

Abstract
Background: Resveratrol, a phytochemical present in grapes, berries, and red wines, has been reported to induce apoptosis in various cancer cells. To explore the molecular mechanisms involved in the anticancer activity, the apoptotic activity of resveratrol in hepatic cancer cells was investigated. Materials and Methods: 1-Dimensional (1D) and 2-dimensional (2D) polyacrylamide gel electrophoresis and mass spectrometry analysis were used to determine proteomic expression profiles in SK-Hep-1 cells. Results: Resveratrol inhibited cell proliferation, generated reactive oxygen species, and caused DNA single-strand breaks. 2D gel electrophoresis showed one up-regulated protein (Ras-related protein Rab 37) and five down-regulated proteins (annexin A8, thymidine kinase, maspin, peroxiredoxin-2, and guanine nucleotide-binding protein). Most of the proteins obtained from the 2D gel electrophoresis were identified as apoptosis-related proteins. From the 1D gel electrophoresis analysis, 14 proteins were identified which had no matched peptide sequence in the controls at the level of even one unique peptide unit. Resveratrol regulated the expression of proteins involved in the redox pathways and apoptosis. Conclusion: Resveratrol causes hepatic cancer cell death by suppressing the expression of antioxidant proteins.
- Resveratrol
- 2-dimensional gel electrophoresis
- reactive oxygen species
- apoptosis
- proteomics
- SK-HEP-1
- hepatic cancer cells
Regular consumption of fruits and vegetables plays an important role in the prevention of many diseases (1). Indeed, abundant epidemiological data show that high consumption of plant products reduces the incidence of several types of cancer (2). These beneficial effects of plant-rich diets have been attributed to a large intake of different groups of phytochemicals. A number of phytochemicals exert growth inhibitory effects on diverse cancer cell lines such as breast, prostate, oral, liver, and colon (3, 4).
Resveratrol, a phenolic phytochemical found in grapes and red wine, has been shown to prevent cell proliferation and induce apoptosis in various cancer cell lines (5, 6). Studies have shown that the cytotoxicity of resveratrol towards human colon cancer cells is accompanied by cell cycle arrest in the S/G2-phase (7). In human prostate cancer cell lines, resveratrol has been investigated for its ability to prevent growth and cause cell cycle arrest and apoptosis (8). Extensive in vitro and in vivo data from human cancer cells support a role for resveratrol in potent chemoprevention.
Studies have shown that the anticancer effects of phenolic compounds arise from their prooxidant properties, which cause oxidative DNA damage and apoptosis (9). Polyphenol-induced reactive oxygen species (ROS) accumulation modifies DNA bases, breaks DNA strands (10, 11), and contributes to other forms of DNA damage (12, 13). Resveratrol possesses prooxidant activity, and its 4-hydroxystilbene structure generates free radicals (14).
Numerous investigations into the precise mechanisms underlying the anticancer properties of resveratrol have been conducted. However, the exact mechanism is not fully understood, and proteomic research is lacking.
Orally administered resveratrol has a strong affinity for the liver, where it is metabolized and accumulates (15). Moreover, a 14C-labelled resveratrol distribution assay in mice showed that the liver had the highest accumulation of resveratrol. In addition, Lancon et al. studied the kinetics of resveratrol uptake by hepatic cells in vitro and found that the polyphenol had not caused cytotoxicity or hepatocyte lysis after 30 min of exposure (16). These findings support a role for resveratrol in the chemoprevention of liver cancer. Herein the proteomic expression profiles produced by resveratrol in human hepatic cancer SK-HEP-1 cells were determined using 1-dimensional (1D) and 2-dimensional (2D) polyacrylamide gel electrophoresis (PAGE) and mass spectrometry (MS) analysis to clarify the mechanism underlying the anticancer effect of resveratrol. Resveratrol-induced cytotoxicity, ROS generation and DNA damage and the effect of resveratrol on caspase-3 activity in SK-HEP-1 cells were also examined.
