

Abstract
Activated leucocyte adhesion molecule (ALCAM) was originally identified as a transmembrane receptor which is involved in T-cell activation and has other still unresolved functions in hematopoiesis, development, inflammation and transendothelial migration of neutrophils. ALCAM is a member of a subfamily of immunoglobulin receptors with five immunoglobulin-like domains (VVC2C2C2) in the extracellular domain and is expressed in many types of tumors. The tumor-type-dependent impact of its expression level with respect to prognosis points to a possible context-dependent function. Most functional investigations have focused on malignant melanoma, in which high ALCAM expression at the protein level correlates with a poor prognosis. ALCAM mediates low-affinity homophilic interactions and much stronger interactions with CD6. Modulation of ALCAM function with agents such as transfected dominant negative ALCAM and ligand-binding secreted ALCAM both lead to inhibition of matrix metalloproteinase-2 activation, but their impact with respect to invasion in vitro and metastasis in vivo are different. Single-chain Fv fragments directed against ALCAM are efficiently internalized, paving the way for exploration of immunoconjugates as therapeutic agents. Validation experiments of the target with modulatory agents for possible therapeutic application in oncology are discussed.
- Adhesion and aggregation
- dissemination and metastasis
- homo- and heterophilic interactions
- malignant melanoma
- prognostic marker
- target validation
- therapeutic intervention
- review
As described in a landmark paper by Hanahan and Weinberg (1), the hallmarks of cancer are limitless replicative potential, sustained angiogenesis, evasion of apoptosis, self-sufficiency with respect to growth signals, insensitivity to anti-growth signals, tissue invasion and metastasis. Recently, cancer-related inflammation was added as the seventh hallmark of cancer (2). Adhesion molecules are involved in all of the hallmark-related processes. They can be categorized into immunoglobulins, cadherins, selectins, integrins and mucins, and are involved in cell cell adhesion, interaction with the extracellular matrix, growth of the primary tumor and its vascularization, local invasion, dissemination by intra- and extravasation, and the formation of metastases via the blood or lymphatic system in distant organs. Therefore, modulation of the function of adhesion molecules is a strategy with potential impact for intervention in the progression of cancer. Many of the cancer-related adhesion molecules are involved in cancer pathophysiology as well as in inflammation. In this review, we focus on the immunoglobulin-like adhesion molecule activated leukocyte cellular adhesion molecule (ALCAM) and discuss its involvement in the pathogenesis of different types of tumors. We also outline translational aspects regarding therapeutic intervention.
ALCAM – General Features
ALCAM is a member of the immunoglobulin superfamily and was identified by expression cloning based on its ability to bind to CD6 making use of COS cells transfected with c-DNA libraries (3, 4). ALCAM is able to mediate homophilic as well as heterophilic (CD6) interactions.
The human gene for ALCAM is located on chromosome 3 (3q13.1q13.2), is composed of 16 exons and has a size of over 200 kb. ALCAM is a type I transmembrane molecule with an extracellular domain of 500 amino acids, a transmembrane domain of 22 amino acids, a short cytoplasmic domain of 34 amino acids and a molecular mass of 105 kDa (69 kDa after deglycosylation). ALCAM consists of five extracellular Ig domains (VVC2C2C2) and was identified on activated leukocytes, hematopoietic stem cells and myeloid progenitors. Topology and amino acid sequence of human ALCAM are shown in Figures 1 and 2. Besides expression on hematopoietic cells, ALCAM is expressed on neuronal cells, mesenchymal stem cells, bone marrow stromal cells and hematopoiesis supporting osteoblastic cells. ALCAM expression was described during organ develop-ment in the central and peripheral nervous system, sensory organs, hematopoiesis, endothelial and epithelial lineage (all reviewed in 5, 6). The functional contribution of ALCAM is still poorly resolved.
ALCAM becomes rapidly expressed upon polyclonal stimulation of resting T-cells in vitro, reaching a maximum after 3 days of culture and decreasing to undetectable levels at day 8 (7). A role of ALCAM in hematopoiesis is indicated by its expression in a subpopulation of CD34+ bone marrow cells, as well as the surrounding bone marrow stromal cells (8). ALCAM also was identified as an adhesion molecule on metastasizing melanoma cell lines (9). ALCAM orthologs are expressed in many species and have many designations such as ALCAM/CD166 ligand, MEMD, SB10 antigen and HCA in humans; HB2, KG-CAM and F84.1 antigen in the rat; MuSC in the mouse; DM GRASP, SC1 and BEN in chicken; and Neurolin and E21 antigen in the zebrafish (5, 6). The amino acid sequences of human, chimpanzee, rhesus, mouse, rat, chicken, xenopus and zebrafish ALCAM are displayed in Figure 3. The amino acid identity score of the human ALCAM protein versus the ortholog proteins as specified is 100, 99, 93, 93, 74, 57 and 39%, respectively.
