

Abstract
Cancer chemotherapy relies heavily on DNA damaging agents such as cisplatin to induce tumour cell death. The response of cells to genotoxic insult, including cell cycle arrest, DNA repair and cell death, is mediated by the DNA damage response (DDR). To address the relationship between the DDR and the outcome of exposure, this study utilised a magnetic-activated cell sorting (MACS®)-based approach to isolate apoptotic and non-apoptotic cells from a DNA polymerase eta-deficient human cell line. The pattern of phosphorylation of the key DNA damage response protein RPA2 on serine 4/8 was altered in apoptotic cells isolated following cisplatin treatment. By combining MACS® with multi-antibody screening for phosphorylated proteins, apoptosis-associated changes were characterized in a number of key signalling pathways. Phosphorylation of Erk1 on Thr202/Tyr204, and Erk2 on Thr185/Tyr187 was increased in apoptotic cells. This approach provides novel insights into the relationship between cisplatin-induced protein phosphorylation and the cellular consequences of exposure to this chemotherapeutic agent.
- Apoptotic cell isolation
- RPA
- Erk1/2
The use of DNA damaging agents such as cisplatin in cancer therapy is hampered by tumour resistance, and by secondary cancer development years after the initial treatment. The cellular DNA damage response (DDR) is a complex protein network that senses DNA damage, and transduces a signal to downstream effectors, which ultimately determine cell fate, including cell cycle arrest, cell survival, or cell death (1). Post-translational modification of proteins, including phosphorylation, plays a critical role in DDR signalling (2). The phosphoinositide 3-kinase-like protein kinases (PIKKs), ataxia telangiectasia mutated (ATM), ATM- and Rad3-related (ATR) and DNA-dependent protein kinase catalytic subunit (DNA-PKcs) are apical kinases in the DDR (1). DDR activation affects a variety of cellular processes, including non-nuclear signalling pathways. Recent screens in human cells treated with ionising- or UV-radiation have identified several hundred proteins phosphorylated at PIKK consensus sites that are involved not only in cell cycle regulation, DNA metabolism and apoptosis, but also in cell structure, protein trafficking and signal transduction (3, 4). Inhibition of p38 mitogen-activated protein kinase (p38 MAPK) and jun N-terminal kinase (Jnk) regulates induction of apoptosis in response to DNA damage in mouse embryo fibroblasts (5). Furthermore, extracellular-signal regulated protein kinase (Erk) signalling is required for IR-induced G2/M arrest in human MCF-7 cells (6).
Individual proteins can play multiple roles in the DDR. For example, human single-stranded DNA binding protein, replication protein A (RPA), composed of three subunits (RPA1, RPA2 and RPA3), functions in DNA replication, repair and recombination (7). The RPA2 subunit is hyperphosphorylated by PIKKs on several N-terminal sites in response to IR (8), UV (9, 10), and cisplatin (11), and following induction of Fas-mediated apoptosis in Jurkat cells (12).
Since different signalling events may contribute to cell death or survival in response to DNA damage, the phosphorylation state of individual proteins is predicted to differ between apoptotic and non-apoptotic cells. To directly address this question, the present study utilised a magnetic bead-based system for the separation of apoptotic and non-apoptotic cells, and performed subsequent analysis of the phosphorylation status of a number of proteins in key cell signalling pathways, using multi-antibody screening. The DNA polη-deficient human cell line XP30RO was used as a model system in which to investigate cisplatin-induced protein phosphorylation and apoptosis, since cells lacking DNA polη are more sensitive than normal cells to platinum-based chemotherapeutic drugs, due to a defect in bypass of platinum-induced DNA lesions (11). In addition, a number of cisplatin-induced DNA damage responses are more strongly activated in polη-deficient cells (11) providing a robust signal for phosphoprotein screening. Using magnetic-activated cell sorting (MACS®; Miltenyi Biotech Ltd, Bergish-Gladbach, Germany) in combination with multiprotein immunoblotting provided novel insights into differential protein phosphorylation in annexin V-positive and annexin V-negative human cells following exposure to cisplatin.
Materials and Methods
Cell culture. SV40-transformed XP30RO cells (GM0317A), which lack functional DNA polη due to a mutation in POLH, were cultured as previously described (13, 14). At 24 h before cisplatin treatment, 1.5×106 cells were seeded in 100 mm culture dishes. Cells were treated with DMSO or with cisplatin (Sigma-Aldrich, MO, USA) (11), as indicated in individual experiments.
Isolation of apoptotic cells. Following cisplatin exposure, cells were separated into apoptotic and non-apoptotic populations using the Annexin V MicroBead system (Miltenyi Biotech Ltd.). Cells were harvested by trypsinisation, resuspended in 1X annexin V-binding buffer, and incubated with Annexin V MicroBeads (15 μl) at 8°C for 15 min. The cells were washed and resuspended in 1X annexin V-binding buffer. The cell suspension was then applied to an activated magnetic separation column, type MS (Miltenyi Biotech Ltd.), in an OctoMacs magnetic separator (Miltenyi Biotech Ltd.). Unbound cells were allowed to flow through the column, which was then washed four times with 500 μl of annexin V-binding buffer. The magnetic field was removed and column-bound cells were eluted in 1 ml of 1X annexin V-binding buffer. Cells were separated over a second MS column. The flow-through and column-bound cells were collected for analysis by Western blotting and flow cytometry.