Materials and Methods
Reagents. Resveratrol, MTT, DMSO, PBS, and 2',7'-dichlorofluorescein diacetate (DCF-DA) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). The human hepatic cancer cell line SK-HEP-1 was purchased from the Korean Cell Line Bank.
Cell viability assay. Cell viability was determined by the conventional MTT assay. Cells were pre-incubated in a 96-well plate for 24 h, at a density of 1×106 cells /well To evaluate dose- and time-dependent cell death, the cells were treated with different doses of resveratrol (50, 100, 150, 200, 250 μM) for 24 or 48 h. The treated cells were cultured in medium containing 0.1% DMSO and resveratrol, whereas the medium of the control cells lacked the polyphenol. After incubation, the cells were treated with MTT solution for 1 h. After loading, the medium containing the MTT solution was carefully removed and DMSO solution was added. The plate was vibrated for 30 min and absorbance at 570 nm was measured. Cell viability was also assessed using the trypan blue dye exclusion assay.
Cell cycle determination. Because cell cycle deregulation and apoptosis are closely related, the effects of resveratrol on cell cycle distribution were examined using a flow cytometry assay. The SK-HEP1 cells were treated with 225 μM resveratrol for 24 h. At the end of treatment, the cells were detached from the plates by the addition of 0.25% trypsin. The cells were then washed in PBS, fixed in 70% ethanol at 4°C and treated with 10 mg/mL RNAse for 30 min at 37°C. The DNA content was determined in an FACS flow cytometer after staining the cells with 50 μg/mL propidium iodide for 15 min in the dark. The number of cells in each cell cycle phase was determined using ModFit software (Verity Software House Inc., Topsham, ME, USA).
Comet assay. The comet assay was used to detect DNA strand breaks, and it was performed according to the manufacturer's (Trevigen, Gaithersburg, MD, USA) protocol. Slide reading was performed immediately with an Axiovert 200 (Zeiss Inc. Oberkochen, German) at ×1000 magnification, after staining with Hoechst 33342 solution (20 μg/mL, 50 μL/slide). Fluorescence was measured at an excitation wavelength of 480 nm and an emission wavelength of 520 nm.
Reactive oxygen species detection. A fluorometric assay was used as a probe for ROS in order to assess whether resveratrol promoted oxidative stress in the SK-HEP-1 cells. After 24 h resveratrol treatment, 50 μM DCF-DA were loaded. Medium containing DCF-DA was incubated for 10 min at 37°C in the dark. After probe loading, the cells were washed with Hank's solution and observed by fluorescence microscopy. Oxidation causes the intracellular formation of fluorescent DCF, a marker of ROS accumulation, and a consequent change in fluorescence. Fluorescence was measured at an excitation wavelength of 480 nm and an emission wavelength of 520 nm.
Caspase-3 assay. The effect of resveratrol on caspase-3 activity in SK-HEP-1 cells was analyzed by measuring chrometric p-nitroanilide (pNA), which is formed by cleavage of Ac-Asp-Glu-Val-Asp (DEVD)-pNA by caspase-3. Caspase-3 activity was assessed using an Apo-Alert colorimetric caspase assay kit (Calbiochem, San Diego, CA, USA). The cells were treated with or without 225 μM resveratrol for 24 h, then collected and lysed in lysis buffer and an equal amount of cell lysate protein was added to the reaction mixtures containing labeled caspase-3-specific substrate, DEVD-pNA. After 30 min incubation, caspase-3 activity was measured by changes in absorbance at 405 nm. The increase in caspase-3 activity caused by resveratrol was determined by comparing the absorbance of treated and control samples.