Besides CD166, two paralogs containing the VVC2C2C2 motif have also been described: MCAM/MelCAM (CD146/MUC18) and basal cell adhesion molecule/Lutheran blood group antigen (B-CAM/Lutheran) (10). Other possible paralogs are advanced glycosylation end product-specific receptor (AGER) (11) and the proteins cell adhesion molecules 2, 3 and 4 (CADM-2, -3 and -4) (12-14). The amino acid sequences of the paralogs are displayed in Figure 4. Amino acid identity (similarity) score of ALCAM versus these paralogs as specified is 22.7 (41.7), 23.7 (38.5), 19.4 (29.9), 15.3 (28), 15.9 (27.7) and 16.1 (27)%, repectively. Analysis with respect to conserved residues (cysteine and tryptophan), as well as of the gene structures (start of exons and phase of introns) was performed to evaluate their ancestry. Despite the lack of some Ig-like domains in AGER and CADM-4, the nearly perfect conservation of cysteines and tryptophans and the very high conservation of exon start positions and even, in most cases, the phase of the introns argue in favor of a common ancestor for this group of genes.
A splice variant of ALCAM encoding the single amino-terminal Ig-like domain of ALCAM was identified in endothelial cells (15) (Figure 2). Both ALCAM and ligand-binding secreted ALCAM (sALCAM) were shown to be expressed by a variety of cultured endothelial cells and were differentially regulated in human microvascular endothelial cells (HMEC) upon tumor-necrosis factor α stimulation. Functional effects of ALCAM were assessed by over-expression studies. ALCAM enhanced tube formation in endothelial-like yolk sac cells on matrigel, whereas it inhibited their migration in vitro. sALCAM completely abolished these effects. sALCAM also enhanced migration of endothelial yolk sac cells that do not express ALCAM, indicating that sALCAM has an ALCAM-independent effect on cell migration in addition to modulating ALCAM function. sALCAM is purported to bind to ALCAM and thus to inhibit ALCAM-ALCAM homophilic interactions. These findings would indicate that sALCAM probably has ALCAM-dependent and ALCAM-independent functions.

Topology of ALCAM. The two Ig-like V-type domains and the three Ig-like C2-type domains are shown in red and green; glycosylation sites and the cytoplasmic domain are indicated.
Identification of ALCAM as an Oncology-related Target
Several genomics- and proteomics-based approaches have pinpointed ALCAM as an oncology-related target. Proteomic identification of differentially expressed plasma proteins in renal carcinoma by stable isotope labeling of a von Hippel-Lindau transfectant cell model revealed up regulation and differential glycosylation of ALCAM in von Hippel defective cell lines (16). Selection of phage antibodies against tumor cells in situ on both fresh frozen and paraffin-embedded tissues resulted in identification of ALCAM as a prostate cancer marker (17). Two single-chain Fv fragments were conjugated to liposomes and it was shown that they were delivered to the interior of prostate cancer cells. Similar conclusions were obtained by combined affinity labeling, high-resolution two dimensional gel electrophoresis and mass spectrometric analysis of proteins expressed on the surface of normal and prostate cancer cells (18). Matched pairs of mRNAs isolated from micro-dissected malignant and benign prostate tissues revealed ALCAM as a differentially expressed gene and as a new prognostic marker for patient survival (19). ALCAM was revealed as a N-glycoprotein bearing β 1-6 branched oligosaccharides on the A375 melanoma cell line by tandem mass spectrometry (20). It is well known that metastatic potential is associated with increased (GlcNAc) β 1-6 branching of N-glycans in tumor cells. ALCAM has been identified by several groups as an antigen which is shared by colorectal cancer stem cells (21-23). An increased expression of stem cell markers CD166 (ALCAM), CD133 and nestin during progression of malignant melanoma was described (24).

Amino acid sequence of ALCAM and splice variant V1. Relevant domains are indicated by a colour code as specified, the first amino acids of ALCAM exons are indicated.