Flow cytometry. A volume of 10 μl FITC-conjugated annexin V (Miltenyi Biotech Ltd.) was added to the cell suspension at the same time as the cells were incubated with annexin V-MicroBeads for cell separation. All steps using FITC-conjugated annexin V were carried out in the dark. Following cell separation, annexin V-FITC staining was analysed immediately using a Becton Dickinson (NJ, USA) FACSCalibur™ flow cytometer, and CellQuest™ software.
Western blotting. Following cell treatment and separation, whole cell lysates were prepared, and immunoblotting was carried out as described previously (10).
Multi-antibody immunoblotting. XP30RO cells were treated either with DMSO or with 5 μg/ml cisplatin for 18 h, and then separated into annexin V-positive and annexin V-negative cell fractions. Protein lysates were prepared according to the manufacturer' recommended protocol (Kinexus Bioinformatics Corporation, Vancouver, Canada.). Protein lysates were prepared from at least three independent experiments for each condition, and were combined to give a total of 400 μg of protein for each sample. Protein samples, at a concentration of 1 mg/ml in a 3:1(v/v) solution of lysis buffer: 4X Laemmli buffer, were shipped to Kinexus for analysis using the Kinetworks™ Phospho-Site Screen (KPSS12.1) (15). At the Kinexus facility, multi-immunoblotting was carried out using a 20-lane multiblotter apparatus and cocktails of the antibodies in the KPSS12.1 screen. Following protein detection, the intensity of individual bands was quantified and normalised between immunoblots, to obtain an intensity value for each band (`normalised trace quantity'), measured in counts per minute (CPM).
Results
Separation of annexin V-positive and annexin V-negative human cells. To compare DNA damage-induced protein phosphorylation in apoptotic and non-apoptotic cells, XP30RO cells were either untreated or treated with cisplatin. Both adherent and suspension cells were harvested 18 h later. Adherent cells were separated into apoptotic and non-apoptotic cells by MACS®, using the annexin V-MicroBead separation system. Redistribution of phosphatidylserine to the outer surface of the plasma membrane is a characteristic of apoptosis in many cell types (16), and specific binding of annexin V to externalised phosphatidylserine (17) has been exploited to isolate apoptotic cells (18-20). Apoptotic XP30RO cells bound to magnetic annexin V beads and were retained in a column upon application of a magnetic field, while unbound cells flowed through (`flow-through cells') (Figure 1). It was possible to obtain an enriched population of cells with externalised phosphatidylserine by applying the eluted column-bound cells to a second MACS® column (Figure 1A).
Flow-through and column-bound cells could be distinguished on the basis of annexin V-binding, as determined by flow cytometry (Figure 1B). In all cases, the peak of FITC-annexin V fluorescence in column-bound XP30RO cells was between one and two orders of magnitude higher than that of flow-through cells. Thus, column-bound cells obtained following separation over two columns were annexin V-positive, and flow-through cells were annexin V-negative, consistent with the separation being mediated by the binding of annexin V to externalised phosphatidylserine. Column-bound annexin V-positive cells represented 3.3% of the untreated cell population, compared to 9% of cells following treatment with 5 μg/ml cisplatin (Figure 1C), as determined by directly counting the number of cells recovered in each fraction. While annexin V-positive cells represented a small proportion of the cell population, the proportion was increased by three-fold following cisplatin treatment. The annexin V-based separation system generated an enriched population of annexin V-positive cells for further analysis.
Annexin V-positive cells are apoptotic. Caspase activation and degradation of caspase substrates are key markers of apoptosis. To characterise the cells obtained following MACS®, cleavage of caspase-3 and cleavage of the caspase substrate poly(ADP-ribose) polymerase-1 (PARP-1) were analysed by Western blotting in protein extracts from unfractionated, annexin V-positive and annexin V-negative cells, and from cells in suspension. A low level of caspase-3 cleavage was detected in extracts of unfractionated cisplatin-treated XP30RO cells (Figure 1D). Consistent with enrichment for apoptotic cells using MACS®, following treatment with 5 μg/ml cisplatin, cleavage of both caspase-3 and PARP-1 was readily detectable in annexin V-positive XP30RO cells, but not in annexin V-negative cells (Figure 1D). In contrast, suspension cells, which were already detached from the surface at the time of harvest, showed a high level of both caspase-3 and PARP-1 cleavage in comparison to annexin V-positive cells (Figure 1D), indicating that the apoptotic programme is not completed in annexin V-positive cells. These cells represent a good model in which to investigate protein phosphorylation in cells undergoing cisplatin-induced apoptosis.
