
Fucoxanthin Prevents Pancreatic Tumorigenesis in C57BL/6J Mice That Received Allogenic and Orthotopic Transplants of Cancer Cells

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
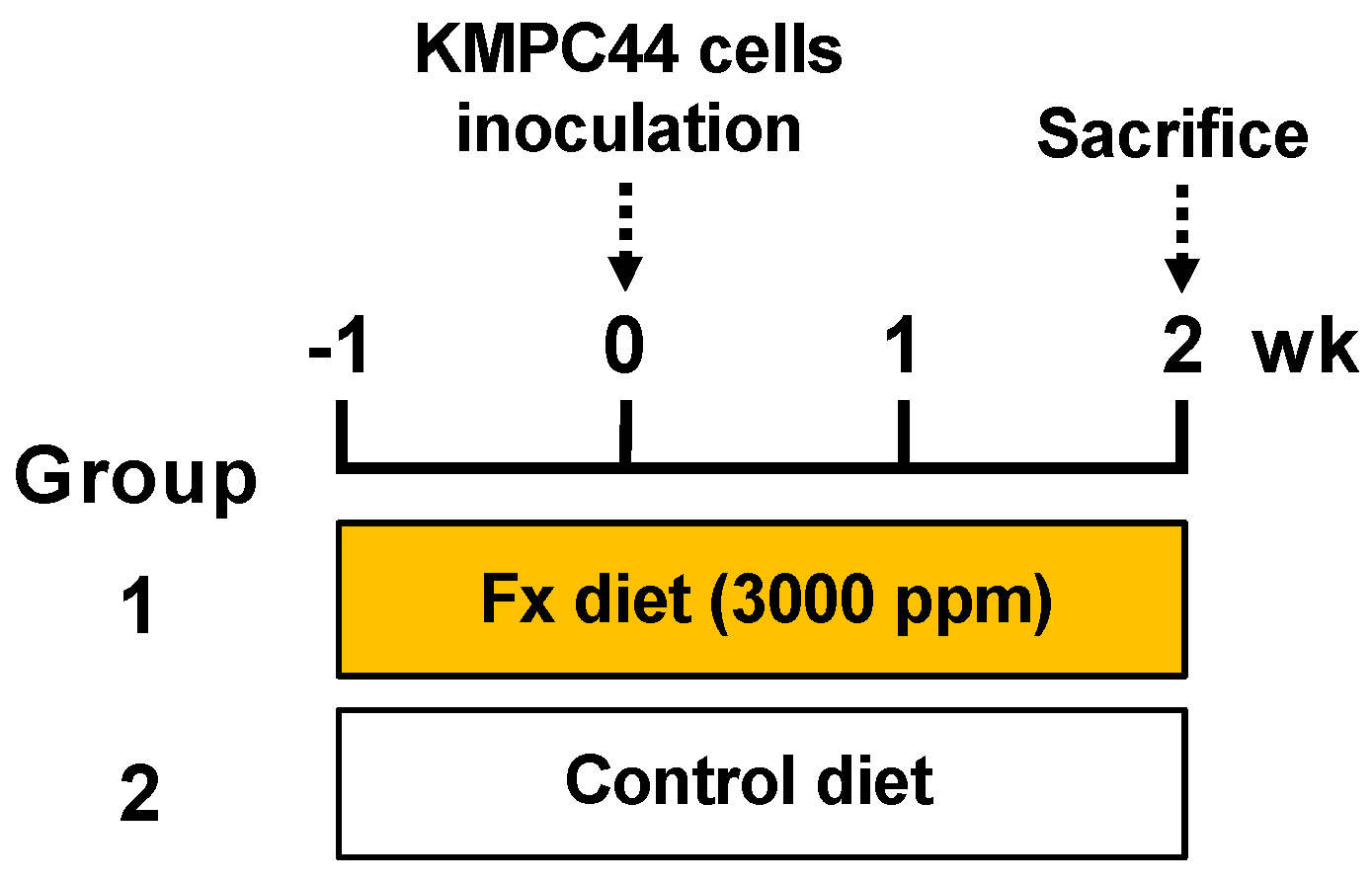
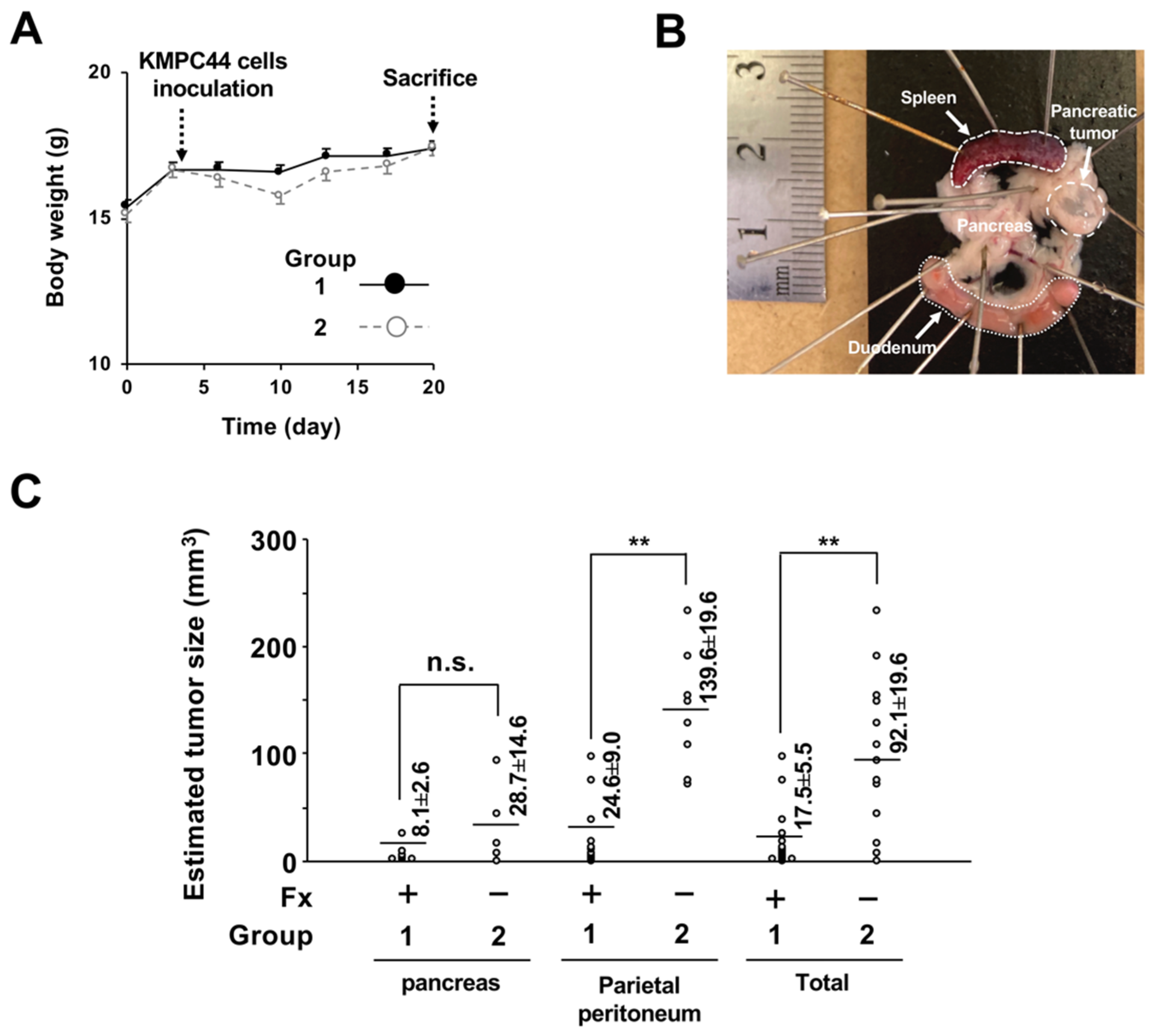
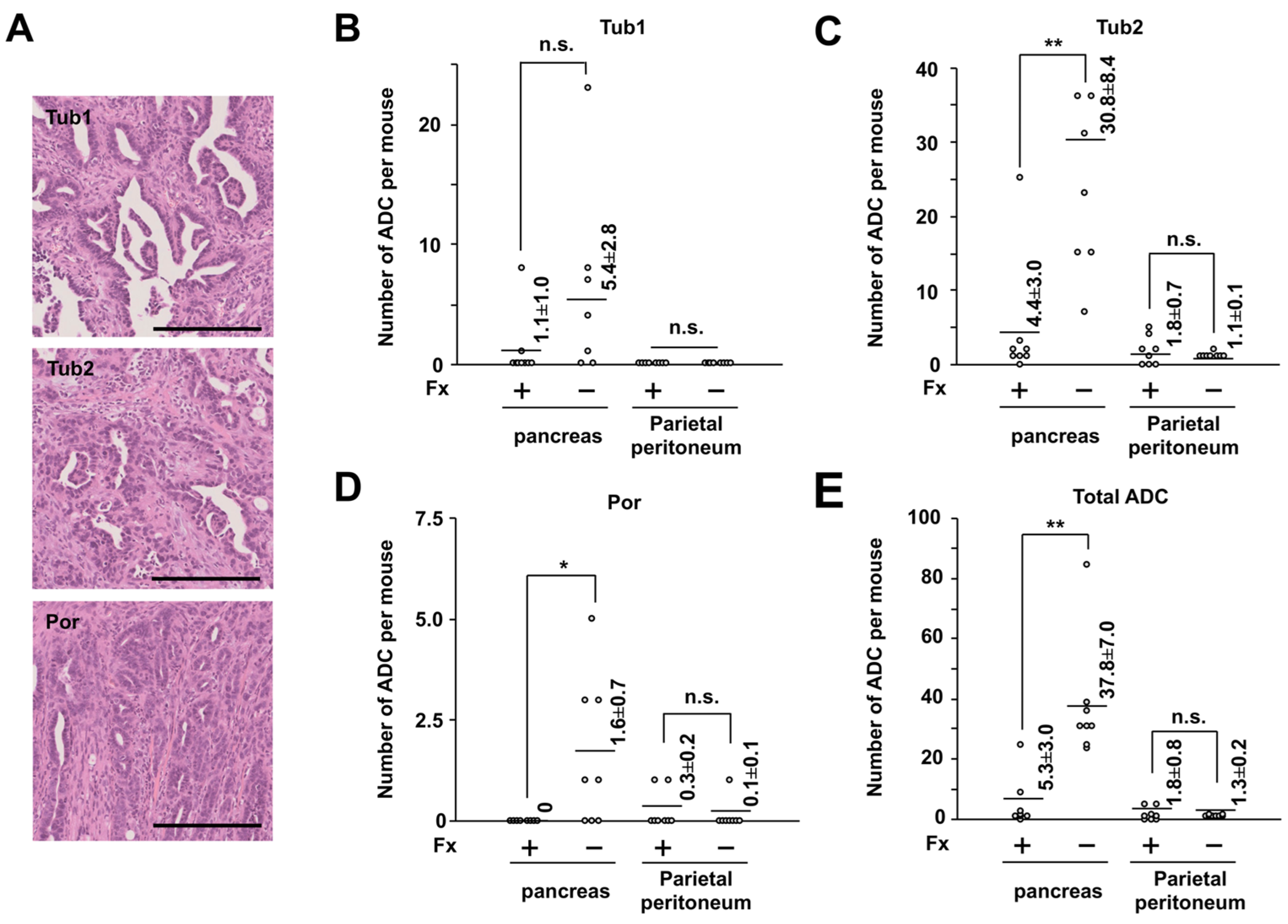
2.1. Cancer Chemopreventive Effect of Fx on a Pancreatic Cancer Model Mouse with Allogenic and Orthotopic Transplantations of KMPC44 Cells
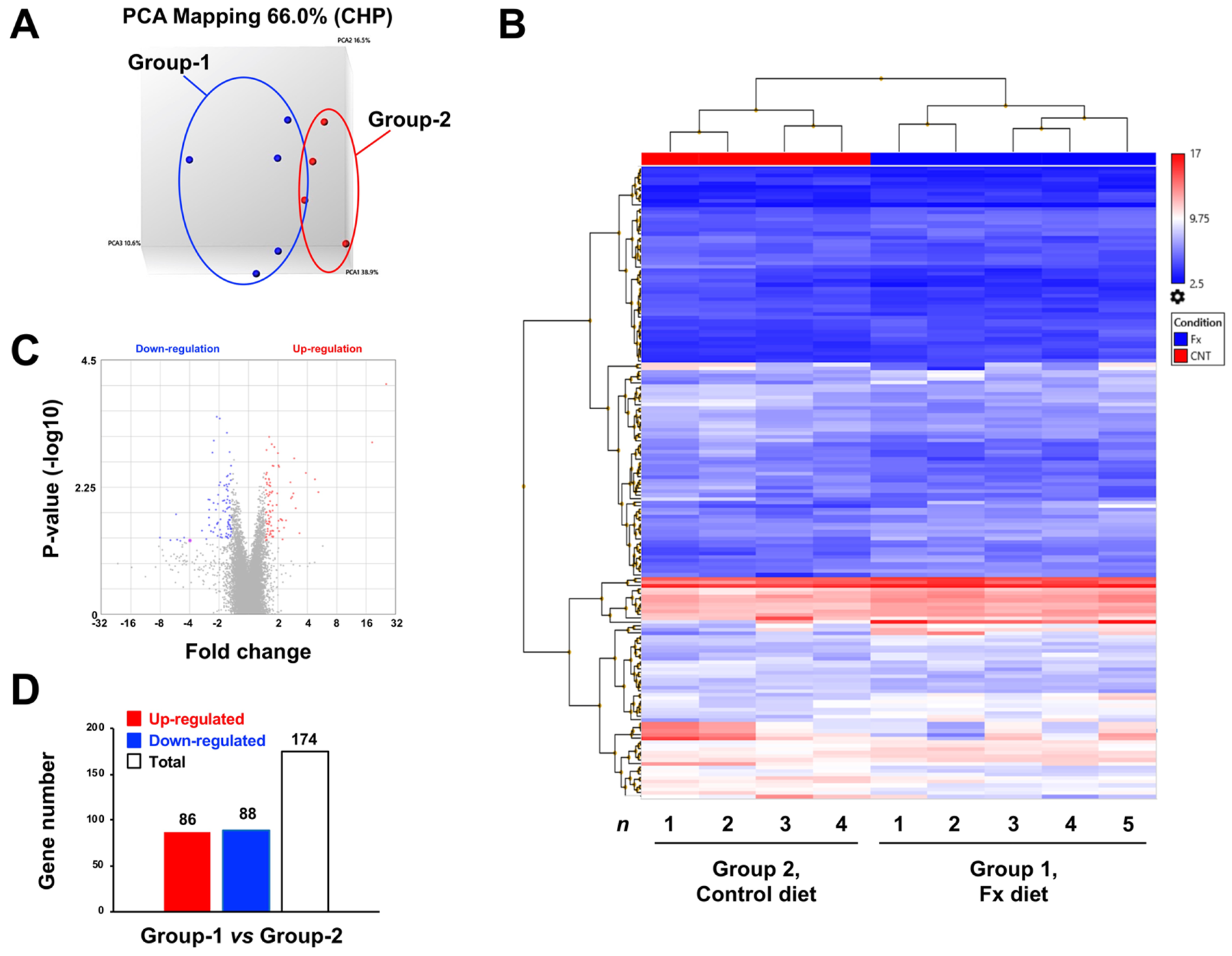
2.2. Altering Effect of Fx on the Transcriptome in Tumor Tissue of a Pancreatic Cancer Model Mouse

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Description | Fold Change 2 | p-Value 3 |
|---|---|---|---|
| Chil4 | Chitinase-like 4 | 25.77 | 0.0001 |
| Chil3 | Chitinase-like 3 | 18.43 | 0.0009 |
| Clca1 | Chloride channel accessory 1 | 5.20 | 0.0069 |
| Ccl24 | Chemokine (C-C motif) ligand 24 | 4.80 | 0.0041 |
| Ccl17 | Chemokine (C-C motif) ligand 17 | 3.87 | 0.0032 |
| Clca3b | Chloride channel accessory 3B | 3.32 | 0.0368 |
| Sprr1a | Small proline-rich protein 1A | 3.03 | 0.0132 |
| S100a8 | S100 calcium binding protein A8 (calgranulin A) | 2.94 | 0.0017 |
| Duox2 | Dual oxidase 2 | 2.88 | 0.0072 |
| Tarm1 | T cell-interacting, activating receptor on myeloid cells 1 | 2.72 | 0.0089 |
| Duoxa2 | Dual oxidase maturation factor 2 | 2.68 | 0.0046 |
| Anxa10 | Annexin A10 | 2.43 | 0.0219 |
| Ppp1r35 | Protein phosphatase 1, regulatory subunit 35 | 2.31 | 0.0293 |
| F7 | Coagulation factor VII | 2.21 | 0.0218 |
| A630095E13Rik | RIKEN cDNA A630095E13 gene | 2.17 | 0.0211 |
| Aldh1a3 | Aldehyde dehydrogenase family 1, subfamily A3 | 2.13 | 0.0205 |
| Creb3l1 | cAMP responsive element binding protein 3-like 1 | 2.06 | 0.0024 |
| Krt19 | Keratin 19 | 2.06 | 0.0337 |
| Ptgs2 | Prostaglandin-endoperoxide synthase 2 | 2.03 | 0.0219 |
| Ccl22 | Chemokine (C-C motif) ligand 22 | 2.02 | 0.0023 |
| Gene Symbol | Description | Fold Change 2 | p-Value 3 |
|---|---|---|---|
| Prss3 | Protease, serine 3 | −8.03 | 0.0439 |
| Klk1b5 | Kallikrein 1-related peptidase b5 | −6.31 | 0.0491 |
| Gc | Group specific component | −5.52 | 0.0169 |
| Klk1b4 | Kallikrein 1-related peptidase b4 | −5.36 | 0.0476 |
| Ccl21a4 | Chemokine (C-C motif) ligand 21A (serine) | −3.98 | 0.0493 |
| Mmrn1 | Multimerin 1 | −2.99 | 0.0329 |
| Dpt | Dermatopontin | −2.71 | 0.0464 |
| Bank1 | B cell scaffold protein with ankyrin repeats 1 | −2.57 | 0.0092 |
| Xlr4b | X-linked lymphocyte-regulated 4B | −2.54 | 0.0193 |
| Gkn3 | Gastrokine 3 | −2.53 | 0.014 |
| Tbx1 | T-box 1 | −2.49 | 0.0094 |
| Cd209f | CD209f antigen | −2.43 | 0.0338 |
| Epha7 | Eph receptor A7 | −2.34 | 0.0014 |
| Slc30a8 | Solute carrier family 30 (zinc transporter), member 8 | −2.29 | 0.0349 |
| Cd19 | CD19 antigen | −2.27 | 0.0107 |
| Egflam | EGF-like, fibronectin type III and laminin G domains | −2.26 | 0.0008 |
| Btla | B and T lymphocyte attenuator | −2.15 | 0.014 |
| 5330417C22Rik | RIKEN cDNA 5330417C22 gene | −2.11 | 0.0003 |
| Kmo | Kynurenine 3-monooxygenase (kynurenine 3-hydroxylase) | −2.09 | 0.025 |
| Klk1b8 | Kallikrein 1-related peptidase b8 | −2.09 | 0.025 |
| Cpn1 | Carboxypeptidase N, polypeptide 1 | −2.09 | 0.0246 |
| Vtn | Vitronectin | −2.07 | 0.0409 |
| Efcab3 | EF-hand calcium binding domain 3 | −2.06 | 0.0237 |
| Klk1b3 | Kallikrein 1-related peptidase b3 | −2.06 | 0.011 |
| Triqk | Triple QxxK/R motif containing | −2.05 | 0.0077 |
| Ntrk2 | Neurotrophic tyrosine kinase, receptor, type 2 | −2.00 | 0.0052 |
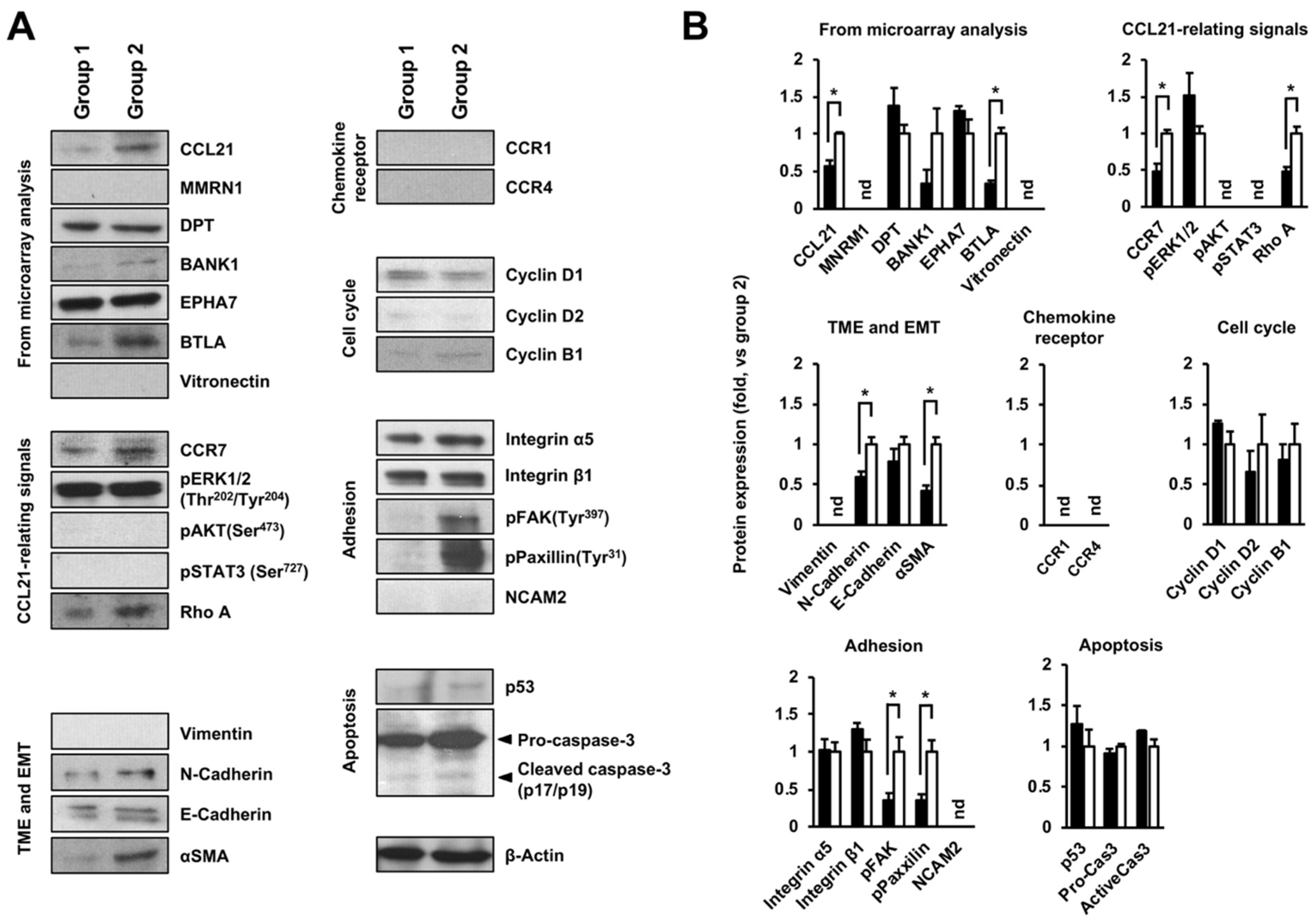
2.3. Altering Effect of Fx on the Protein Expressions in Tumor Tissue of a Pancreatic Cancer Model Mouse
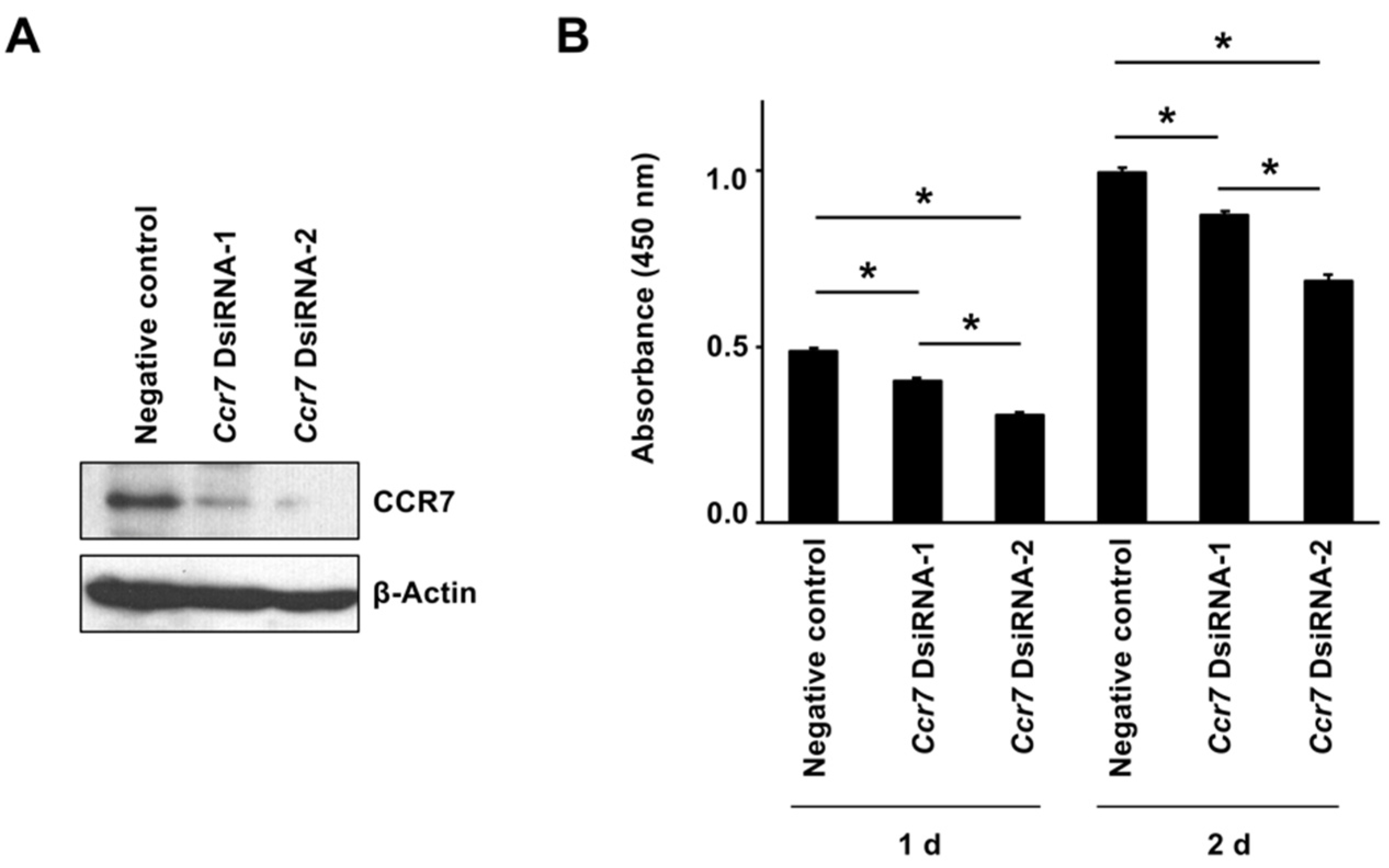
2.4. Altering Effect of Fx on Cell Growth in KMPC44 Cells with Ccr7 Knockdown
3. Discussion
4. Conclusions
5. Materials and Methods
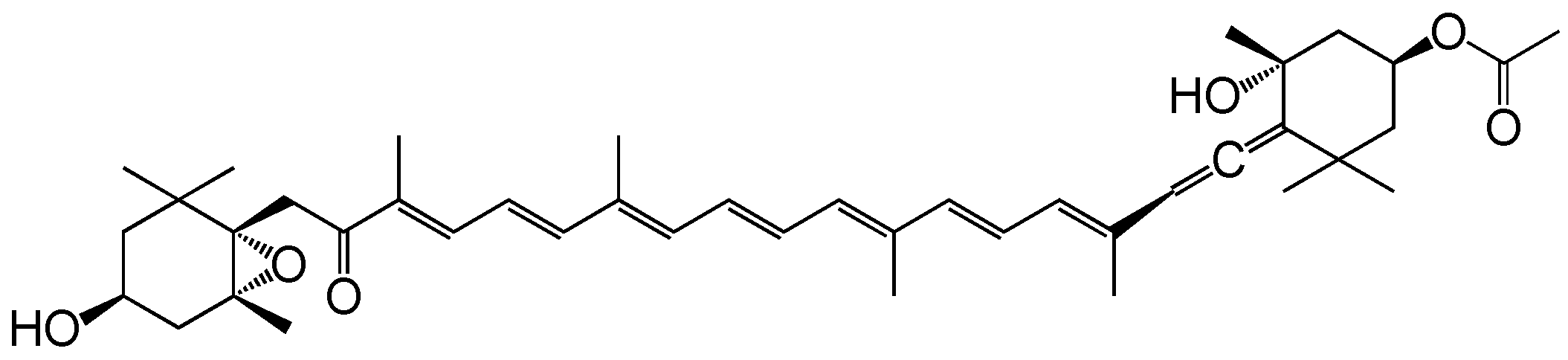
5.1. Chemical
5.2. Cell Culture
5.3. Construction of Pancreatic Cancer Model Mice with Allogenic and Orthotopic Transplantations of KMPC44 Cells, and Animal Experiments
5.4. Microarray Analysis
5.5. Western Blot Analysis
5.6. Fluorescence Immunohistochemistry
5.7. Gene Knockdown
5.8. Cell Viability Assay
5.9. Statistics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Wehner, M.R.; Matrisian, L.M.; Nead, K.T. Estimated Projection of US Cancer Incidence and Death to 2040. JAMA Netw. Open 2021, 4, e214708. [Google Scholar] [CrossRef]
- Pelosi, E.; Castelli, G.; Testa, U. Pancreatic cancer: Molecular Characterization, Clonal Evolution and Cancer Stem Cells. Biomedicines 2017, 5, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Cancer Society, ACS. Cancer Facts & Figures; American Cancer Society: Atlanta, GA, USA, 2021; Available online: cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2021.html (accessed on 20 October 2021).
- The Cancer Genome Atlas Research Network. Integrated Genomic Characterization of Pancreatic Ductal Adenocarcinoma. Cancer Cell 2017, 32, 185–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waddell, N.; Pajic, M.; Patch, A.M.; Chang, D.K.; Kassahn, K.S.; Bailey, P.; Johns, A.L.; Miller, D.; Nones, K.; Quek, K.; et al. Whole Genomes Redefine the Mutational Landscape of Pancreatic Cancer. Nature 2015, 518, 495–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witkiewicz, A.K.; McMillan, E.A.; Balaji, U.; Baek, G.; Lin, W.C.; Mansour, J.; Mollaee, M.; Wagner, K.U.; Koduru, P.; Yopp, A.; et al. Whole-exome sequencing of pancreatic cancer defines genetic diversity and therapeutic targets. Nat. Commun. 2015, 6, 6744. [Google Scholar] [CrossRef]
- Jones, S.; Zhang, X.; Parsons, D.W.; Lin, J.C.H.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Kamiyama, H.; Jimeno, A.; et al. Core Signaling Pathways in Human Pancreatic Cancers Revealed by Global Genomic Analyses. Science 2008, 321, 1801–1806. [Google Scholar] [CrossRef] [Green Version]
- Cicenas, J.; Kvederaviciute, K.; Meskinyte, I.; Meskinyte-Kausiliene, E.; Skeberdyte, A.; Cicenas, J. KRAS, TP53, CDKN2A, SMAD4, BRCA1, and BRCA2 Mutations in Pancreatic Cancer. Cancers 2017, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Puleo, F.; Nicolle, R.; Blum, Y.; Cros, J.; Marisa, L.; Demetter, P.; Quertinmont, E.; Svrcek, M.; Elarouci, N.; Iovanna, J.; et al. Stratification of Pancreatic Ductal Adenocarcinomas Based on Tumor and Microenvironment Features. Gastroenterology 2018, 155, 1999–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, P.; Chang, D.K.; Nones, K.; Johns, A.L.; Patch, A.M.; Gingras, M.C.; Miller, D.K.; Christ, A.N.; Bruxner, T.J.C.; Quinn, M.C.; et al. Genomic analyses identify molecular subtypes of pancreatic cancer. Nature 2016, 531, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Zhu, M.; Kashyap, P.C.; Chia, N.; Tran, N.H.; McWilliams, R.R.; Bekail-Saab, T.S.; Ma, W.W. The role of microbiome in pancreatic cancer. Cancer Metastasis Rev. 2021, 40, 777–789. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Petricoin, E.F.; Maitra, A.; Rajapakse, V.; King, C.; Jacobetz, M.A.; Ross, S.; Conrads, T.P.; Veenstra, T.D.; Hitt, B.A.; et al. Preinvasive and invasive ductal pancreatic cancer and its early detection in the mouse. Cancer Cell 2003, 4, 437–450. [Google Scholar] [CrossRef] [Green Version]
- Ijichi, H.; Chytil, A.; Gorska, E.; Aakre, M.E.; Fujitani, Y.; Fujitani, S.; Wright, C.V.E.; Moses, H.L. Aggressive pancreatic ductal adenocarcinoma in mice caused by pancreas-specific blockade of transforming growth factor-beta signaling in cooperation with active Kras expression. Genes Dev. 2006, 20, 3147–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hingorani, S.R.; Wang, L.; Multani, A.S.; Combs, C.; Deramaudt, T.B.; Hruban, R.H.; Rustgi, A.K.; Chang, S.; Tuveson, D.A. Trp53R172H and KrasG12D cooperate to promote chromosomal instability and widely metastatic pancreatic ductal adenocarcinoma in mice. Cancer Cell 2005, 7, 469–483. [Google Scholar] [CrossRef] [Green Version]
- Terasaki, M.; Kubota, A.; Kojima, H.; Maeda, H.; Miyashita, K.; Kawagoe, C.; Mutoh, M.; Tanaka, T. Fucoxanthin and Colorectal Cancer Prevention. Cancers 2021, 13, 2379. [Google Scholar] [CrossRef]
- Tavares, R.S.N.; Stuchi Maria-Engler, S.; Colepicolo, P.; Debonsi, H.M.; Schäfer-Korting, M.; Marx, U.; Rigo Gaspar, L.; Zoschke, C. Skin Irritation Testing Beyond Tissue Viability: Fucoxanthin Effects on Inflammation, Homeostasis, and Metabolism. Pharmaceutics 2020, 12, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iio, K.; Okada, Y.; Ishikura, M. Single and 13-week oral toxicity study of fucoxanthin oil from microalgae in rats. Shokuhin Eiseigaku Zasshi 2011, 52, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Beppu, F.; Niwano, Y.; Tsukui, T.; Hosokawa, M.; Miyashita, K. Single and repeated oral dose toxicity study of fucoxanthin (FX), a marine carotenoid, in mice. J. Toxicol. Sci. 2009, 34, 501–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Ozaki, Y.; Mizuno, M.; Yoshida, M.; Nishitani, Y.; Azuma, T.; Komoto, A.; Maoka, T.; Tanino, Y.; Kanazawa, K. Pharmacokinetics of fucoxanthinol in human plasma after the oral administration of kombu extract. Br. J. Nutr. 2012, 107, 1566–1569. [Google Scholar] [CrossRef]
- Yonekura, L.; Kobayashi, M.; Terasaki, M.; Nagao, A. Keto-Carotenoids Are the Major Metabolites of Dietary Lutein and Fucoxanthin in Mouse Tissues. J. Nutr. 2010, 140, 1824–1831. [Google Scholar] [CrossRef]
- Shiratori, K.; Ohgami, K.; Ilieva, I.; Jin, X.H.; Koyama, Y.; Miyashita, K.; Yoshida, K.; Kase, S.; Ohno, S. Effects of fucoxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Exp. Eye Res. 2005, 81, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Nishino, H.; Murakoshi, M.; Tokuda, H.; Satomi, Y. Cancer prevention by carotenoids. Arch. Biochem. Biophys. 2009, 483, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Ono, S.; Hashimoto, S.; Kubota, A.; Kojima, H.; Ohta, T.; Tanaka, T.; Maeda, H.; Miyashita, K.; Mutoh, M. Suppression of C-C chemokine receptor 1 is a key regulation for colon cancer chemoprevention in AOM/DSS mice by fucoxanthin. J. Nutr. Biochem. 2022, 99, 108871. [Google Scholar] [CrossRef]
- Rwigemera, A.; Mamelona, J.; Martin, L.J. Inhibitory effects of fucoxanthinol on the viability of human breast cancer cell lines MCF-7 and MDA-MB-231 are correlated with modulation of the NF-kappaB pathway. Cell Biol. Toxicol. 2014, 30, 157–167. [Google Scholar] [CrossRef]
- Terasaki, M.; Nishizaka, Y.; Murase, W.; Kubota, A.; Kojima, H.; Kojoma, M.; Tanaka, T.; Maeda, H.; Miyashita, K.; Mutoh, M.; et al. Effect of Fucoxanthinol on Pancreatic Ductal Adenocarcinoma Cells from an N-Nitrosobis(2-Oxopropyl)Amine-Initiated Syrian Golden Hamster Pancreatic Carcinogenesis Model. Cancer Genom. Proteom. 2021, 18, 407–423. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Inoue, T.; Murase, W.; Kubota, A.; Kojima, H.; Kojoma, M.; Ohta, T.; Maeda, H.; Miyashita, K.; Mutoh, M.; et al. A Fucoxanthinol Induces Apoptosis in a Pancreatic Intraepithelial Neoplasia Cell. Cancer Genom. Proteom. 2021, 18, 133–146. [Google Scholar] [CrossRef]
- Hitoe, S.; Shimoda, H. Seaweed Fucoxanthin Supplementation Improves Obesity Parameters in Mild Obese Japanese Subjects. Funct. Foods Health Dis. 2017, 7, 246–262. [Google Scholar] [CrossRef]
- Mikami, N.; Hosokawa, M.; Miyashita, K.; Sohma, H.; Ito, Y.M.; Kokai, Y. Reduction of HbA1c levels by fucoxanthin-enriched akamoku oil possibly involves the thrifty allele of uncoupling protein 1 (UCP1): A randomised controlled trial in normal-weight and obese Japanese adults. J. Nutr. Sci. 2017, 6, e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasaki, M.; Uehara, O.; Ogasa, S.; Sano, T.; Kubota, A.; Kojima, H.; Tanaka, T.; Maeda, H.; Miyashita, K.; Mutoh, M. Alteration of fecal microbiota by fucoxanthin results in prevention of colorectal cancer in AOM/DSS-treated mice. Carcinogenesis 2021, 42, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Ikuta, M.; Kojima, H.; Tanaka, T.; Maeda, H.; Miyashita, K.; Mutoh, M. Dietary Fucoxanthin Induces Anoikis in Colorectal Adenocarcinoma by Suppressing Integrin Signaling in a Murine Colorectal Cancer Model. J. Clin. Med. 2020, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Gunn, M.D.; Tangemann, K.; Tam, C.; Cyster, J.G.; Rosen, S.D.; Williams, L.T. A chemokine expressed in lymphoid high endothelial venules promotes the adhesion and chemotaxis of naive T lymphocytes. Proc. Natl. Acad. Sci. USA 1998, 95, 258–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunn, M.D.; Tam, S.K.C.; Kakiuchi, T.; Matsuzawa, A.; Williams, L.T.; Nakano, H. Mice Lacking Expression of Secondary Lymphoid Organ Chemokine Have Defects in Lymphocyte Homing and Dendritic Cell Localization. J. Exp. Med. 1999, 189, 451–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakata, B.; Fukunaga, S.; Noda, E.; Amano, R.; Yamada, N.; Hirakawa, K. Chemokine Receptor CCR7 Expression Correlates with Lymph Node Metastasis in Pancreatic Cancer. Oncology 2008, 74, 69–75. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, L.; Qiu, X.; Jiang, L.; Huang, B.; Li, H.; Li, Z.; Luo, W.; Wang, E. CCL21/CCR7 Promotes G2/M Phase Progression via the ERK Pathway in Human Non-Small Lung Cancer Cells. PLoS ONE 2011, 6, e21119. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.Y.; Safdar, J.; Li, Z.N.; Fang, Q.G.; Zhang, X.; Xu, Z.F.; Sun, C.F. CCR7 Regulates Cell Migration and Invasion through JAK2/STAT3 in Metastatic Squamous Cell Carcinoma of the Head and Neck. BioMed Res. Int. 2014, 2014, 415375. [Google Scholar] [CrossRef]
- Cai, Q.Y.; Liang, G.Y.; Zheng, Y.F.; Tan, Q.Y.; Wang, R.W.; Li, K. CCR7 enhances the angiogenic capacity of esophageal squamous carcinoma cells in vitro via activation of the NF-κB/VEGF signaling pathway. Am. J. Transl. Res. 2017, 9, 3282–3292. [Google Scholar] [PubMed]
- Li, K.; Xu, B.; Xu, G.; Liu, R. CCR7 regulates Twist to induce the epithelial-mesenchymal transition in pancreatic ductal adenocarcinoma. Tumour Biol. 2016, 37, 419–424. [Google Scholar] [CrossRef]
- Xu, Z.; Zheng, X.; Yang, L.; Liu, F.; Zhang, E.; Duan, W.; Bai, S.; Safdar, J.; Li, Z.; Sun, C. Chemokine receptor 7 promotes tumor migration and invasiveness via the RhoA/ROCK pathway in metastatic squamous cell carcinoma of the head and neck. Oncol. Rep. 2015, 33, 849–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirth, M.; Gandla, J.; Höper, C.; Gaida, M.M.; Agarwal, N.; Simonetti, M.; Demir, A.; Xie, Y.; Weiss, C.; Michalski, C.W.; et al. CXCL10 and CCL21 Promote Migration of Pancreatic Cancer Cells Toward Sensory Neurons and Neural Remodeling in Tumors in Mice, Associated with Pain in Patients. Gastroenterology 2020, 159, 665–681. [Google Scholar] [CrossRef]
- Sperveslage, J.; Frank, S.; Heneweer, C.; Egberts, J.; Schniewind, B.; Buchholz, M.; Bergmann, F.; Giese, N.; Munding, J.; Hahn, S.A. Lack of CCR7 expression is rate limiting for lymphatic spread of pancreatic ductal adenocarcinoma. Int. J. Cancer 2012, 131, E371–E381. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, D.; Li, Y.; Liu, Y.; Xie, X.; Wu, Y.; Zhou, Y.; Ren, J.; Zhang, J.; Zhu, H.; et al. CCL21/CCR7 Axis Contributed to CD133+ Pancreatic Cancer Stem-Like Cell Metastasis via EMT and Erk/NF-Κb Pathway. PLoS ONE 2016, 11, e0158529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Lou, W.; Ji, Y.; Zhang, S. Effect of CCR7, CXCR4 and VEGF-C on the lymph node metastasis of human pancreatic ductal adenocarcinoma. Oncol. Lett. 2013, 5, 1572–1578. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Cui, K.; Wang, C.L.; Wang, A.L.; Zhang, B.; Zhou, W.Y.; Zhao, W.H.; Li, S. The chemotactic interaction between CCL21 and its receptor, CCR7, facilitates the progression of pancreatic cancer via induction of angiogenesis and lymphangiogenesis. J. Hepatobiliary Pancreat. Sci. 2011, 18, 821–828. [Google Scholar] [CrossRef]
- Watanabe, N.; Gavrieli, M.; Sedy, J.R.; Yang, J.; Fallarino, F.; Loftin, S.K.; Hurchla, M.A.; Zimmerman, N.; Sim, J.; Zang, X.; et al. BTLA is a lymphocyte inhibitory receptor with similarities to CTLA-4 and PD-1. Nat. Immunol. 2003, 4, 670–679. [Google Scholar] [CrossRef]
- Haymaker, C.L.; Wu, R.C.; Ritthipichai, K.; Bernatchez, C.; Forget, M.A.; Chen, J.Q.; Liu, H.; Wang, E.; Marincola, F.; Hwu, P.; et al. BTLA marks a less-differentiated tumor-infiltrating lymphocyte subset in melanoma with enhanced survival properties. Oncoimmunology 2015, 4, e1014246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, K.M.; Nelson, C.A.; Šedý, J.R. Balancing co-stimulation and inhibition with BTLA and HVEM. Nat. Rev. Immunol. 2006, 6, 671–681. [Google Scholar] [CrossRef]
- Derré, L.; Rivals, J.P.; Jandus, C.; Pastor, S.; Rimoldi, D.; Romero, P.; Michielin, O.; Olive, D.; Speiser, D.E. BTLA mediates inhibition of human tumor-specific CD8+ T cells that can be partially reversed by vaccination. J. Clin. Investig. 2010, 120, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.L.; Lin, H.W.; Chien, C.L.; Lai, Y.L.; Sun, W.Z.; Chen, C.A.; Cheng, W.F. BTLA blockade enhances cancer therapy by inhibiting IL-6/IL-10-induced CD19high B lymphocytes. J. Immunother. Cancer 2019, 7, 313. [Google Scholar] [CrossRef]
- Oguro, S.; Ino, Y.; Shimada, K.; Hatanaka, Y.; Matsuno, Y.; Esaki, M.; Nara, S.; Kishi, Y.; Kosuge, T.; Hiraoka, N. Clinical significance of tumor-infiltrating immune cells focusing on BTLA and Cbl-b in patients with gallbladder cancer. Cancer Sci. 2015, 106, 1750–1760. [Google Scholar] [CrossRef]
- Lan, X.; Li, S.; Gao, H.; Nanding, A.; Quan, L.; Yang, C.; Ding, S.; Xue, Y. Increased BTLA and HVEM in gastric cancer are associated with progression and poor prognosis. OncoTargets Ther. 2017, 10, 919–926. [Google Scholar] [CrossRef] [Green Version]
- Bian, B.; Fanale, D.; Dusetti, N.; Roque, J.; Pastor, S.; Chretien, A.S.; Incorvaia, L.; Russo, A.; Olive, D.; Iovanna, J. Prognostic significance of circulating PD-1, PD-L1, pan-BTN3As, BTN3A1 and BTLA in patients with pancreatic adenocarcinoma. Oncoimmunology 2019, 8, e1561120. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Wu, L. Friend or Foe: Prognostic and Immunotherapy Roles of BTLA in Colorectal Cancer. Front. Mol. Biosci. 2020, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Majima, T.; Tsutsumi, M.; Nishino, H.; Tsunoda, T.; Konishi, Y. Inhibitory effects of beta-carotene, palm carotene, and green tea polyphenols on pancreatic carcinogenesis initiated by N-nitrosobis(2-oxopropyl)amine in Syrian golden hamsters. Pancreas 1998, 16, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Appel, M.J.; Roverts, G.; Woutersen, R.A. Inhibitory effects of micronutrients on pancreatic carcinogenesis in azaserine-treated rats. Carcinogenesis 1991, 12, 2157–2161. [Google Scholar] [CrossRef]
- Kuno, T.; Takhashi, S.; Tomita, H.; Hisamatsu, K.; Hara, A.; Hirata, A.; Kobayashi, H.; Mori, H. Preventive effects of fermented brown rice and rice bran against N-nitrosobis(2-oxopropyl)amine-induced pancreatic tumorigenesis in male hamsters. Oncol. Lett. 2015, 10, 3377–3384. [Google Scholar] [CrossRef] [Green Version]
- Okamura, T.; Umemura, T.; Inoue, T.; Tasaki, M.; Ishii, Y.; Nakamura, Y.; Park, E.Y.; Sato, K.; Matsuo, T.; Okamoto, S.; et al. Chemopreventive effects of 4-methylthio-3-butenyl isothiocyanate (Raphasatin) but not curcumin against pancreatic carcinogenesis in hamsters. J. Agric. Food. Chem. 2013, 61, 2103–2108. [Google Scholar] [CrossRef] [PubMed]
- Kuroiwa, Y.; Nishikawa, A.; Kitamura, Y.; Kanki, K.; Ishii, Y.; Umemura, T.; Hirose, M. Protective effects of benzyl isothiocyanate and sulforaphane but not resveratrol against initiation of pancreatic carcinogenesis in hamsters. Cancer Lett. 2006, 241, 275–280. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murase, W.; Kamakura, Y.; Kawakami, S.; Yasuda, A.; Wagatsuma, M.; Kubota, A.; Kojima, H.; Ohta, T.; Takahashi, M.; Mutoh, M.; et al. Fucoxanthin Prevents Pancreatic Tumorigenesis in C57BL/6J Mice That Received Allogenic and Orthotopic Transplants of Cancer Cells. Int. J. Mol. Sci. 2021, 22, 13620. https://doi.org/10.3390/ijms222413620
Murase W, Kamakura Y, Kawakami S, Yasuda A, Wagatsuma M, Kubota A, Kojima H, Ohta T, Takahashi M, Mutoh M, et al. Fucoxanthin Prevents Pancreatic Tumorigenesis in C57BL/6J Mice That Received Allogenic and Orthotopic Transplants of Cancer Cells. International Journal of Molecular Sciences. 2021; 22(24):13620. https://doi.org/10.3390/ijms222413620
Chicago/Turabian StyleMurase, Wataru, Yukino Kamakura, Serina Kawakami, Ayaka Yasuda, Momoka Wagatsuma, Atsuhito Kubota, Hiroyuki Kojima, Tohru Ohta, Mami Takahashi, Michihiro Mutoh, and et al. 2021. "Fucoxanthin Prevents Pancreatic Tumorigenesis in C57BL/6J Mice That Received Allogenic and Orthotopic Transplants of Cancer Cells" International Journal of Molecular Sciences 22, no. 24: 13620. https://doi.org/10.3390/ijms222413620