Homophilic ALCAM-ALCAM interactions
The affinity of the homophilic ALCAM-ALCAM interaction (Kd=29-48 μM, koff ≥5.3 s-1) is two orders of magnitude lower than that of the heterophilic human ALCAM CD6 interaction (Kd=0.4-1 μM, koff ≥0.4-0.63 s-1) (25). A structure function analysis using two monoclonal ALCAM antibodies and a series of amino-terminally deleted ALCAM constructs revealed that homophilic cell adhesion depends on ligand binding mediated by the membrane distal amino-terminal immunoglobulin domain and on avidity controlled by ALCAM clustering at the cell surface involving membrane-proximal immunoglobulin domains (26). The homophilic interaction may be strengthened by cis-oligomerization of ALCAM on the cell surface and network formation on apposing cells. This reinforcement may increase avidity, compensate for the relatively low affinity constant and contribute to stabilizing homophilic cell-cell interactions. The resulting model predicts that the two membrane distal V-type Ig loops (domains V1 and V2) form a ligand-binding module and mediate receptor trans-trans-interactions between apposing cells, whereas the three membrane proximal C2-type Ig-loops (domains 3 to 5) form an oligomerization module involved in cis-clustering on the cell surface. The extracellular structures of ALCAM thus provide two structurally and functionally distinguishable modules, one involved in ligand binding and the other in avidity. Coexpression of an ALCAM deletion mutant which lacks the ligand-binding domain (domains 1 and 2), ΔN-ALCAM (domains 3 to 5), and endogenous wild-type ALCAM (domains 1 to 5) inhibited homophilic cell cell interactions by interference with ALCAM avidity, while homophilic, soluble ligand binding remained unaltered. Functionality of both modules is required for stable ALCAM ALCAM cell cell interaction. Deletion of the V domain (V1) not only disrupted homophilic ALCAM-mediated cell cell interaction but also prevented binding of soluble ALCAM-Fc (domains 1 to 5). Coordinate oligomerization and ligand binding leads to the formation of a tight bilayered ALCAM-network, enabling stable adhesive interactions. The cytoskeleton was shown to regulate ALCAM-mediated cell adhesion because inhibition of actin polymerization by cytochalasin D (CytD) strongly induces homotypic ALCAM-ALCAM interactions (27). Induction of cell adhesion is likely due to clustering of ALCAM on the cell surface which is observed after CytD treatment. It was shown that the lateral mobility of ALCAM is increased 30 fold after CytD treatment. In contrast, both the surface distribution and adhesion of glycophosphatidyl-inositol (GPI)-anchored ALCAM mutant are insensitive to CytD, despite an increase in lateral motility of GPI-ALCAM upon CytD treatment, demonstrating that clustering of ALCAM is essential for cell adhesion, whereas enhanced diffusion of ALCAM alone is not sufficient for cluster formation.

Amino acid sequences of ALCAM orthologs. Ig-like V-type, Ig-like C2-type domains and the transmembrane domains are indicated by a colour code as specified.

Amino acid sequences of ALCAM paralogs. Gene evolution-relevant parameters are indicated as specified.
Activation of ALCAM-mediated adhesion is regulated through actin cytoskeleton-dependent clustering. The link between ALCAM and the cytoskeleton has not yet been established. Possible candidates are the ezrin-radixin-moesin (ERM) family of proteins that link transmembrane molecules to the actin cytoskeleton by binding to the positively charged amino acids clusters in the cytoplasmic domain of a number of associated proteins (28). Unlike adhesion mediated by integrins and cadherins, ALCAM-mediated adhesion is independent of Rho-like GTPase activity such as RhoA, Rac1 and CDC42 (29). By contrast, it was shown that protein kinase Cα (PKCα) plays a major role in ALCAM-mediated adhesion. PKC inhibition by chelerythrine chloride and myristoylated PKC pseudosubstrate strongly reduce cytoskeleton dependent ALCAM-mediated adhesion. It can be ruled out that ALCAM is a direct PKC substrate since ALCAM is not phosphorylated, and serine as well as threonine residues are dispensible for ALCAM-mediated adhesion. ALCAM-mediated adhesion requires both the disruption of the cytoskeleton and the presence of active PKCα. Furthermore, it was shown that E cadherin and ALCAM are coordinately recruited to cell cell contacts by α-catenin (30).