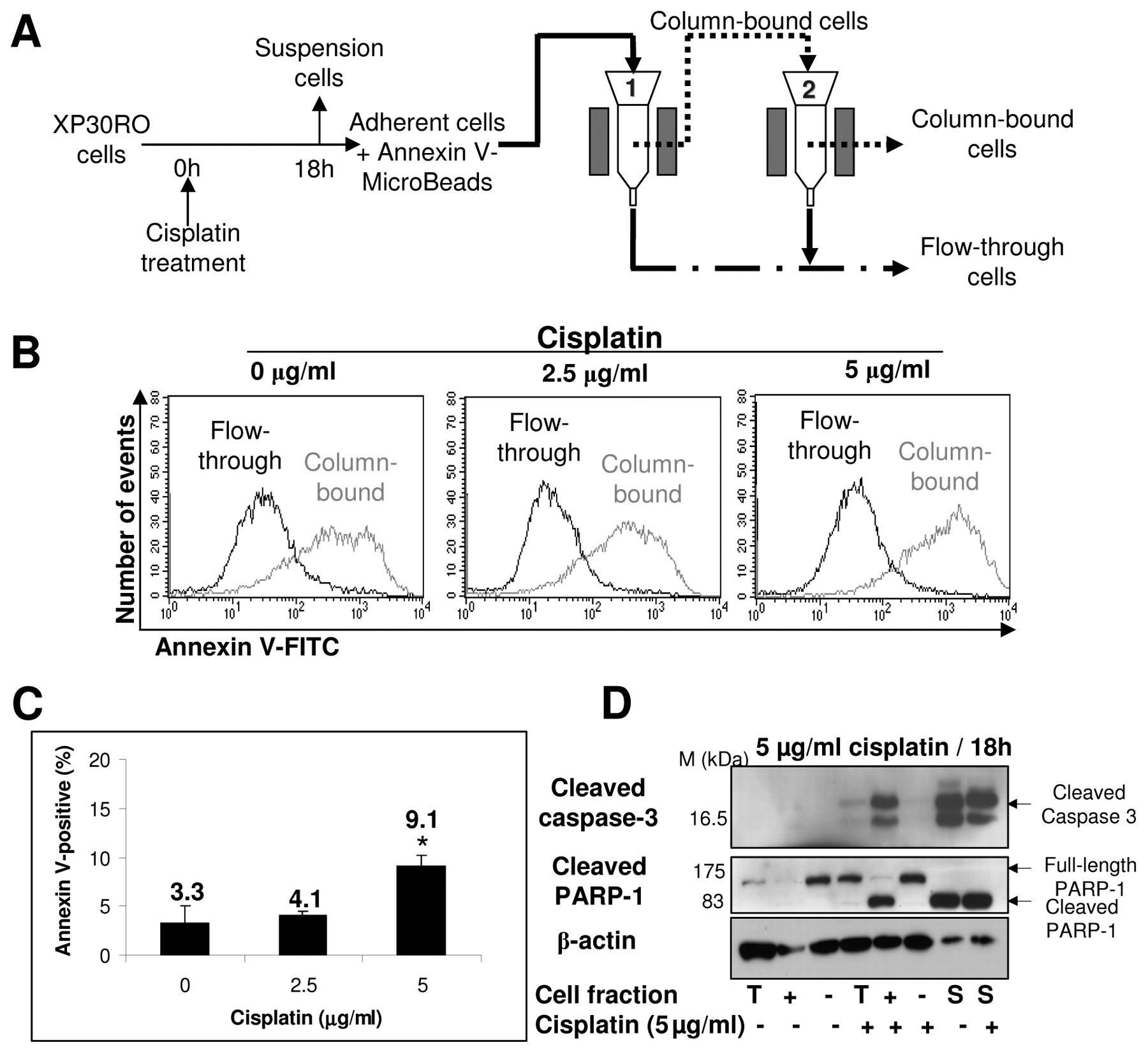
Separation of annexin V-positive and annexin V-negative cells using the annexin V-MicroBead separation system. A: XP30RO cells were treated with DMSO, 2.5 or 5 μg/ml cisplatin for 18h. Adherent cells were incubated with annexin V-MicroBeads and applied to a MACS® column. Column-bound cells were eluted and applied to a second MACS® column. Flow-through and column-bound cells were collected for further analysis. B: XP30RO cells were labelled with annexin V-FITC during separation. 10,000 events were analysed by flow cytometry for each condition. Data are representative of three experiments. C: The percentage of annexin V-positive cells following cisplatin treatment. Data are an average of three experiments; error bars represent one standard deviation. *p<0.05, as determined using ANOVA. D: Lysates from annexin V-positive and annexin V-negative XP30RO cells were analysed by Western blotting using anti-cleaved PARP-1 (which also recognises the full-length protein), anti-cleaved caspase-3 and anti-β-actin. Data are representative of three experiments. T, Total cell population; +, annexin V-positive cells; –, annexin V-negative cells; S, cells in suspension at the time of harvest.

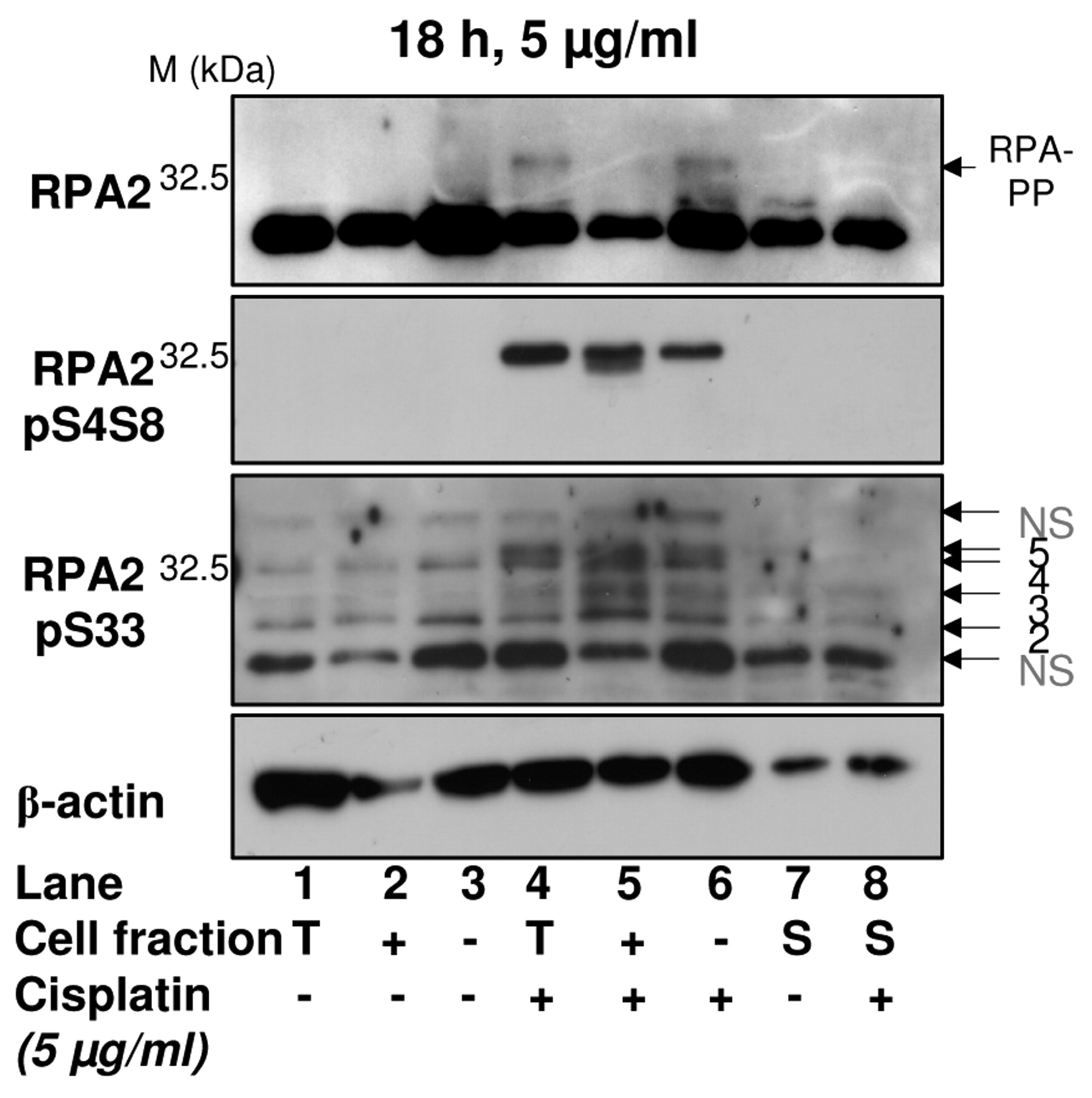
Phosphorylation of RPA2 in apoptotic and non-apoptotic cells. XP30RO cells treated with DMSO or 5 μg/ml cisplatin were fractionated 18 h later using annexin V-beads. Cell extracts were analysed by Western blotting using antibodies specific for RPA2, and RPA2 phosphorylated on Ser4/Ser8 or Ser33. Data are representative of three experiments. T, Total cell population; +, annexin V-positive cells; –, annexin V-negative cells; S, cells in suspension at the time of harvest.
Cisplatin-induced phosphorylation of RPA2 differs in annexin V-positive and -negative cells. To determine whether annexin V-based MACS® was able to reveal differences in cisplatin-induced protein phosphorylation in apoptotic and non-apoptotic XP30RO cells, Western blotting with phosphospecific antibodies was used to examine a known DNA damage response, namely hyperphosphorylation of RPA2. Following DNA damage, the RPA2 subunit of the trimeric single-stranded DNA binding protein RPA is phosphorylated in a PIKK-dependent manner on multiple N-terminal sites, including Ser4/Ser8 and Ser33 (8, 9, 21). Cisplatin induced phosphorylation of RPA2 on Ser4/Ser8 and Ser33 in XP30RO cells (Figure 2, lanes 1-3 vs. lanes 4-6). However, the pattern of RPA2 phosphorylation differed between annexin V-positive and annexin V-negative cell extracts (Figure 2, lanes 5 and 6). Following separation of XP30RO cells, the level of the slow mobility hyperphosphorylated form of RPA2 (9) was lower in annexin V-positive cells compared to the annexin V-negative cells, as detected using anti-total RPA2 antibody (Figure 2, upper panel, lane 5 and 6). This might result from a lower abundance of total RPA2 in annexin V-positive cell extracts. To investigate the phosphorylation state of RPA2, phosphospecific antibodies against RPA2 pSer4/Ser8 and pSer33 were used. One band, corresponding to RPA2 phosphorylated on Ser4/Ser8, was detected in extracts of cisplatin-treated annexin V-negative cells, while in annexin V-positive cells, two forms of phosphorylated RPA2 were detected (Figure 2, second panel, lanes 5 and 6). The failure to detect both forms of RPA2 in extracts of unfractionated cisplatin-treated cells may be due to the small proportion of apoptotic cells present (Figure 1C), such that this form is only detectable in extracts enriched for apoptotic cells using MACS®. Phosphorylation of RPA2 on Ser33 was slightly higher in extracts of apoptotic cells, particularly for phosphorylated forms 2 and 3 (Figure 2, third panel, lane 5 vs. lane 6). Little phosphorylated RPA2 was detected in suspension cells using any of the anti-RPA2 antibodies (Figure 2, lanes 7 and 8). Thus, separation of XP30RO cells into annexin V-positive and annexin V-negative fractions revealed differential cisplatin-induced phosphorylation of the key DDR protein RPA2 in extracts of the two subsets of cells.
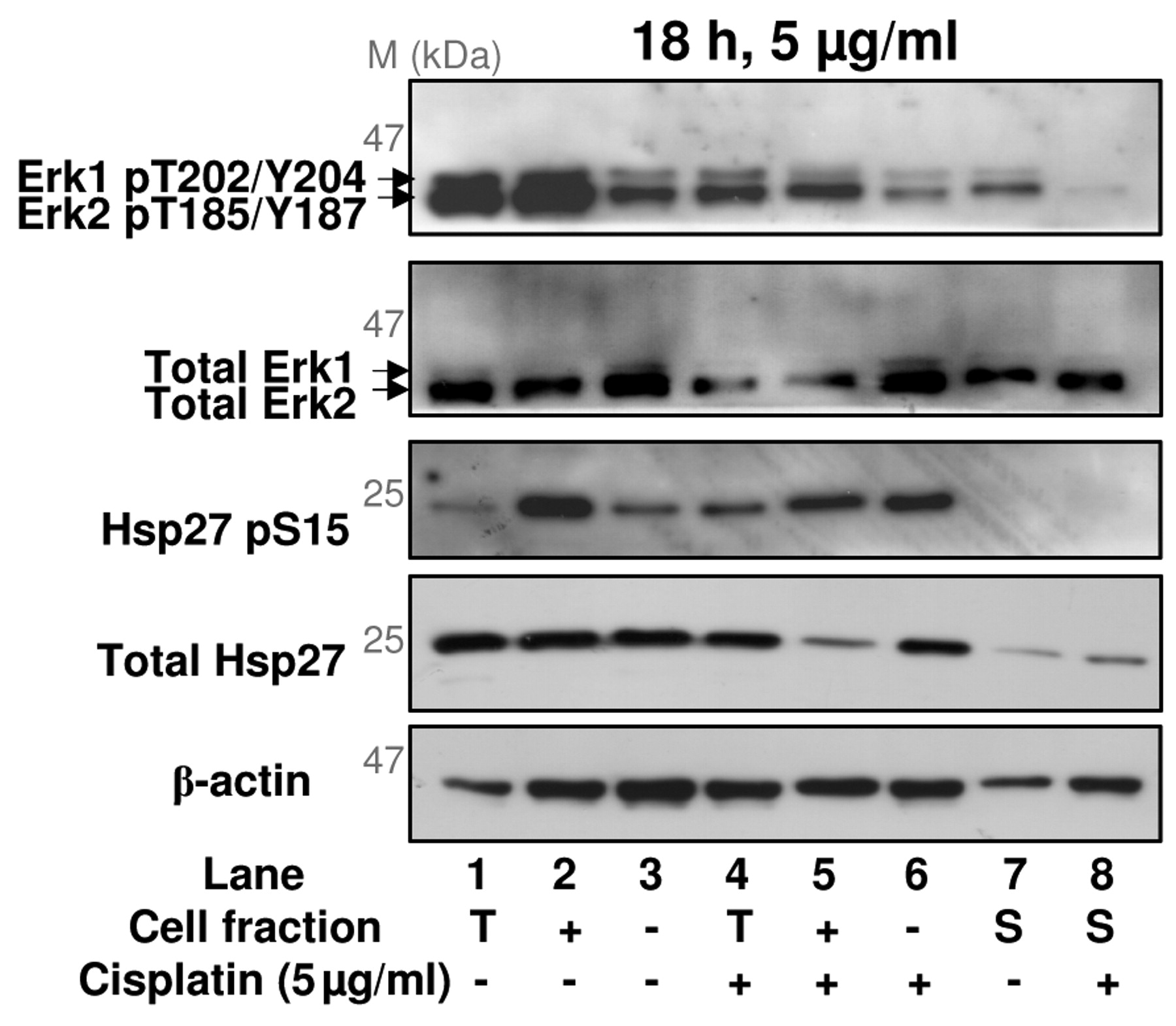
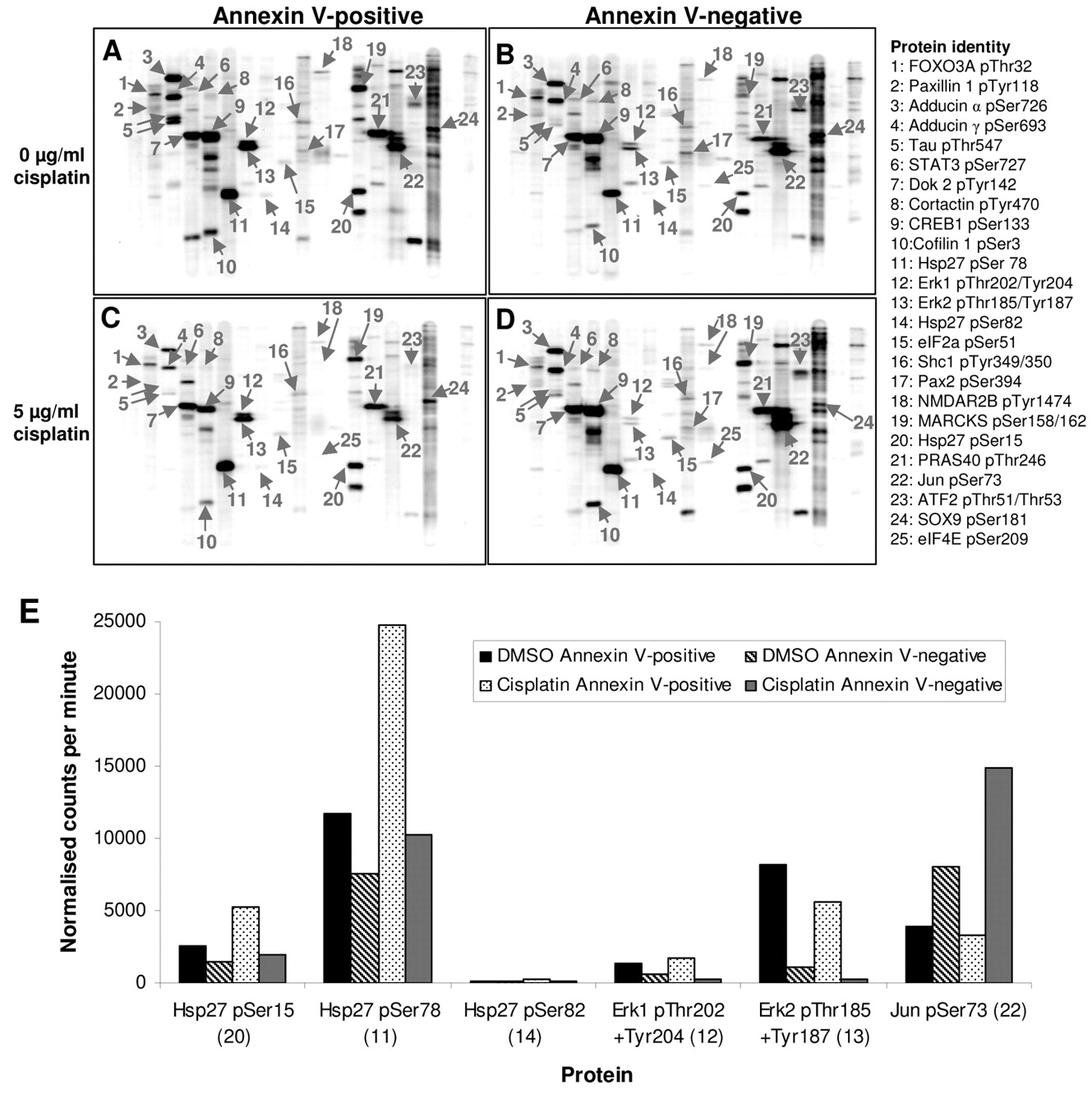
Characterization of protein phosphorylation in annexin V-positive and annexin V-negative cells, using cell separation and multi-antibody immunoblotting. The cellular response to cisplatin involves a number of cell signalling pathways in addition to PIKK-mediated protein phosphorylation (22). To provide insight into cisplatin-induced cell signalling in apoptotic or non-apoptotic cells, multi-antibody screening was used to analyse the phosphorylation state of an array of proteins from key signalling pathways, including components of the p38 MAPK, Erk, JNK, PKB/Akt, PKA and PKC pathways. Whole cell lysates of annexin V-positive and annexin V-negative cells, prepared 18h after treatment with DMSO or 5 μg/ml cisplatin, were analysed using the Kinetworks™ Substrates of Kinase multi-antibody immunoblotting screen (KPSS12.1). Of 40 phosphorylation sites on 33 different proteins examined, 25 phosphorylation events were detected in the cell lysates (Figure 3A-D). Quantification of individual phosphoprotein bands and signal normalisation between immunoblots was carried out, allowing the intensity of individual bands to be compared between different cell extracts. Following normalisation, a number of phosphoproteins that are differentially phosphorylated in a cisplatin- or cell fraction-dependent manner were identified, including Hsp27, Erk1, Erk2 and Jun (Figure 3E).
The 23 kDa chaperone protein heat shock protein 27 (Hsp27) is a downstream target of the p38 MAPK pathway, and is phosphorylated by MAPK-activated protein kinase 2 (MAPKAPK2) on Ser15, Ser78 and Ser82 in vitro and in vivo (23, 24). MACS® revealed that Hsp27 phosphorylation on Ser15 (Figure 3A-D, Band 20) and Ser78 (Figure 3A-D, Band 11) is higher in annexin V-positive XP30RO cells than in annexin V-negative cells (Figure 3E), in particular following cisplatin treatment. In contrast, phosphorylation of Hsp27 on Ser82 (Figure 3A-D, Band 14) was barely detected in either cell fraction, and was not induced by cisplatin treatment (Figure 3E).

Kinetworks™ multi-antibody immunoblot analysis of annexin V-positive and annexin V-negative cell lysates. XP30RO cells were treated with DMSO or 5 μg/ml cisplatin and fractionated as described in Figure 2. Cell lysates from three independent experiments were pooled and analysed using multi-immunoblotting, to probe for 33 individual phosphoproteins (Kinetworks™ Substrates of Kinase screen). The resulting immunoblots for each sample are shown. Annexin V-positive fraction of untreated (A) and cisplatin-treated (C) cells. Annexin V-negative fraction of untreated (B) and cisplatin-treated (D) cells. Specific phosphoprotein bands are labelled 1-25 and listed beside the immunoblots. E: Quantification of band intensity from multi-immunoblots of specific phosphoproteins. The normalised trace quantity of specific phosphoprotein bands is shown in normalised counts per minute. The number beside each protein corresponds to the band number in Figure 3A-D.
The extracellular signal-regulated protein kinases Erk1 (43 kDa) and Erk2 (41 kDa) translocate to the nucleus on activation, and up-regulate expression of proteins involved in cell proliferation and cell cycle progression (25). Phosphorylation of Erk1 on Thr202 and Tyr204, and Erk2 on Thr185 and Tyr187 is carried out by MEK1 or MEK2, and results in Erk activation (25). Overall, Erk1 phosphorylation on Thr202 and Tyr204 was low in XP30RO cell extracts under the conditions tested (Figure 3E, band 12). However, following separation of apoptotic and non-apoptotic cells, Erk1 phosphorylation on Thr202 and Tyr204 was found to be higher in annexin V-positive cells than in annexin V-negative cells (Figure 3E, band 12). An overall stronger signal was detected for Erk2 phosphorylated on Thr185/Tyr187 (Figure 3, A-D, band 13). In this case also, the extent of Erk2 phosphorylation on Thr185/Tyr187 was greater in annexin V-positive cells, with a slight decrease in both fractions following cisplatin treatment. In contrast, phosphorylation of the transcription factor Jun on Ser73 was higher in annexin V-negative cells, and was increased following cisplatin exposure (Figure 3E).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
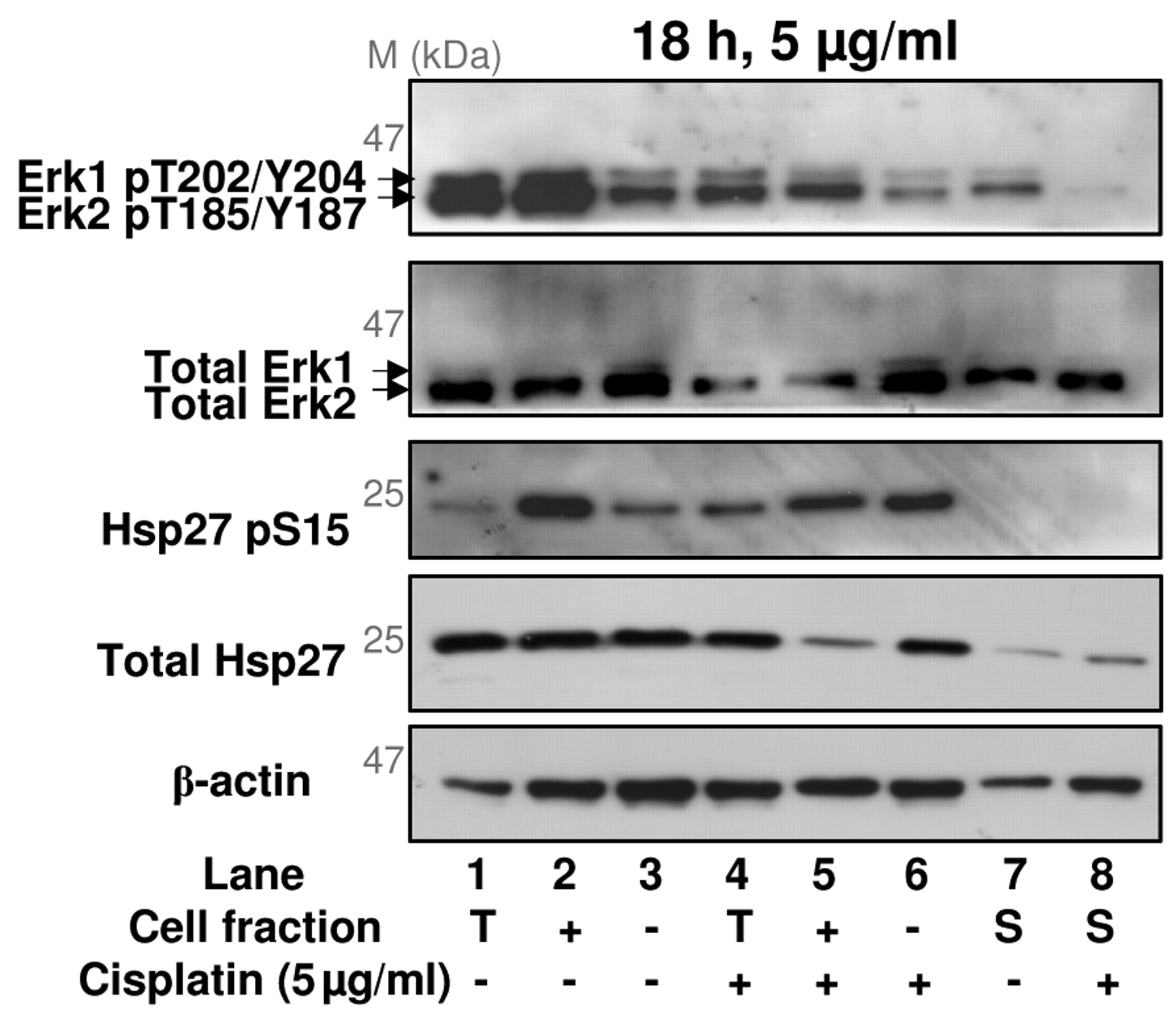
Western blot analysis of Erk1, Erk2 and Hsp27 phosphorylation in apoptotic and non-apoptotic XP30RO cells. Extracts from cells treated and fractionated as described in Figure 2, were analysed by Western blotting for Erk1 pThr202/Tyr204, Erk2 pThr185/Tyr187, total Erk1/Erk2, Hsp27 pSer15 and total Hsp27. Data is derived from one experiment. T, Unfractionated (total) cells; +, annexin V-positive cells; –, annexin V-negative cells.
To validate the changes in phosphorylation observed in extracts of XP30RO cells separated by annexin V-based MACS®, phosphorylation of Erk1, Erk2 and Hsp27 was analysed by Western blotting of protein extracts, independently prepared from cisplatin-treated XP30RO cells. Consistent with the results of the multi-antibody screen, the majority of Erk1 phosphorylated on Thr202/Tyr204, and Erk2 phosphorylated on Thr185/Tyr187 was found in annexin V-positive cell extracts rather than in annexin V-negative cell extracts (Figure 4, upper panel). Furthermore, Erk2 phosphorylation on Thr185/Tyr187 was lower in extracts of apoptotic cisplatin-treated cells (Figure 4, lane 2 vs. lane 5).
Hsp27 phosphorylation on Ser15 was greater in annexin V-positive cells than in annexin V-negative cells (Figure 4, third panel). The overall level of Hsp27 protein was reduced in extracts of cisplatin-treated annexin V-positive cells compared to extracts of annexin V-positive cells derived from untreated cells (Figure 4, fourth panel, lanes 2 and 5). However, the level of phosphorylation was similar in both cell fractions, indicating that phosphorylation of Hsp27 is greater in annexin V-positive cells. These data were consistent with the results of the multi-antibody screen (Figure 3E, bands 13, 20).
Discussion
The present study described a MACS®-based approach to isolation of apoptotic and non-apoptotic cell populations, and highlighted the importance of investigating signalling pathways in each cell population. A number of lines of evidence indicated that annexin V-positive XP30RO cells isolated using annexin V-based MACS® are apoptotic, including data from flow cytometry using annexin V-FITC (Figure 1B) and the presence of cleaved caspase-3 and PARP-1 in cell extracts (Figure 1D).
Cisplatin-induced protein phosphorylation was characterised following MACS®-based separation. RPA2 hyperphosphorylation was detected in annexin V-positive cells but not in suspension cells, demonstrating that DNA damage-induced protein phosphorylation can still be detected in apoptotic cells isolated using this separation system. Isolation of apoptotic cells and subsequent screening by Western blotting revealed differences in protein phosphorylation and abundance in each subset of cells, which could not be identified by analysing extracts of the total cell population. For example, distinct phosphorylated forms of RPA2 were present in extracts of apoptotic and non-apoptotic cells, and the total level of Hsp27 protein differed between the two cell populations.
Integration of the separation system with multi-antibody immunoblotting revealed that a number of cellular signalling proteins including Erk1, Erk2, Jun and Hsp27 are differentially phosphorylated in apoptotic and non-apoptotic cells and in response to cisplatin. Protein lysates from at least three independent experiments were combined and then analysed by multi-immunoblotting to generate the data shown here. Independent demonstration that Erk1, Erk2 and Hsp27 phosphorylation shows a similar pattern when analysed by Western blotting as when analysed by the Kinexus screen supports the use of the multi-immunoblotting approach. This approach allows phosphorylation of specific proteins to be integrated with the outcome of exposure to cisplatin.
The functional significance of apoptotic cell-specific phosphorylation events detected here requires further investigation. Specific phosphorylated forms of RPA2 were detected in apoptotic cells following cisplatin (Figure 2). Individual phosphorylation events, such as those that generate multiple forms of RPA2 phosphorylated on Ser4/Ser8, may play a role in apoptosis in response to DNA damage, as phosphorylation of RPA2 on Ser4/Ser8 is required for efficient apoptosis in response to HU (26).
Erk1 and Erk2 phosphorylation is detected predominantly in apoptotic XP30RO cells, independent of cisplatin treatment, supporting a pro-apoptotic role for phosphorylated Erk1 and Erk2 under the conditions described here. However, given evidence that Erk1 and Erk2 can have either pro-apoptotic (27, 28) or anti-apoptotic effects (29-31), depending on the cellular context, further investigation of the role of Erk1/2 phosphorylation in cisplatin-mediated apoptosis is required.
Phosphorylation of Hsp27 on Ser15 and Ser78 was detected primarily in apoptotic cells following cisplatin treatment (Figure 3), suggesting a role for Hsp27 phosphorylation on these residues in apoptosis. Phosphorylation on Ser82 of Hsp27 was low in all cell fractions (Figure 3), indicating that Hsp27 pSer82 is not involved in cell fate following cisplatin treatment, and that phosphorylation on Hsp27 Ser82 may be carried out by a separate kinase. Hsp27 can be phosphorylated on Ser82 by Akt in vitro and in vivo (32), however, the functional significance of cisplatin-induced Hsp27 phosphorylation in apoptosis requires further elucidation.
In conclusion, the MACS®-based approach described in this study allows separation of apoptotic and non-apoptotic cells following cisplatin treatment in a human cell line, and subsequent analysis of cellular responses to cisplatin. Combination of MACS® with multi-antibody screening revealed that Erk1/2 and Hsp27 are differentially phosphorylated in apoptotic cells. This approach may provide insights into the relationship between protein phosphorylation in response to cisplatin, and the outcome of exposure to this chemotherapeutic drug.
Acknowledgments
This research was supported by an Irish Research Council for Science Engineering and Technology postgraduate scholarship (E.O'M.), and in part by a Science Foundation Ireland basic research grant (M.P.C., S.C.-H.) and by the NUI, Galway Beckman fund (E.O'M.). We thank Dr. Adrienne Gorman for antibodies against ERK-1 and -2, and Hsp27.
- Received August 27, 2010.
- Accepted September 10, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved