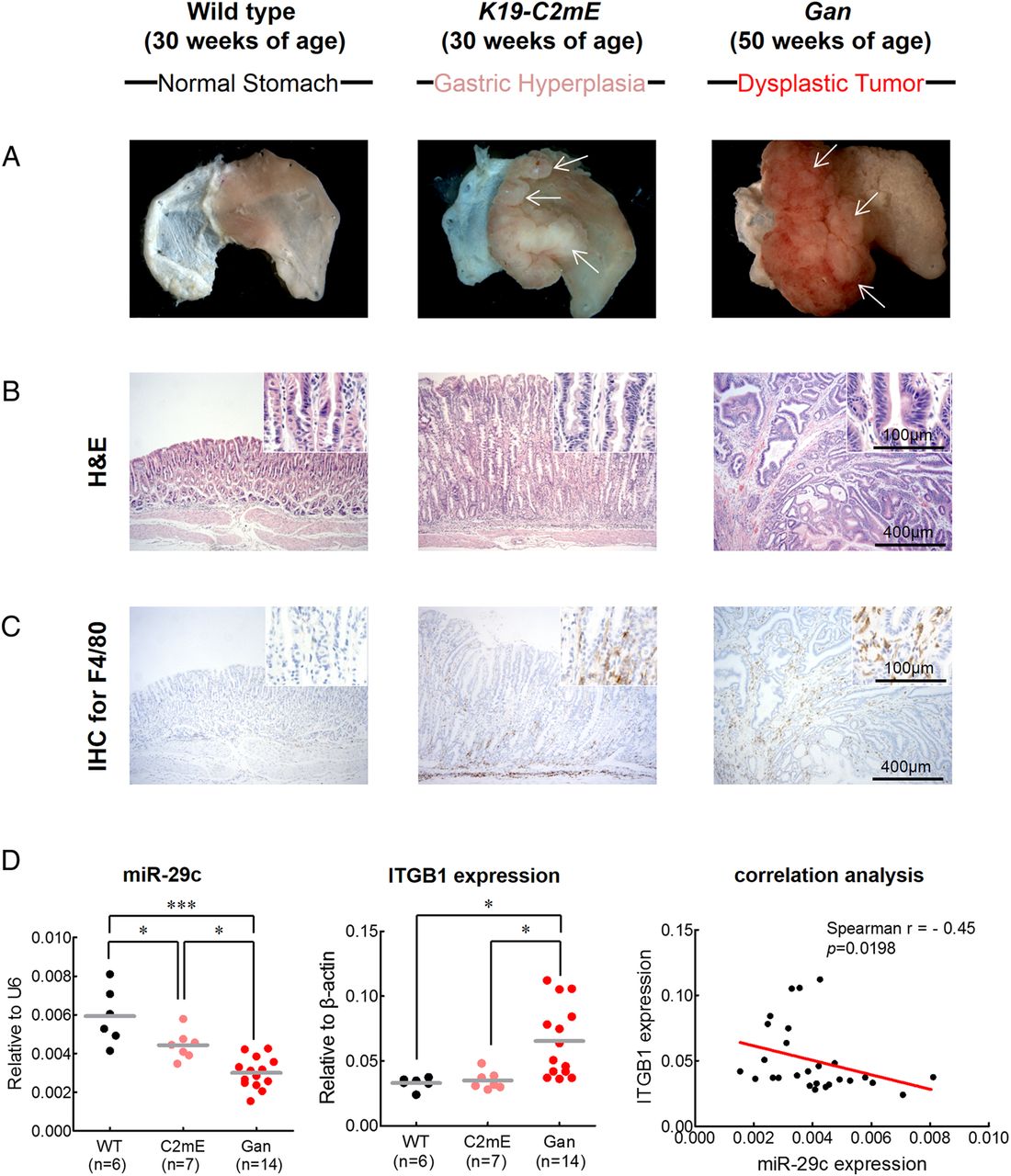
Based on the results to date, we hypothesised that miR-29c may affect the initiation of gastric carcinogenesis. To address this issue, we established two different transgenic animal models: a K19-C2mE mouse model for gastritis and a Gan mouse model for GC. Previously, we demonstrated that K19-C2mE mice develop gastric hyperplasia in the stomach via induction of the COX-2/PGE2 pathway and Gan mice acquire inflammation-associated GC via induction of COX-2/PGE2 and Wnt signalling pathways.18 ,19 We found that the K19-C2mE mice developed mucosal hyperplasia in the proximal glandular stomach at 30 weeks of age, while Gan mice developed dysplastic gastric tumours (figure 7A, B) at 50 weeks of age. Abundant activated macrophages are accumulated in the gastric mucosa of the transgenic mice, which promote proliferation of epithelial progenitor cells through Wnt signalling.25–27 Thus, to estimate gastric tumourigenesis process, we analysed macrophage infiltration in these mice by immunohistochemistry for F4/80, the best known marker for mature mouse macrophages28 (figure 7C). The wild type mouse displayed rare macrophage infiltration in glandular stomach, whereas abundant macrophage infiltrates were observed in mucosal stroma of the K19-C2mE mice and in the Gan mice bearing gastric tumours.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Functional analysis of miR-29c in gastritis (K19-C2mE mice) and gastric tumour (Gan mice) transgenic mice models. Gastritis in K19-C2mE mice and gastric tumours in K19-Wnt1/C2mE mice. (A) Macroscopic photographs and (B) H&E staining of the glandular stomach of the wild type (left), K19-C2mE (middle) and Gan (right) mice. White arrows in middle panel of (A) indicate gastric hyperplasia lesions. White arrows in right panel of (A) indicate gastric dysplastic gastric tumours. (C) Immunohistochemistry (IHC) staining for macrophages using F4/80 antibody in the wild type mouse stomach (left), K19-C2mE (middle) and Gan (right) mice. (D) Expression status of miR-29c (left) and ITGB1 (middle), and correlation analysis (right) between miR-29c and target gene (ITGB1) expression in transgenic mice models. Red line represents linear regression line. *p<0.05, ***p<0.001, one-way ANOVA test.
Finally, we analysed the expression of miR-29c and its target gene, ITGB1, in these two transgenic mice models. Compared with the wild type animals, both K19-C2mE and the Gan mice showed significant decrease in miR-29c expression (p<0.0001; figure 7D). Furthermore, miR-29c expression were significantly lower in the gastric tumours of the Gan mice compared with the K19-C2mE mice with gastric hyperplasia (p=0.0011). By contrast, ITGB1 expression was significantly increased in the Gan mice compared with the C2mE and wild type animals, respectively. Moreover, ITGB1 expression was significantly and inversely correlated with miR-29c expression in both transgenic mice models (r=−0.45, p=0.0198). These results provide novel and important evidence for the functional role of miR-29c in gastric mucosa and suggest that loss of miR-29c expression in the precancerous state may facilitate initiation of gastric carcinogenesis.
Discussion
In this study, we first performed state-of-the-art NGS in matched pairs of human GCs and NM to identify miR-29c as a GC-specific miRNA. Thereafter, using a series of in vitro and in vivo assays, we uncovered that miR-29c act as an important tumour suppressor in the normal gastric mucosa. Although decreased miR-29c expression has been reported previously in human cancers,21 ,22 ,29 ours is the first study that provides a novel and comprehensive insight into the functional role of miR-29c as it relates to the pathogenesis of GC. To the best of our knowledge, this is the first study that has: (i) used NGS to identify GC-specific miRNA signatures, (ii) used a series of cell culture and animal models to uncover the tumour suppressive role of miR-29c in gastric epithelium, (iii) provided promising evidence for the potential therapeutic use of miR-29c in GC, (iv) identified ITGB1 as a novel target gene of miR-29c and (v) demonstrated that suppression of miR-29c in transgenic animal models prevented gastric tumour growth.
Although a few previous studies have attempted NGS for the development of genomic signatures in GC, the primary goal of our study was to focus on GC-specific miRNA signatures. Using the Illumina HiSeq 2000 platform, deep sequencing was performed to generate whole-transcriptome profiles of miRNAs in paired GC and corresponding NM tissues. To further ensure the reliability and reproducibility of our discovery data set and to narrow down the most promising candidate miRNAs, we compared our results with two other data sets that were generated using two different high-throughput profiling techniques (SOLiD sequencing platform and miRNA microarray platform) in independent GC cohorts. A total of four miRNAs (miR-29c, miR-135b, miR-148a and miR-204) were common between these three independent GC miRNA profiles. Among these, compared with the cancerous tissue, high levels of miR-29c expression were observed in normal gastric tissues, and these results were confirmed by ultra-deep sequencing. In additional data analysis using a different tool for miRNA deep sequencing analysis, miRNAkey,30 miR-29c was confirmed as a prominent candidate miRNA dysregulated in GC (see online supplementary figure S1). This consistent finding from these independent cohorts as well as different analysis tools was in part the rationale for selection and systematic exploration for the role of miR-29c in gastric neoplasia.
Ours is the first study that used a series of in vitro and in vivo experiments to provide a direct experimental evidence for the functional role of miR-29c as a tumour suppressor in gastric neoplasia. In addition, we identified ITGB1 as a novel miR-29c-target gene, and confirmed its direct interaction with the miR-29c. In terms of miR-29c target genes, RCC221 and MCL122 have been previously reported, and we were also able to confirm a direct interaction of the two target genes (RCC2 and MCL1) with miR-29c using luciferase reporter assays (see online supplementary figure S2). The expression status of these two genes did not have a direct correlation with miR-29c expression in GC tissues (see online supplementary figure S3), while high levels of ITGB1 expression have been shown to associate with poor prognosis and recurrence in patients with GC.23 ,24 In addition, previous studies have shown the interaction of ITGB1 with cytokines, growth factors and extracellular matrix proteins.31 ,32 Overexpression of ITGB1 has been found in various epithelial malignancies including breast cancer and glioblastoma, during invasion, angiogenesis and metastasis.33–35 Intriguingly, our mechanistic and functional data permit us to better appreciate the functional role of ITGB1 in human cancers; its expression positively regulated GC cell adhesion, invasion and migration. Furthermore, our observation for an inverse correlation between miR-29c expression and ITGB1 expression in GC tissues fills this important void in literature for the missing experimental evidence for the function of miR-29c and ITGB1 in gastric pathogenesis.
Another interesting aspect of our study is that we first demonstrated the tumour initiator role of miR-29c using transgenic mice models of gastric hyperplasia and carcinogenesis. Our data revealed that while miR-29c was significantly downregulated in a stepwise manner in wild type-hyperplasia-dysplasia cascade, the expression of its target gene, ITGB1, was conversely upregulated in these animal models of gastric carcinogenesis. Collectively, our findings provide a novel mechanistic insight that was previously unrecognised, and highlight that suppression of miR-29c is an early phenomenon in gastric carcinogenesis and may trigger the initiation of GC.
Our study also highlighted the potential clinical application of miR-29c in the diagnosis and treatment of patients with GC. Although miR-29c expression status was not associated with prognosis and metastasis in our patient cohort, suppression of miR-29c was an early event in gastric carcinogenesis, suggesting the possibility for the application of miR-29c as a diagnostic biomarker for this malignancy. With regard to its therapeutic potential, we noted that treatment of miR-29c mimics markedly reduced tumour volume in GC xenograft mouse model. In addition, a recent study showed restoration of miR-29c expression by treatment of celecoxib, a selective COX-2 inhibitor, in human GC cell lines.22 In this context, our observation that miR-29c was significantly suppressed in gastritis and gastric tumour mice models through genetic induction of COX-2/mPGES-1 and Wnt/PGE2 pathways suggests the importance of miR-29c as a potential therapeutic target for COX-2 meditated gastric carcinogenesis.
In conclusion, this study provides discovery and validation of GC-specific miRNA transcriptome profiles generated by NGS. We identified that miR-29c is a potent tumour suppressor in the stomach, and its growth inhibitory effects are, in part, mediated through its downstream target gene, ITGB1. Using cell culture and animal models, functional characterisation for the role of miR-29c reveals that loss of its expression is an early event in gastric carcinogenesis.
Acknowledgments
We thank Dr Margaret M Hinshelwood for her skilful editing and revision of the manuscript.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
- Data supplement 2 - Online figures
Footnotes
-
T-SH, KH, GX contributed equally.
-
Contributors Study concept and design: T-SH, KH, ANC, AG, VNK and H-KY; provision of samples: YO, YT, HO, MO, H-JL and H-KY; acquisition of data: T-SH, KH, ANC, BC, HO and MO; analysis and interpretation of data: T-SH, KH, ANC, GX, VNK, AG and H-KY; statistical analysis: T-SH, KH, ANC and GX; drafting of the manuscript: T-SH, KH, ANC, GX, AG and H-KY.
-
Funding This study was supported by A3 Foresight programme from National Research Foundation of Korea.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval Institutional Review Boards.
-
Provenance and peer review Not commissioned; externally peer reviewed.