ALCAM-CD6 Interaction
CD6 was identified as a heterophilic ligand for ALCAM. CD6 is expressed on T-lymphocytes, thymocytes and a subset of B cells as a surface receptor (31). The extracellular region of CD6 contains three scavenger-receptor cysteine-rich (SRCR) domains, thus including CD6 into the SRCR superfamily of protein receptors (32). Several studies show a role for CD6 as a co-stimulatory molecule in T-cell activation (32). It has been shown that the amino-terminal Ig-like domain of ALCAM binds to the membrane proximal SRCR domain of CD6 with a 1:1 stoichiometry (33) and that these interactions play a role in mediating the binding of thymocytes to thymic epithelial cells and of T-cells to activated leukocytes. Both ALCAM and CD6 are actively recruited at the T-cell–APC (antigen-presenting cell) interface and contribute to stabilization of the immunological synapse. It was demonstrated that ALCAM-CD6-mediated adhesion contributes to both early and later stages of dendritic cell induced T-cell activation and proliferation (34). ALCAM-CD6 interactions play a pivotal role in stabilizing T-cell-APC contacts. CD6 has a dual function in that it facilitates stable adhesion and acts as a co stimulatory molecule. Once the antigen-specific T-cell receptor–major histocompatibility complex antigen (TCR-MHC) contact has been established, CD6 is rapidly recruited to the T-cell–APC interface where it stabilizes the immunological synapse by interacting with ALCAM. Subsequent CD6-modulating events result in a strong and sustained co-stimulatory signal that is essential for T-cell proliferation. It has been demonstrated that ALCAM-CD6 interaction results in activation of three mitogen-activated protein kinase (MAPK) cascades, extracellular signal-regulate kinases 1 and 2 (ERK1/2), p38 and c-Jun N-terminal kinase (JNK) (35). Agents interfering with ALCAM function should therefore be evaluated with respect to their immunosuppressive properties.

ALCAM mRNA expression in melanoma, esophageal and bladder cancer tissues and in corresponding non-transformed tissues. Comparison of ALCAM expression signals between tumor and normal tissue for (A) skin, (B) esophagus and (C) bladder. Gene expression data were derived from the GEO database (skin [GSE3189], esophagus [5364], bladder [GSE3167] and n depicts the number of biological samples within each group. CIS, Carcinoma in situ; sTCC, superficial transitional cell carcinoma; mTCC, muscle invasive transitional cell carcinoma.

ALCAM neighourhood analysis. Ingenuity pathway analysis (IPA) was used to create the neighborhood of genes for ALCAM. All molecules with a reported relationship to ALCAM are shown. Edges are annotated by the relationship type of two molecules: PP, Protein-protein interaction; E, expression; LO, location; PD, protein-DNA interaction; A, activation; RR, RNA-RNA interaction. All neighboring molecules of ALCAM were manually grouped and annotated according to their molecular type. MIRs, MicroRNAs; LDLR, low density lipoprotein receptor; L1CAM, L1 cell adhesion molecule; ERK1/2, mitogen-activated protein kinase 1/3; CLRN1, clarin 1; ARHGDIG, Rho GDP dissociation inhibitor (GDI) gamma; CYTH3, cytohesin 3; STK11, serine/threonine kinase 11; EGFR, epidermal growth factor receptor; MYC, myelocytomatisis viral oncogene homolog; YY1, YY1 transcription factor; EGF, epidermal growth factor; CD40L, CD40 ligand; TNF, tumor necrosis factor; CCL5, chemokine (C-C motif) ligand 5; CD6, CD6 molecule; NGFR, nerve growth factor receptor.
ALCAM Expression as a Prognostic Marker in Human Cancer
Immunohistochemistry-based analysis of a series of human melanocytic lesions reveals that ALCAM expression correlates with melanoma progression (36). Most naevi (34 out of 38) and all thin melanomas (Clark levels I and II) did not express ALCAM. Immunoreactivity was detected in the invasive vertical growth phase of 2 out of 13 Clark level III lesions, in 13 out of 19 Clark level IV lesions, and in 4 out of 4 Clark level V lesions. Thirteen out of twenty-eight of metastatic melanoma lesions were ALCAM positive. ALCAM expression was observed in fewer than 10% of the lesions that were thinner than 1.5 mm and in over 70% of lesions that were thicker than 1.5 mm. This study revealed ALCAM as an important marker of melanoma progression. An independent investigation revealed expression of ALCAM in 11/71 (15% of nevi), 37/70 (53%) of primary melanomas and 58/84 (69%) of metastatic melanomas (37). As shown in Figure 5A, based on data derived from databases, at the RNA level the opposite was observed, with highest steady-state ALCAM mRNA levels in melanocytes and lower levels in nevi followed by malignant melanoma. Apparently, regulation of expression of ALCAM might occur at the RNA level.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
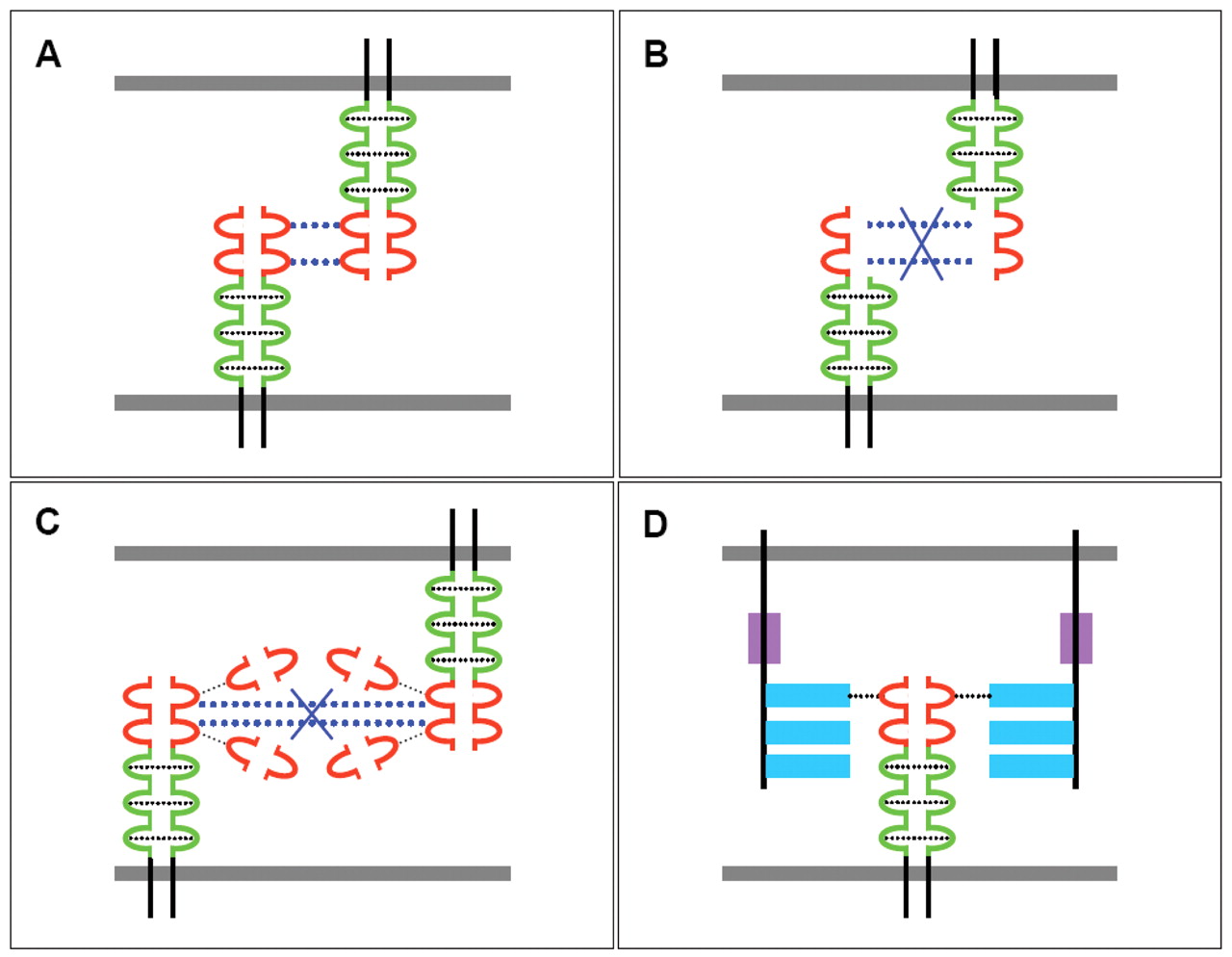
Schematic display of ALCAM-related interactions on the cell surface and with adjacent cells. A: Homophilic ALCAM-ALCAM interactions on the cell surface mediated by the C-domains and between adjacent cells mediated by the V-domains. B: Impact of interactions between ALCAM and ΔN-ALCAM (domains 3 to 5) with respect to adhesive interactions. C: Interference of secreted V1 domains with ALCAM-ALCAM interactions between adjacent cells. D: ALCAM CD6 interaction. ALCAM V and C domains are shown in red and blue, green bars and stippled light blue bars indicate adhesion between V and C domains. The scavenger-receptor cysteine-rich and the membrane proximal stalk region are shown as filled boxes in blue and magenta. The plasma membrane is depicted as a filled dark blue box.
Analysis of ALCAM expression of 107 oral squamous cell carcinomas, 78 oral lesions (58 hyperplasias and 20 dysplasias) and 30 histologically normal oral tissues identified cytoplasmic accumulation of ALCAM as a predictor of poor prognosis (38). ALCAM expression was observed in 42/65 of human esophageal squamous cell carcinomas and in 17/25 (68%) of dysplasias compared to paired distant esophageal tissues (39) at the RNA and protein levels. These findings indicate that ALCAM expression occurs at a premalignant state (dysplasia) and correlates with enhanced tumor invasion and metastasis.
As shown in Figure 5B, we were able to confirm increased steady-state levels of ALCAM mRNA based on data derived from databases in esophageal carcinomas compared to corresponding matching non-transformed tissues. Analysis of 109 well-characterized epithelial ovarian cancer patient samples showed that a decrease or loss of ALCAM membrane expression is a marker of poor outcome (40). A study using immunohistochemical analysis of paraffin-embedded samples from patients with pancreatic cancer concluded that increased ALCAM is an independent prognostic marker of poor survival and early tumor relapse (41). Fifty-two bladder carcinomas were analysed for ALCAM expression by immunohistochemistry and ALCAM expression was found to be strongly associated with a poor prognosis in grade 3 tumors (42). As shown in Figure 5C, analysis of steady state ALCAM mRNA level in databases revealed increased expression of ALCAM mRNA in bladder carcinoma compared to matching non-transformed bladder tissues. A total of 111 colorectal carcinomas were screened for membranous and cytoplasmic expression of ALCAM and in survival analysis, membranous ALCAM expression correlated significantly with shortened patient survival (43). Analysis of RNA from 54 matched pairs of microdissected prostate tissue (tumoral and normal) showed ALCAM mRNA up regulation in low grade prostate cancer and progressive loss in high-grade lesions (44). ALCAM was shown to be up-regulated in 86% of prostate carcinomas by immunohistochemistry (45). Transcript analysis of frozen breast-related tissues (normal breast=32, breast cancer=120) suggest that decreased ALCAM expression is of clinical significance in breast cancer and that reduced expression correlates with a more aggressive phenotype and poor prognosis (46).
Analysis of ALCAM expression at the RNA and protein level in 162 primary mammary carcinomas resulted in the finding that high ALCAM expression levels in primary tumors might be a suitable marker for prediction of the response to adjuvant chemotherapy in breast cancer (47). A study based on immunohistochemical analysis of ALCAM in 162 primary breast carcinomas indicated that strong cytoplasmic ALCAM expression might be a marker for aggressive breast cancer (48). ALCAM expression was assessed by RNA analysis and immunohistochemistry in 234 frozen primary breast cancer tissues and 34 non-neoplastic mammary tissues. The study showed that patients who develop skeletal metastasis tend to have the lowest levels of ALCAM transcripts in their breast cancer tissues (49). Another study reported that serum ALCAM appears to be a biomarker for breast cancer and may have value for disease diagnosis (50).
Taken together the impact of ALCAM expression on prognosis seems to depend on the tumor type, in some types of cancer, membranous versus cytoplasmic expression of ALCAM should be considered and the impact of heterologous proteins associating with ALCAM is an unresolved issue because the identification of proteins of this category is an ongoing effort (51).
Oncology-related Functional Issues
ALCAM was shown to be localized specifically at cell cell junctions in endothelial cells indicating its role as a potential anti-angiogenesis target (52). Transendothelial migration of activated monocytes was blocked by ALCAM antibodies supporting the role of ALCAM in this process. The findings support the concept that ALCAM is a component of cell junctions and a member of the molecular machinery controlling diapedesis.
The role of ALCAM shedding was investigated in a panel of ovarian carcinoma cell lines (53). It was shown that ALCAM is shed from epithelial ovarian cancer cell lines in vitro by the metalloproteinase A disintegrin and metalloproteinases (ADAM17) (TACE) leading to the generation of soluble ALCAM, consisting of most of the extracellular domain of ALCAM comprising the ligand-binding domain and the oligomerization domain. A similar molecule was found in ascites and sera of patients, suggesting that a similar process occurs in vivo. The process was enhanced by epidermal growth factor, pervanadate and phorbol 12-myristate 13-acetate and was be inhibited by pharmacologic inhibitors of matrix metalloproteinases (MMP) and ADAM. ADAM17/TACE was expressed by epithelial ovarian cancer cell lines and silencing of ADAM17/TACE by RNAi reduced shedding. Inhibitors of ADAM function blocked the motility of epithelial ovarian cancer cell lines in a wound-healing assay. Conversely, an antibody blocking ALCAM adhesive function and internalization enhanced the motility of epithelial ovarian carcinoma cells. A similar role in ovarian carcinoma motility has been assigned to L1CAM which can be released by the combined action of ADAM17/TACE and ADAM10 (54). These findings need to be extended with patient-derived specimen with known clinical history.
ALCAM-mediated cell-to-cell adhesion was shown to contribute to MMP-2 activation (55), which involves formation of an intermediate ternary complex of membrane type 1 MMP (MT1-MMP/MMP14), tissue inhibitor of metalloproteinase 2 (TIMP2) and pro-MMP2 at the cell surface. BLM melanoma cells transfected with a dominant-negative ALCAM mutant (ΔN-ALCAM) showed inhibition of MMP-2 activation, but increased invasive behaviour, suggesting that these properties should be less dependent on MMP-2 activity. These findings would support the role of ALCAM as a cell density sensor and initiator of a signal toward MMP-2 activation. These observations are not restricted to metastatic BLM melanoma cells, they also have been shown in ALCAM-positive human fibrosarcoma HT1080 cells. Other cell adhesion molecules might perform similar functions as ALCAM. The extracellular MMP inducer EMMPRIN/CD147 is highly expressed in tumor cells and induces adjacent fibroblasts to express MMPs. Ectopic expression of ΔN-ALCAM in melanoma cells induces loss of cellular aggregation and increased cell mobility in skin reconstructs. In addition an acceleration of spontaneous lung metastasis formation was observed at the expense of primary tumor growth in mouse xenografts (56).
In order to elucidate ALCAM function in more detail, two monoclonal antibodies were analysed (26): mAb J4-81, which specifically recognizes domain 1 (V1), and AZN L50, which maps to an epitope of the membrane proximal domains 4 and 5. mAb J4-81 enhanced homophilic ALCAM-mediated cell adhesion and aggregation, possibly by inducing a conformational change that promotes ligand binding. This is in contrast with the inhibitory effect of mAb J4-81 on the heterophilic ALCAM CD6 interaction. The inhibitory effect of mAb AZN-L50 on ALCAM-mediated cell adhesion and aggregation indicated that domains 4 and 5 are essential for homophilic ALCAM-ALCAM interaction.
As previously outlined in this review, using a series of progressively truncated ALCAM mutants, it was shown that the amino-terminal domain 1 (V1) is critically involved in the homophilic interaction (26). Expression of ΔN-ALCAM (domains 3 to 5), which lacks the ligand-binding function, reduced cell aggregation, but not soluble ligand binding in the wild-type ALCAM-expressing BLM cells. Since the epitope of the ALCAM-blocking mAb AZN L-50 is localized in domains 4 and 5, it is tempting to speculate that the inhibitory effect of this mAb is a result of prohibited lateral oligomerisation of ALCAM molecules by steric hindrance. This notion is supported by the observation that mAb AZN-L50 does not affect soluble ligand binding and therefore only modulates ALCAM avidity.
The secreted variant of ALCAM (sALCAM) containing the V-type domain 1 (V1) confers ligand binding and attenuates invasion of the highly metastatic BLM melanoma cells (57). Overexpression of sALCAM in metastatic melanoma cells disturbed clustering of endogenous ALCAM and inhibited activation of MMP-2. In contrast to the promotive effects of ΔN-ALCAM, sALCAM impaired the migratory effect of transfected cells in vitro, reduced basement-membrane penetration in reconstituted human skin equivalents and diminished metabolic capacity in nude mice. sALCAM presumably blocks homophilic receptor ligand binding. Mimicry of limited or enhanced ALCAM recruitment by overexpression of sALCAM or ΔN-ALCAM directly correlates with attenuated or enhanced metastatic capacities. L1CAM/CD171, another marker of progression for several types of cancer, including melanoma, was suppressed upon sALCAM overexpression but was up regulated by ΔN-ALCAM. Whereas ectopic ΔN-ALCAM mainly increased the invasive behaviour, sALCAM which is anticipated to prevent ligand binding (15), attenuated the formation of the intercellular ALCAM network and inhibited melanoma migration in vitro and lung metastasis in nude mice. These are contradictive effects of sALCAM and ΔN-ALCAM on melanoma invasion and metastasis, however, both approaches modulating ALCAM functionality impaired the activation of pro-invasive MMP-2. The partially overlapping and opposite effects induced by alternative strategies targeting ALCAM function suggest that ALCAM fulfills a role in melanoma progression beyond its adhesive function. ALCAM-related interactions based on Ingenuity data are summarized in Figure 6. ALCAM-related interactions such as homophilic interactions, consequences of a dominant-negative ALCAM mutant and ALCAM splice variant V1, as well as interaction of ALCAM with CD6 are summarized in Figure 7 A, B, C and D, respectively.
ALCAM as a Target for Antibody-related Approaches
Making use of an internalizing single-chain variable fragment (scFv) directed against ALCAM and conjugated to immunoliposomes, cytotoxic payloads (topotecan, vinorelbine and doxorubicin) were delivered to prostate cancer cells (DU-145, PC3, LNCaP) (58). Immunoliposomal topotecan was the most effective in cytotoxicity assays in all three tumor cells. Liposomal doxorubicin was highly effective against LNCaP, but not PC3 or DU-145 cells, despite efficient intracellular delivery. Post-internalization events thus modulate the efficacy of intracellularly delivered liposomal drugs. Internalized scFv directed against ALCAM was converted into full-length human IgG1 and shown to retain its internalization properties in hormone-resistant prostate cancer cell lines (59). Independently, a scFv fragment directed against ALCAM was isolated by panning a phage display library of human Fv fragments on IGROV-1 human ovarian carcinoma cells (I/F8 scFv) which cross-react with murine, monkey and human ALCAM indicating that the epitope is highly conserved (60). I/F8 scFv induces ALCAM internalization and co-localises with clathrin, but not with caveolin. Endocytosed ALCAM was shown to recycle back to the cell surface. I/F8 scFv-saporin immunotoxin was efficiently delivered into tumor cells and shown to kill ALCAM expressing tumor cells selectively. ALCAM-directed antibodies were generated with a mimotope eliciting GD2 ganglioside cross-reactive IgG antibody responses as well as MHC class I-restricted CD8+ cells to syngeneic neuroblastoma cells (61). The mimotope was generated by screening a phage display peptide library with anti-GD2 mab 14G2a.
Translational Aspects
As outlined, ALCAM is expressed in many types of tumors, but it is also broadly expressed in normal tissues, which may raise toxicity issues if this target is hit by agents such as mAbs, immuno-conjugates or small molecules. It has been shown that it is possible to generate cross-reactive antibodies (mouse, rat, monkey, human) (60, 62), allowing target-guided toxicology investigations. The impact of ALCAM expression on different types of tumors is clearly documented; however, in some types of tumors, high, in others low expression of ALCAM correlates with a poor prognosis. Further complicating parameters such as membranous versus cytoplasmic expression have to be considered. This would suggest that the prognostic impact of ALCAM expression is context dependent. With the exception of CD6, heterophilic ALCAM-interacting proteins have not yet been described. It has been shown that the actin cytoskeleton and PKCα are involved in mediating homophilic ALCAM-ALCAM interactions and one report refers to MAPK activation (35) induced by ALCAM ALCAM interaction, but in general, ALCAM-related signaling is poorly resolved and needs further investigations also in the context of identification of biomarkers reflecting target modulation. Inhibition of MMP-2 activation after inhibition of ALCAM-ALCAM interaction may point in this direction. The impact of ALCAM modulation by mAbs covering different epitopes and RNAi needs to be systematically investigated in a panel of tumor cells with defined ALCAM status regarding their impact on proliferation, apoptosis and invasion. Inhibition of ALCAM-ALCAM interactions mediated by sALCAM and ΔN-ALCAM resulted in a paradoxical outcome. ΔN-ALCAM mediated increased invasion in vitro, inhibition of growth of xenografts, but increased metastasis, whereas sALCAM inhibited in vitro invasion, as well as metastasis of xenografts (55 – 57). Due to the low affinity ALCAM-ALCAM interaction, a genetically engineered sALCAM with improved affinity might be considered as an agent for in vitro and in vivo target validation experiments. Monoclonal antibodies have been identified which interfere with oligomerisation of ALCAM (domains 4/5 epitope-related) and others which inhibit ligand-binding (V1 epitope related) (26). Although investigated in BLM and HT1080 cells, a more comprehensive evaluation in a panel of tumor cells (melanoma-derived with different degrees of ALCAM expression and cell lines derived from other tumor types) with in vitro assays and xenograft models should help to gain a clearer picture regarding the usefulness of mAbs for treatment of ALCAM-positive tumor types. The contribution of ALCAM modulation versus immune-effector cell based efficacy (antibody-dependent cytotoxicity and complement-dependent cytotoxicity) with respect to inhibition of tumor growth and metastasis in xenograft models should also be investigated. It has been shown that anti-ALCAM sc-Fv fragments are efficiently internalized by ALCAM-positive tumor cells and cytotoxic payload carrying anti ALCAM sc-Fv immunoliposomes are able to kill tumor cells with a clear dependency of efficacy on the cell line investigated. A sc Fv saporin conjugate targeting ALCAM was identified as a cytotoxic agent (60). Due to the broad expression of ALCAM, preclinical in vivo experiments need to be designed which challenge the issue of a possible therapeutic window. In addition, the processing of ALCAM by ADAM17/TACE and its impact on prognosis and physiological function of ALCAM in ovarian carcinoma and other types of tumor remain to be investigated in more detail. The relationship between L1CAM and ALCAM with respect to co-expression in tumors and their possible reciprocal modulation by ALCAM inhibitory agents such as sALCAM and ΔN-ALCAM also needs further clarification.
- Received May 31, 2010.
- Revision received July 5, 2010.
- Accepted July 8, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved