


Abstract
Background/Aim: Intraoperative electron radiation therapy (IOERT) is a therapeutic approach that delivers a single high dose of ionizing radiation (IR) directly to the tumor bed during cancer surgery. The main goal of IOERT is to counteract tumor growth by acting on residual cancer cells as well as to preserve healthy surrounding tissue from the side-effects of radiation therapy. The radiobiology of the healthy tissue response to IR is a topic of interest which may contribute to avoiding impairment of normal tissue and organ function and to reducing the risks of secondary cancer. The purpose of the study was to highlight cell and gene expression responses following IOERT treatment in the human non-tumorigenic MCF10A cell line in order to find new potential biomarkers of radiosensitivity/radioresistance.
Material and Methods: Gene-expression profiling of MCF10A cells treated with 9 and 23 Gy doses (IOERT boost and exclusive treatment, respectively), was performed by whole-genome cDNA microarrays. Real-time quantitative reverse transcription (qRT-PCR), immunofluorescence and immunoblot experiments were carried out to validate candidate IOERT biomarkers. Clonogenic tests and morphological evaluations to examine cellular effects induced by radiation were also conducted. Results: The study revealed a dose-dependent gene-expression profile and specific key genes that may be proposed as novel markers of radiosensitivity. Our results show consistent differences in non-tumorigenic cell tolerance and in the molecular response of MCF10A cells to different IOERTs. In particular, after 9 Gy of exposure, the selection of a radioresistant cell fraction was observed. Conclusion: The possibility of clarifying the molecular strategies adopted by cells in choosing between death or survival after IR-induced damage opens-up new avenues for the selection of a proper personalized therapy schedule.
- Intraoperative electron radiation therapy
- IOERT
- ionizing radiation
- IR
- MCF10A cells
- microarray
- gene-expression profiling
Intraoperative electron radiation therapy (IOERT) differs from conventional radiotherapy (RT), since a large dose of ionizing radiation (IR) is employed in a single fraction directly to the tumor bed during cancer surgery, either as an exclusive treatment of 21-23 Gy or as an advanced boost of 9-12 Gy. The use of IOERT for breast cancer (BC) treatment has increased due to the development of the partial breast irradiation (PBI) strategy with the intent of avoiding tumor recurrence. This segmental RT replaces whole-breast irradiation and is based on the discovery that approximately 85% of local relapses are localized to the same breast quadrant from which the primary tumor was removed (1-6). Preliminary results of PBI with IOERT, both as a boost and as an exclusive treatment, seem to be promising in terms of local disease control, however few data have been collected on long-term toxicity, as well as molecular stress mechanisms specifically induced by high-dose treatments (7-9). IR, both as X-rays, mainly used in conventional external-beam RT, and high-energy electrons generated by linear accelerators in IOERT, cause cell injury to both tumor and normal cells, producing a disequilibrium in the survival/cell death decision (9-11). Increasing evidence suggests that different factors, including the type of radiation and dose, are primarily important in radiation response, related also to the cell type. However, factors establishing the specific cellular fate after IR exposure have not been clearly defined. In addition, it has been shown that cell death induction is a very complex mechanism accounting for the different effects of IR, and cell death modality is not unique in response to radiation in cancer and normal cells (11-13). Several IR-induced genes trigger complex intracellular signaling pathways controlling many processes, such as cell-cycle progression, survival and cell death, DNA repair and inflammation (13-15). Nevertheless, the contribution of these genes and regulatory networks involved in the cellular response to high radiation doses is not completely understood. Despite the great interest of the scientific community on the clinical application of IOERT in BC, very few studies describe the effects of IOERT and particularly the molecular mechanisms of radiation toxicity in normal breast tissue. The gene-expression profiles of non-tumorigenic breast cells treated with high IR doses, such as those used during IOERT need to be further evaluated (16-17). It should also be taken into account that BC is a heterogeneous and complex disease at both molecular and clinical levels. Thus, on the one hand, the failure of radiation treatments associated with cell radioresistance may occur and on the other, effects due to radiosensitivity of the normal tissue surrounding the tumor may be present (18-21).
Recently, we investigated cell response and gene-expression profiles activated by IOERT treatments in the human MCF7 BC cell line and designed two network models induced by 9 and 23 Gy doses using the selected and validated genes. We reported a dose-dependent transcriptome change that regulates cell-fate decisions in two different ways, according to the doses used. IOERT treatments inhibited the growth and proliferation of MCF7 cells, and the post-irradiation cell traits reflected a typical senescent phenotype (22).
In order to assess the toxic effects of IOERT treatment and to select for potential new biomarkers of radiosensitivity/radioresistance, we describe cell and gene-expression responses of the non-tumorigenic mammary MCF10A cell line, used as a model of normal breast epithelial cells, following exposure to 9 and 23 Gy doses.
Understanding the molecular mechanisms of radiation toxicity is critical for the development of counter-measures for radiation exposure, as well as for improvement of clinical radiation effects in cancer treatment.
Materials and Methods
IOERT treatment. The NOVAC7 (Sortina Iort Technologies, Vicenza, Italy) IOERT system was used to perform treatments at different tissue depths. The beam collimation was performed through a set of polymethylmethacrylate applicators: cylindrical tubes with a diameter ranging from 3 to 10 cm and face angle of 0°-45°. The electron accelerator system was calibrated under reference conditions, cell-irradiation setup and the dose distribution were conducted as previously reported (22). IOERT cell treatments were carried out at two dose values, 9 Gy (in boost scheme) and 23 Gy (according to the exclusive modality) to the 100% isodose at a dose rate of 3.2 cGy/pulse.
Cell culture and clonogenic survival assay. The human non-tumorigenic breast epithelial MCF10A cell line was purchased from the American Type Culture Collection (Manassas, VA, USA) and cultured at 37°C in an incubator with 5% CO2 according to the supplier's instructions. All cell culture media and supplements were obtained from Invitrogen (Carlsbad, CA, USA). Cells were seeded in 100-mm Petri dishes or in 24-well plates 48 h before treatments and at sub-confluence were irradiated. Twenty-four hours after irradiation, clonogenic survival assay was performed according to the protocol of Franken et al. (23), as previously described (22). The surviving fraction (SF) of irradiated cells was normalized to the plating efficiency (PE) of untreated control cells (basal). Data represent the average SF±standard deviation (SD) of three biologically independent experiments. Moreover, in order to evaluate cell radiation effects, cells throughout the course of the assays were monitored for cell morphology and growth pattern by photographing five random fields for each treatment under a Zeiss Axiovert phase-contrast microscope (Carl Zeiss, Göttingen, Germany).
Gamma-H2AX immunofluorescence analysis. Cells were grown on glass coverslips to reach 70% confluency before treatment and control cells (basal, i.e. untreated) were grown in parallel. After defined times, cells were processed for immunofluorescence, as previously described (22). The images were acquired by a Nikon Eclipse 80i (Chiyoda, Tokyo, Japan). γH2AX quantification was performed by ImageJ analysis software (http://rsb.info.nih.gov/ij/).
Whole-genome cDNA microarray expression analysis. Gene expression profiling of MCF10A cells treated with 9 and 23 Gy IR doses were performed. Twenty-four hours after each treatment, MCF10A cells were harvested, counted and the pellet stored immediately at −80°C. RNA extraction, quantification and purity evaluation were performed as previously described (22). Gene-expression profiles of MCF10A cells treated with 9 Gy and 23 Gy were carried-out according to Agilent Two-Color Microarray-Based Gene Expression Analysis protocol as described recently by our group (22). Seven replicates were performed. Statistical data analysis, background correction, normalization and summary of expression measure were conducted with GeneSpring GX 10.0.2 software (Agilent Technologies) as previously reported (22). Genes were identified as being differentially expressed if they showed a fold-change (FC) of at least 1.5 and a p-value for the difference of less than 0.05 compared to untreated MCF10A cells used as reference sample. The data discussed here have been deposited in Gene Expression Omnibus of National Center for Biotechnology Information (NCBI) link (24) and are accessible through GEO Series accession number (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE65954). Microarray data are available in compliance with Minimum Information About a Microarray Experiment standards.
Primer sequences used for Real-Time Quantitative Reverse Transcription PCR analyses.
MetaCore network analyses. Gene expression profiles of IOERT-treated MCF10A cells were also analyzed by pathway analysis using the network building tool MetaCore (Thomson Reuters, Philadelphia, PA, USA) consisting of millions of relationships between proteins derived from publications about proteins and small molecules (including direct protein interaction, transcriptional regulation, binding, enzyme-substrate, and other structural or functional relationships). Results, i.e. maps of protein lists from the uploaded dataset, were then compared with all the possible pathway maps for all the proteins in the database, and the p-value was calculated based on the hypergeometric distribution probability test. The most representative networks that were significantly changed were selected and analyzed.
Real-Time Quantitative Reverse Transcription PCR. Candidate genes for qRT-PCR analysis were chosen based on the microarray results. One microgram of total RNA was reverse-transcribed into cDNA and analyzed by real-time PCR as previously described (22). The oligonucleotide primers were selected with Primer3 software (25-26) and tested for their human specificity using the NCBI database. Primer sequences (forward and reverse) used are listed in Table I. Quantitative data, normalized versus 18S rRNA gene, were analyzed by the average of triplicate cycle thresholds (Ct) according to the 2−ΔΔct method using SDS software (Applied Biosystems, Carlsbad, CA). The data shown were generated from three independent experiments and the values are expressed relative to mRNA levels in the untreated MCF10A cells used as control sample as the mean±SD.
Western blot analysis. Whole-cell lysates from 4 to 6×106 treated and untreated cells were collected as recently described (22). Western blots experiments were performed as previously reported (22). The following primary antibodies were used: β-actin (Sigma, St. Louis, MO, USA); poly (ADP-ribose) polymerase 1 (PARP), Polo-like kinase 1 (PLK1), phospho-epidermal growth factor receptor (Cell Signaling Technologies, Danvers, MA, USA); and pro-caspase-8, EGFR, p53, c-MYC, (Santa Cruz, Biotechnology Inc, Heidelberg, Germany).
Results
Clonogenic assay and morphological analysis. In order to assess MCF10A cell viability in terms of reproductive capacity after IOERT, clonogenic survival assays were performed. Twenty-four hours post-irradiation with 9 and 23 Gy doses, cells were seeded appropriately, maintained under normal culture conditions and analyzed at two to three weeks later for the formation of colonies. The results showed that 23 Gy exposure inhibited the growth and proliferation of MCF10A cells, as colony-forming ability was markedly impaired by irradiation and no colonies were observed. On the contrary, following the 9-Gy boost treatment, an SF of 9.6% was found, indicating the selection of a surviving radioresistant cell fraction with reproductive capacity (Figure 1A and 1B).

Micrographs (×10) of MCF10A cells in culture after 1, 2, and 3 weeks post-Intraoperative electron radiation therapy (IOERT) for morphological evaluation and clonogenic survival assay: A: Untreated cells; B: MCF10A cells treated with 9 Gy and 23 Gy; C: radioresistant surviving fraction of MCF10A cells treated with 9 Gy.
In order to evaluate the effects of IOERT on cell morphology, throughout the course of the clonogenic assays, cells were observed under phase-contrast microscopy and random fields for each treatment were photographed. After irradiation with 9 Gy and 23 Gy doses, the response of MCF10A cells in terms of morphology was similar. Cell damage at both the membranous and cytoplasmic levels was observed, starting 72 h post-treatment and increasing within one week. The total detachment of MCF10A cells treated with 23 Gy from the culture substrate occurred progressively from two to three weeks. In the case of MCF10A cells treated with 9 Gy, a radioresistant growing cell fraction, which we maintained in culture up to three weeks post-treatment, was also observed (Figure 1C). Moreover, unlike IOERT-treated MCF7 cells (22), MCF10A cells did not exhibit morphology of a radiation-induced senescent phenotype. Indeed, biochemical tests for senescence-associated β-galactosidase activity did not reveal any cellular senescence activation in response to IOERT (data not shown).
γ-H2AX immunofluorescence analysis. Following exposure to IR, histone H2AX is immediately phosphorylated at serine 139 (γ-H2AX) with consequent foci formation as a sensitive early cell response to the presence of DNA double-strand breaks. To evaluate the time course of the appearance of γ-H2AX foci in MCF10A cells upon IOERT, we performed direct immunofluorescence analyses after 0.5, 1, 3, 6 and 24 h of exposure to 9 and 23 Gy IR doses. γ-H2AX foci formation occurred within 30 min of irradiation at 9 and 23 Gy doses. In particular, in MCF10A cells treated 9 Gy, the number of foci gradually decreased at 6 and 24 h post-treatment, whereas in MCF10A cells treated with 23 Gy, it remained high with respect to untreated cells at 24 h after irradiation (Figures 2 and 3). These results suggest that foci formation in MCF10A cells was rapid, with a dose-dependent increase following exposure to IOERT.

Micrographs (×20) of gamma-H2AX immunofluorescence (γH2AX) analysis in MCF10A cells after 30 minutes, and 1, 3, 6, and 24 h exposure to 9 and 23 Gy doses.
Overview of cDNA microarray gene expression. In this study, a Two-Color Microarray-Based Gene Expression Analysis (Agilent Technologies) was conducted on 9 Gy- and 23 Gy-treated MCF10A cells using untreated MCF10A cells as a reference sample. Comparative differential gene-expression analysis revealed that in MCF10A cells treated with 9 Gy, expression levels of 72 genes were significantly altered, by 1.5-fold or greater, compared to the untreated MCF10A cell reference group: 18 genes were down-regulated and 54 were up-regulated. Moreover, comparative differential gene-expression analysis revealed that 451 genes in MCF10A cells treated with 23 Gy had significantly altered expression levels compared to the untreated MCF10A reference group: 226 genes were down-regulated and 225 were up-regulated (GSE65954). Up- and down-regulated transcripts were selected and grouped according to their involvement in specific biological pathways using integrated pathway enrichment analysis with GeneGo MetaCore. Data sets were loaded into Metacore software and the top enriched canonical metabolic pathways were analyzed. The result of this mapping revealed involvement of a set of factors controlling specific networks such as regulation of cellular process, inflammation, tissue degradation, cell-cycle modulation, and chromatin modification in comparison to the reference sample. Candidate genes were selected, validated and analyzed using the PubMatrix tool, as previously described (22, 27).
Up- and down-regulated genes in MCF10A cells treated with 9 Gy.
Up- and down-regulated genes in MCF10A cells treated with 23 Gy.
Microarray validation experiments. Genes for microarray validation experiments were chosen based on two considerations: i) factors known to be modulated by IR; and ii) less-known genes involved in cell response to high radiation doses for proposal as new molecular markers. In order to identify possible documented relationships between microarray gene-expression lists and processes known to be involved in cell response to IR treatment, we used the PubMatrix V2.1 tool. In this way, bibliographic relationships between differentially expressed genes and some selected queries such as IR, radiation, cancer, BC, apoptosis, inflammation, DNA damage and DNA repair were analyzed. Moreover, based on the microarray data set, the PubMatrix results and MetaCore analyses, we chose 27 candidate genes and performed qRT-PCR validation experiments (Tables II and III). In MCF10A cells treated with 9 Gy, 13 selected genes were validated: 10 genes were up-regulated and among these, the following seven genes were described as being involved in the positive regulation of the cell cycle and nuclear division: G2 and S-phase expressed 1 (GTSE1), Aurora kinase A, transcript variant 1 (AURKA), Cell division cycle 20 (CDC20), CDC25C, PLK1, Centromere protein F, 350/400kDa (mitosin) (CENPF) and Kinesin family member 2C (KIF2C) (Table II). As also shown in Table II, the following three genes were down-regulated: Lysine (K)-specific methyltransferase 2A (MLL), Chemokine (C-X-C motif) receptor 3 (CXCR3) and Zinc finger, -type containing 15 (ZDHHC15). On the other hand, in MCF10A cells treated with 23 Gy, 14 selected genes were validated. As shown in Table III, five genes were up-regulated: Caspase 8, apoptosis-related cysteine peptidase (CASP8), Cyclin-dependent kinase inhibitor 1A (p21, CIP1) (CDKN1A/p21), Growth arrest and DNA-damage-inducible, beta (GADD45B), Notch 1 (NOTCH1) and tumor protein p53 inducible nuclear protein 1 (TP53INP); while nine genes of the histone cluster and involved in cell-cycle modulation, such as CCNB1, CDKN3, Histone cluster 1, H4b (HIST1H4B), HIST1H4C, HIST1H4D, Lamin B1 (LMNB1), NDC80 kinetochore complex component (NDC80), NIMA-related kinase 2 (NEK2) and PLK1, were down-regulated.
Protein expression in response to IOERT treatment. Based on microarray data and following our assumptions regarding possible IOERT-activated cell processes, candidate proteins were selected and assayed by western blot analysis in time-course experiments after 24, 48, 72 h of exposure to 9 and 23 Gy IR doses. A peak of PLK1 expression at 24 h post-IOERT treatment and a progressive decrease at 48 and 72 h were observed in both 9 Gy- and 23 Gy-treated MCF10A cells (Figure 4). The prolonged PLK1 depletion could activate the DNA-damage checkpoint with cell-cycle arrest, as reported by Lei and Erickson (28). These observations are also in line with microarray data analysis. Moreover, at the molecular level, we did not observe an induction of intrinsic apoptotic response, as evidenced by the absence of PARP fragmentation in time-course experiments. PARP levels remained almost unchanged following both treatments (Figure 4). The extrinsic apoptotic pathway, however, seemed to be activated, as a time-dependent increase in the expression of Fas cell surface death receptor (FAS) and a simultaneous decrease in the expression of pro-caspase-8 were observed after both treatments. Finally, a compensatory activation of cellular defense mechanisms, suggested by the increased expression of p53, c-MYC, EGFR and by the mild activation of EGFR-mediated signaling pathway, as also suggested by increased phosphorylation levels of EGFR following treatments, were observed.
Discussion
Medical applications of high doses of charged particles, such as those used during IOERT, involve the exposure of normal cells, tissues and organs proximal to the tumor. The radiobiology of healthy cells and tissue response to IR is a topic of interest which may contribute to avoiding the impairment of normal tissue and organ function and to reducing risks of secondary cancer. To date, few articles have described the biological and molecular basis of IOERT effects (22; 29-30). In particular, gene-expression profiles of breast normal cells induced by high IR doses need to be properly addressed. The purpose of the study was to analyze cell and gene expression response following IOERT treatment with 9 and 23 Gy doses (IOERT boost and exclusive, respectively) in human non-tumorigenic MCF10A mammary cell line as a model of normal breast epithelial cells. To the best of our knowledge, no studies have examined cell and gene expression changes after high-dose electron irradiation in MCF10A cells.

Quantitative presentation of the mean gamma-H2AX fluorescence intensity for MCF10A cells 30 min, and 1, 3, 6, and 24 h after exposure to 9 and 23 Gy.
Firstly, we assessed cell viability in terms of reproductive capacity performing a clonogenic survival assay and observed that the 23 Gy exposure inhibited the growth and proliferation of MCF10A cells. On the contrary, following the 9-Gy boost treatment, a surviving radioresistant cell fraction with reproductive capacity was found. Immunofluorescence analyses showed that γ-H2AX foci formation rapidly increased in a dose-dependent manner following both IOERT modalities, in MCF10A cells treated with 9 Gy, the number of foci gradually decreased after-irradiation, whereas in cells treated with 23 Gy, it remained high at 24 h post-treatment. Foci formation at sites of double-strand breaks reveals the induction of DNA-repair mechanisms, however, if such damage is not adequately repaired it can lead to cell clonogenicity loss via the generation of lethal chromosomal aberrations, apoptotic cell death or cellular senescence (9, 12). Unlike the senescent features that we described in MCF7 cells following 9 and 23 Gy exposures (22), MCF10A cells did not show a radiation-induced senescent phenotype, as displayed by morphological traits and by a lack of β-galactosidase activity (data not shown).

Western blotting analysis for protein response factors.
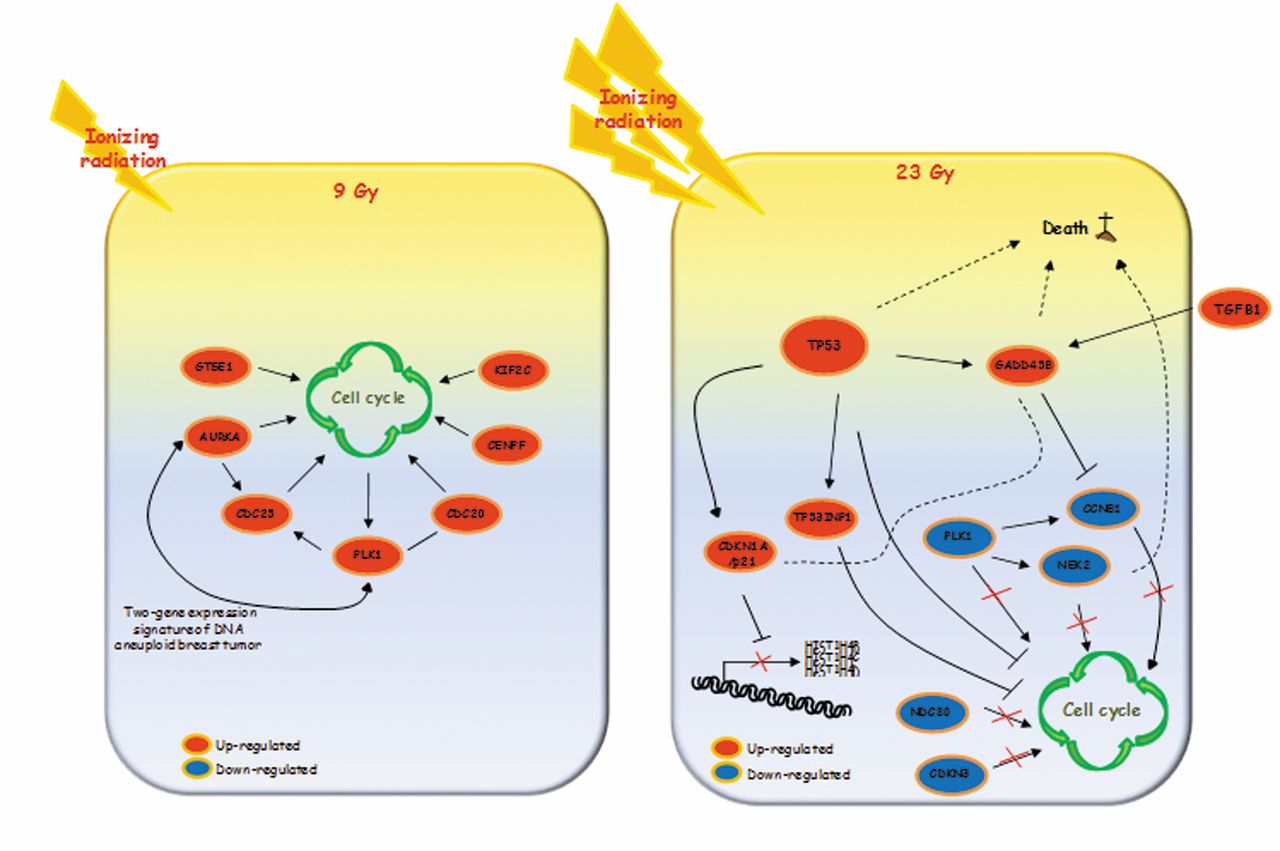
In addition, 24 h after treatments the cell networks involved in IOERT response appeared to be dose-dependent. More precisely, our results revealed that the magnitude of transcriptional variation, defined as the number of differentially expressed genes, could drive two different cell fate decisions in a dose-dependent manner. In order to highlight genes and networks activated after IOERT, we used selected validated genes to design two descriptive models for each dose delivered (Figure 5; models A and B). As reported in model A of Figure 5, the gene-expression profile of MCF10A cells treated with 9 Gy showed involvement of positive cell-cycle modulators. IR is known to activate both pro- and anti-proliferative signal pathways, producing an imbalance in the cell survival vs. death decision (10, 13). More precisely, MCF10A cells treated with 9 Gy activate genes involved in cell-cycle regulation, as suggested by the up-regulation of GSTE1, PLK1, AURKA, CDC25C, CDC20, CENPF and KIF2C genes. Overall, most of these genes code proteins involved in spindle formation, centrosome maturation, mitotic processes, and chromosome instability.
GTSE1 protein, localized on the microtubules, plays a role during the S/G2 phase transition of the cell cycle and is significantly induced by different DNA-damaging agents, including gamma-irradiation and chemotherapeutic drugs (31). Some authors reported a down-regulation of GTSE1 mRNA after stress exposure such as IR, which leads to the activation of apoptotic process (31-32). Recently, GTSE1 protein was also described to play a role in promoting cell survival. In response to DNA damage, after phosphorylation mediated by PLK1, GTSE1 accumulates in the nucleus and binds the tumor-suppressor protein p53; exporting the protein from the nucleus to the cytoplasm, it represses p53 ability to induce apoptosis (33-36). In turn, our results show that PLK1 gene was overexpressed in MCF10A cells treated with 9 Gy, this may contribute to development of the surviving radioresistant cell fraction. PLK1 represents a well-established factor that plays an important role in cell-cycle regulation, acting in centrosome maturation, spindle formation, mitotic entry and cytokinesis (37-38). On the contrary, PLK1 inhibition induces cell-cycle arrest, with subsequent cell death induction. It is interesting to note that a pre-treatment with PLK1 inhibitors sensitized human medulloblastoma cells to IR (37), thus down-regulation of the PLK1 gene observed in MCF10A cells treated with 23 Gy could promote cell death (39-41). AURKA promotes cell-cycle progression, regulating the transition from the G2 to M phase, the centrosome function and assembly of the mitotic spindle. It has also been shown to modulate the activity of tumor suppressors such as p53 and is responsible for BReast CAncer 1, early onset (BRCA1) protein phosphorylation after DNA damage (42-45). In our model, AURKA might promote cell survival following IOERT boost treatment. CDC20 and CDC25C were also up-regulated in MCF10A cells treated with 9 Gy. CDC20 appears to act as a regulatory protein, interacting with several other factors, and is required for two distinct microtubule-dependent processes: nuclear movement prior to anaphase, and chromosome separation. Moreover, CDC25C functions are to drive cell-cycle transition by de-phosphorylating and activating CDKs and to trigger entry into mitosis (22). Thus, in line with the above described gene functions, the cell cycle appears to be activated by multiple regulators after IOERT boost. CENPF and KIF2C, both up-regulated after 9-Gy exposure, have a key role during chromosome segregation in mitosis (46-48). More precisely, CENPF is involved in centromere formation and kinetochore organization during mitosis (46-51). In addition, KIF2C plays important roles in chromosome segregation and in the correction of improper kinetochore–microtubule interactions during mitosis and is regulated by aurora kinase-B (52-54).

Two descriptive models of genes and networks activated in MCF10A cells after Intraoperative electron radiation therapy (IOERT) using the two doses of 9 Gy (model A) and 23 Gy (model B).
In summary, according to our hypothesis, in MCF10A cells treated with 9 Gy, the cell cycle appears to be positively modulated at the transcriptional level by several key factors, which, to our knowledge, have never been described as being correlated to IR cell response. Furthermore, we suggest that the selected up-regulated genes, such as those involved in mitotic aberrations, should be further investigated in order to highlight their possible roles in the molecular mechanisms of mammary carcinogenesis after IR exposure (55-58).
Figure 5 also shows the descriptive model B of selected and validated genes proposed for MFC10A cells treated with 23 Gy of IR. Unlike model A, model B suggests inhibition of the cell cycle through down-regulation of its positive regulators, such as PLK1, NDC80, CDKN3, CCNB1 and NEK2 genes. Overall, these genes have been described as regulators of late cell-cycle phases. More precisely, the NDC80 kinetochore complex component organizes and stabilizes the microtubule-kinetochore interactions and is required for proper chromosome segregation. NDC80, together with other genes such as CDKN3, was found to be altered in tumor epithelial cells during malignant transformation and also correlated with poor tumor differentiation and advanced tumor stage (39, 59). CCNB1 and NEK2, both down-regulated in MCF10A cells treated with 23 Gy, have been described as regulatory factors of the cell cycle, supporting our assumption of an inhibition operating at the transcriptional level (60). In particular, it has been shown that NEK2-knockdown induces aneuploidy and cell-cycle arrest, ultimately leading to cell death. For these reasons, this protein was recently considered an attractive and novel therapeutic target for BC treatment (60-62).
As recently reported, IR induces down-regulation of histone mRNA levels in mammalian cells (22, 63). IR-induced inhibition of histone gene transcription depends on p21 protein expression, which was up-regulated in MCF10A cells treated with 23 Gy. It has been reported that exposure to high- and low-linear energy transfer radiation negatively regulates histone gene expression in human lymphoblastoid and colon cancer cell lines (64). In line with these data, the gene-expression profile of MCF10A cells treated with 23 Gy showed a large number of histone genes to be down-regulated. Three of these were validated, confirming their down-regulation after a high dose of IR; to our knowledge, this result is described for the first time in breast cells (22, 63). In addition in our model B, TP53INP1, a p53-inducible gene able to modulate biological activities of p53, was up-regulated (64-69). Thus, we assayed p53 gene status by qRT-PCR and found it to be up-regulated in MCF10A cells treated with 23 Gy (6.1-fold change with respect to untreated MCF10A cells), as also shown in model B (Figure 5B) (64-69). Cell death programming seems also to be positively modulated by the up-regulation of p53, TP53INP1, CASP8 and GADD45B genes (Figure 5B). Moreover, by western blot analysis, we did not observe induction of intrinsic apoptotic response, as evidenced by the absence of PARP fragmentation. PARP protein levels remained unchanged following both treatments. On the contrary, the extrinsic apoptotic pathway seemed to be activated. Indeed, a time-dependent increase in FAS expression and a simultaneous decrease of pro-caspase-8 expression was observed after both treatments in immunoblotting experiments. Moreover, GADD45B is a member of a group of genes whose transcript levels increased following stressful growth-arrest conditions and treatment with DNA-damaging agents (70). Our findings are in line with several studies that report the involvement of the above-mentioned genes in cell-fate decisions and death modulation after IR exposure (65, 71-72).
Our study, for the first time, indicates which genes are down- or up-regulated after IOERT in non-tumorigenic MCF10A cells and the molecular networks able to regulate survival or death cell decision. We highlight the involvement of well- and lesser-known genes related to the IR response, which are able to drive cell fate in opposite ways. The coded proteins might activate a complex network that positively regulates the cell cycle, promoting radioresistance in MCF10A cells treated with 9 Gy leading to a surviving cell fraction, or inhibits cell-cycle progression in MCF10A cells treated with 23 Gy. Considering their important roles in cell response to high radiation doses such as those used during IOERT, we believe that the genes identified could act as prognostic indicators for RT. For these reasons they need to be further studied in order to improve our knowledge over cell radiation effects.
Conclusion
The success of RT mainly depends on the total administered dose. This must be homogeneously delivered to the tumor and must preserve the surrounding healthy tissue. The radiobiology of the healthy tissue response to IR is a topic of interest that needs more investigation.
High-throughput methodologies, such as DNA microarray, allow the analysis of the mRNA expression of thousands of genes simultaneously in order to discover new genes and pathways as targets of response to IOERT. We observed consistent differences in transcription among the two treatments used and the magnitude of transcriptional variation was dose-dependent. We highlighted novel genes able to activate molecular networks contributing to guiding cell-fate decisions, which may provide the opportunity to explore molecular target-directed interventions in the future.
Acknowledgements
This work was supported by FIRB/MERIT project (RBNE089KHH). The authors thank Marylia Di Stefano, Antonina Azzolina and Patrizia Rubino for their excellent technical assistance.
Footnotes
↵* These Authors contributed equally to this study.
This article is freely accessible online.
Conflicts of Interest
The Authors declare that they have no competing interests.
- Received February 25, 2015.
- Revision received March 12, 2015.
- Accepted March 16, 2015.
- Copyright© 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}