1D and 2D SDS-PAGE. Cells were treated with 225 μM resveratrol and incubated for 24 h in the same culture conditions. Treated and control cells were washed with ice-cold PBS and lysed in buffer containing 10 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% Triton X-100, 0.5% Nonidet P-40, 1 mM EDTA, 1 mM ethylene glycol tetraacetic acid (EGTA), 1 mM phenylmethanesul-phonylfluoride or phenylmethylsulphonyl fluoride (PMSF), 1 μg/mL leupeptin, and 1 μg/mL aprotinin. The protein concentration in the lysate was normalized with Bradford reagent (BioRad, Hercules, CA, USA). For 1D-PAGE, equal amounts (50 μg) of protein were separated on 10% SDS-polyacrylamide gels and stained using Coomassie Brilliant Blue (BioRad). Each lane was broken into 15 pieces and used for MS/MS analysis. 2D electrophoresis was performed as previously described (17) on the protein bands that were markedly increased or decreased in the resveratrol-treated cells by matching bands in the control and resveratrol-treated samples. The first dimensional isoelectric focusing (IEF) was performed on immobilized pH 3-10 gradient immobilized pH gradient (IPG) strips at 20°C. The second dimensional SDS-PAGE was performed in 10% acrylamide gels using a PROTEAN II 2D cell system (BioRad). After electrophoresis, the gels were visualized by silver staining.
Protein identification by mass spectrometry. After 1D-PAGE, the differential expression of proteins was analyzed using a liquid chromatography electrospray ionization quadruple time-of-flight (LC-ESI-Q-TOF) mass spectrometry (MS)/MS system. The system with reversed phase micro liquid chromatography (RP-μLC) comprised a Surveyor MS pump (Thermo Electron, San Jose, CA, USA), a Spark autosampler (Spark, Emmen, the Netherlands), and a Finnigan LTQ linear ion trap MS (Thermo Electron) equipped with NSI sources. Ten μL of tryptic digested peptides were injected. The mobile phase comprised water (eluent A) and acetonitrile (eluent B), and both contained 0.1% formic acid. The flow rate was maintained at 200 nL/min. The gradient was started at 5% B, a linear gradient to 60% B was achieved in 49 min, ramped to 80% B in 5 min, and to 100% A in the next 15 min. All the MS/MS data were searched against the International Protein Index (IPI) human protein database using the SEQUEST algorithm (Thermo Electron). From the 2D gels, differentially-expressed protein bands were excised and 25 mM ammonium bicarbonate/50% acetonitrile (1:1) solution was added. After shrinking in a dehydration buffer, the gel pieces were dried with a speed vacuum. The gel pieces were then soaked in 25 mM ammonium bicarbonate buffer containing 5 μL/mL trypsin. Mass spectra were obtained using a Voyager-DE STR matrix-assisted laser desorption ionization (MALDI)-TOF (Applied Biosystems, Cambridge, MA, USA). The matrix solution was prepared by dissolving 10 mg α-cyano-4-hydroxycinnamic acid in 1 mL of 50% acetonitrile and 0.1% trifluoroacetic acid. A matrix peak (m/z=379.0930 Da) and a trypsin fragment peak (m/z=2163.0616 Da) served as internal standards for mass calibration.
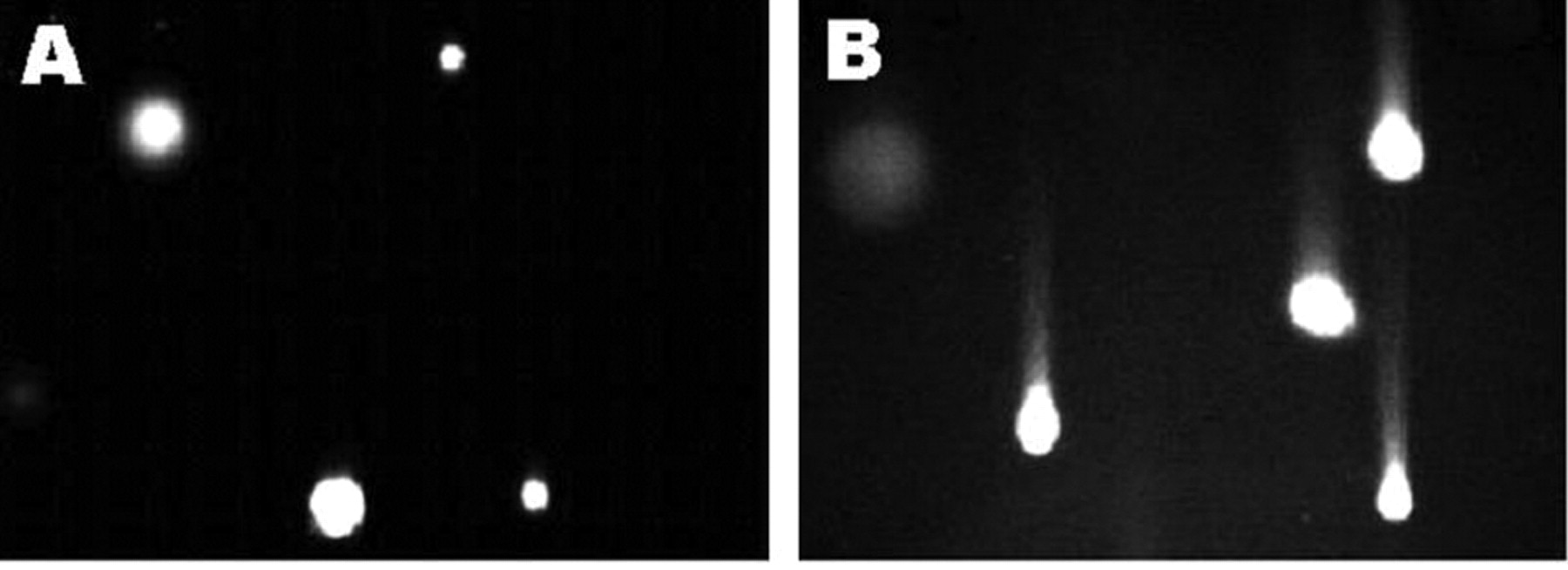
Resveratrol-induced DNA damage in SK-HEP-1 cells. DNA strand breaks were determined using the comet assay. Cells were cultured in medium containing 0.1% DMSO (A) or in the presence of 225 μM resveratrol (B). Results are representative of three independent experiments.
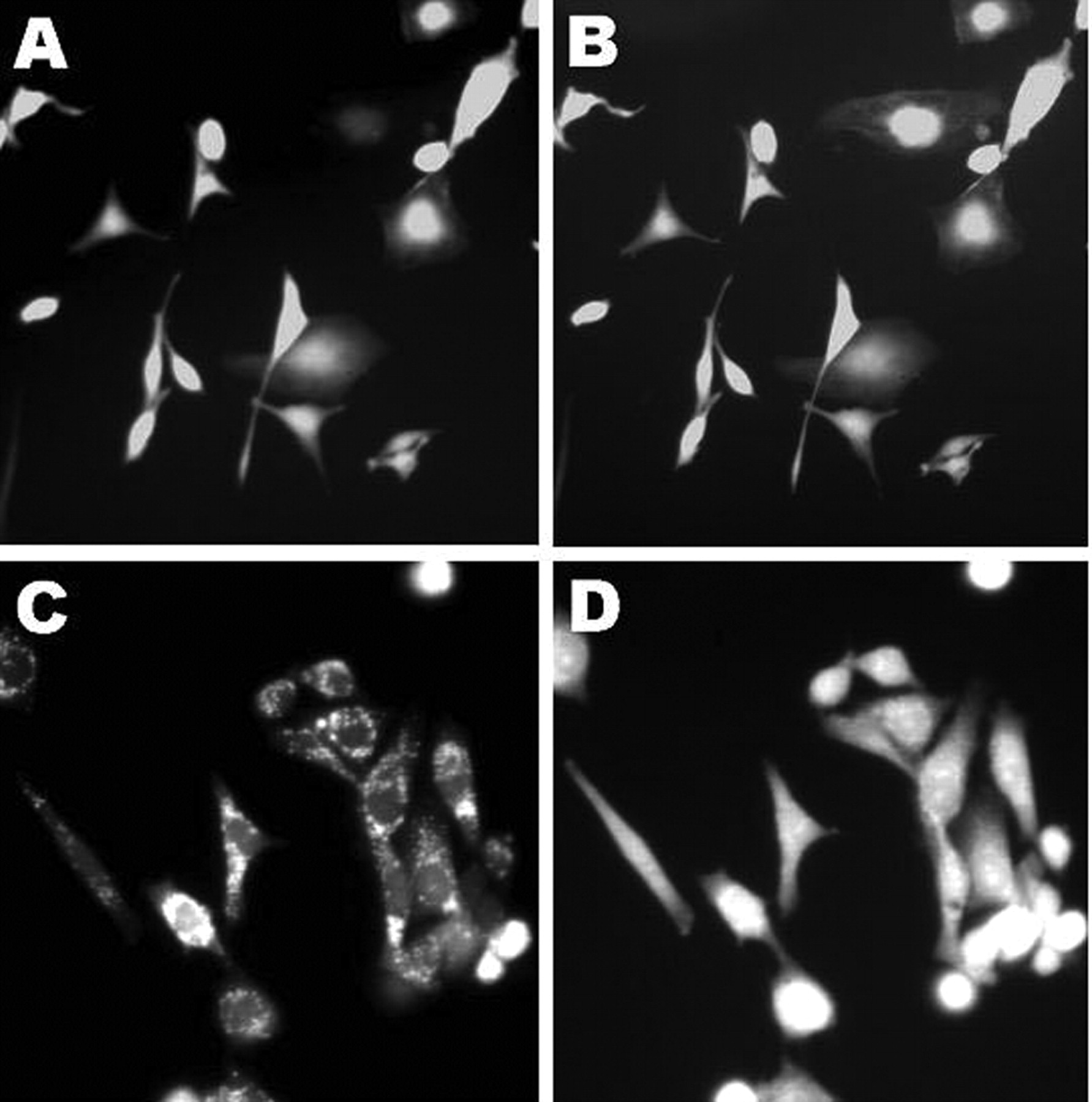
Effect of resveratrol on intracellular accumulation of reactive oxygen species (ROS). Intracellular ROS levels were determined using dichlorofluorescein fluorescence. Cells were cultured in medium containing 0.1% DMSO (A, B) or in the presence of 225 μM resveratrol (C, D). Fluorescence photomicrographs were taken at 5 s intervals in both control and resveratrol-treated groups.
Results
Effect of resveratrol on growth and cell cycle arrest. Decreased cell viability was observed with increasing concentrations of resveratrol. The polyphenol reduced the number of viable cells to 92%, 88%, 66%, 45%, and 31% of control levels after 24 h, with 50, 100, 150, 200 and 250 μM resveratrol respectively. These effects were greater after 48 h under similar conditions. Thus, resveratrol caused dose- and time-dependent cell death in the SK-HEP-1 cells. These effects were confirmed by direct cell counting using trypan blue dye exclusion.
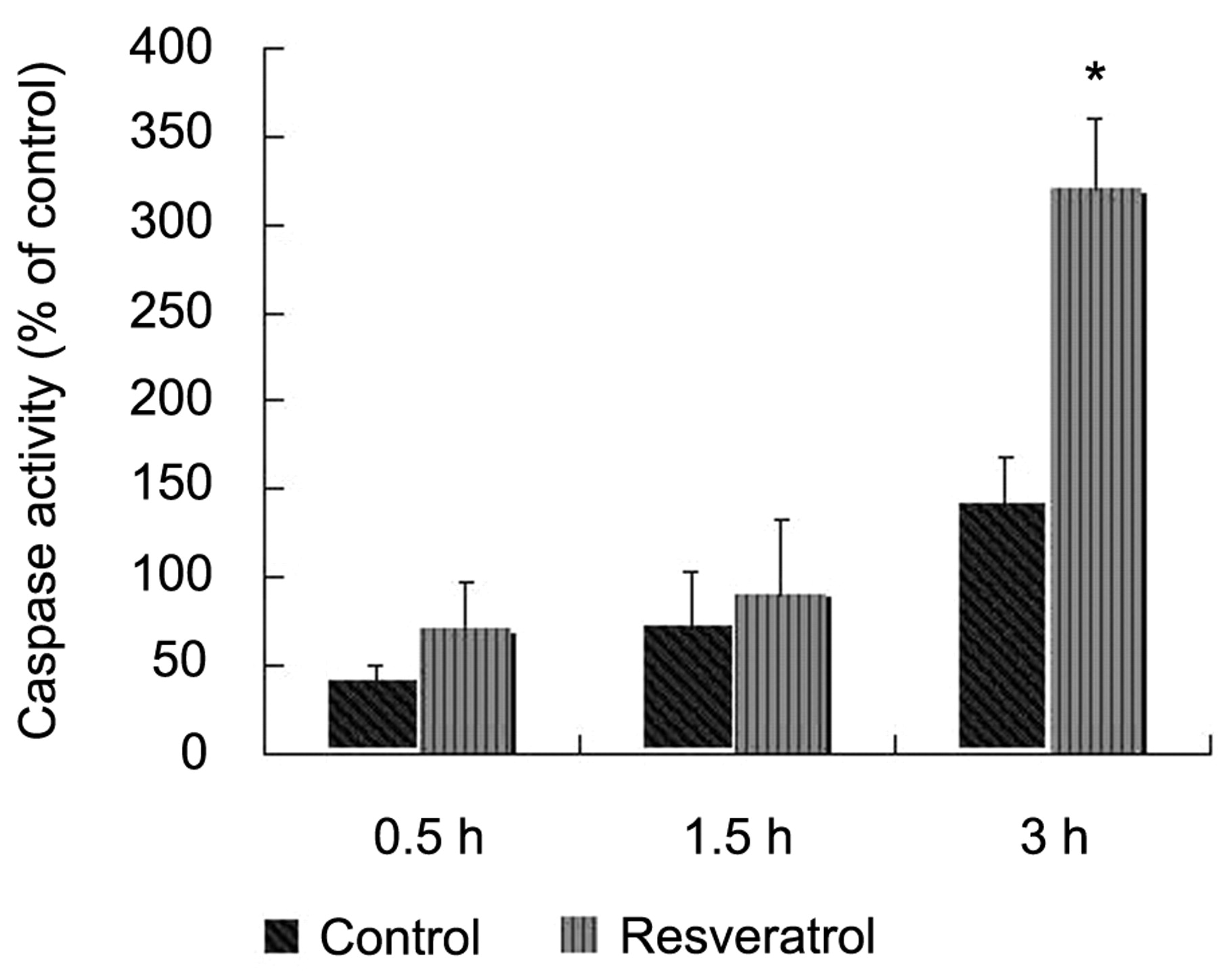
Effect of resveratrol on caspase-3 activity. Caspase-3 activity was determined at a wavelength of 405 nm using chromophore p-nitroanilide (pNA) from DEVD-pNA after 24 h treatment with or without resveratrol. Data are presented as the percent increase in caspase-3 activity in treated cells. Data are mean±SD of three independent experiments. *p<0.01 compared to control, two-tailed Student's t-test.
The number of viable cells was approximately 40% that of the control when treated with 225 μM resveratrol for 24 h. The population of cells in the G0/G1 phase increased from 11.80% to 69.36% following resveratrol treatment. Resveratrol prevented G1/S transition and induced consequent G1 phase arrest. The increase in the number of cells in G0/G1 was accompanied by a decrease in the number of cells in G2/M. The percentage of cells in the latter phase decreased from 83.31% to 14.55%.
Effect of resveratrol on apoptosis. DNA damage and intracellular ROS generation were examined as potential apoptotic phenomenon induced by resveratrol. DNA damage was detected using the comet assay. Resveratrol caused significant tail migration and intensive fluorescence compared to that seen in the control samples. Figure 1 shows representative images of control and resveratrol-treated cells in the assay. Intracellular ROS accumulation is presented as fluorescent pictures taken at intervals of 5 s (Figure 2). Compared to the control, the cells treated with 225 μM resveratrol had more intensive fluorescence.
The time-dependent analysis of caspase-3 activity in the SK-HEP-1 cells is presented in Figure 3. As shown in the figure, a stable increase in caspase-3 activity occurred up to 3 h.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
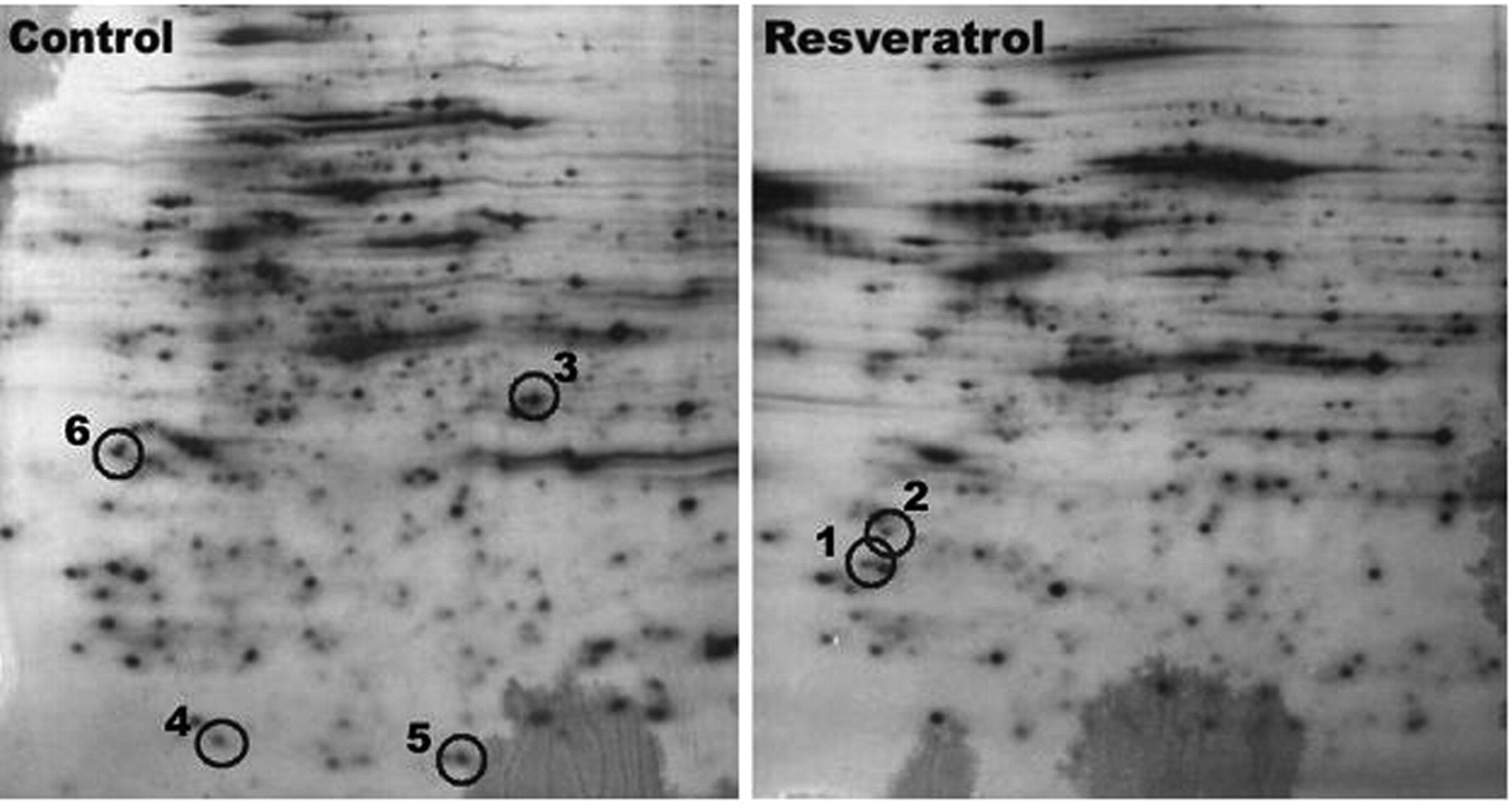
Representative images of 2-dimensional (2D) silver-stained SDS-PAGE gels. Showing the pattern of separated protein bands in resveratrol-treated or untreated cells after 24 h. The circle indicates bands that were selected for mass spectrometric analysis. Resveratrol up-regulated protein represented by band 2 and down-regulated proteins represented by bands 1 and 3 to 6.
Protein identification using 1- and 2D PAGE. Six independent sets of 2D electrophoresis were carried out on the differentially-expressed proteins identified using MALDI-TOF MS. Representative 2D gel images showing the pattern of separated protein bands in the control and resveratrol-treated cells are shown in Figure 4. The circles indicate bands that were selected for MS analysis. Among these six, resveratrol up-regulated one (Ras-related protein Rab 37) and down-regulated five (annexin A8, thymidine kinase, maspin, peroxiredoxin-2, guanine nucleotide-binding protein) (Table I).
The protein expression was further evaluated by loading the lysates from the resveratrol-treated cells onto a 1D-SDS-PAGE gel and by separating them into 15 pieces in both lanes. The whole bands from each lane were analyzed using LC-ESI-Q-TOF MS/MS. This approach revealed that resveratrol induced an overall proteomic expression of 721 proteins. By comparing the digested peptides in the control and resveratrol-treated samples, 14 resveratrol-specific proteins for which the control samples had no matched peptide sequences, even at the level of one unique peptide unit, were identified: apolipoprotein-L2, calcium-binding mitochondrial carrier protein, NADH-cytochrome b5 reductase, heme oxygenase 1, transferrin receptor protein 1, sulfide:quinone oxidoreductase, 26S proteasome non-ATPase regulatory subunit 1, dehydrogenase/reductase short-chain dehydrogenase (SDR) family member 7, retinol dehydrogenase 11, tyrosine-protein kinase CSK, chloride intracellular channel protein 5, annexin A7, glutathione-S-transferase (GST) mu 3, and p21 ras GTPase-activating protein-associated p62 (Table II).
Protein expression changes in redox pathways. Most of the proteins obtained from the 1D- and 2D-PAGE analysis are associated with apoptosis (Table I and Table II). The 2D analysis showed that resveratrol down-regulated peroxiredoxin-2 and maspin, which occurred simultaneously with a significant increase in ROS. These two proteins regulate not only apoptosis, but also redox pathways.
Identification of proteins spots from 2D- PAGE and MALDI-TOF MS.
Discussion
The present study confirmed that resveratrol causes ROS generation and accumulation and subsequent DNA single-strand breaks associated with apoptosis and illustrating the prooxidant properties of this polyphenol. Resveratrol up-regulated or down-regulated six proteins in the hepatic cancer cells (Table I). Of particular interest is the down-regulation of peroxiredoxin and maspin because they are involved in redox regulation.
Peroxiredoxin-2 plays a significant role in maintaining intracellular H2O2 levels and cellular redox potential (18-23). Its regulation of intracellular H2O2 promotes hyperproliferation of cells but is insufficient to induce apoptosis. A recent study reported that suppression of peroxiredoxin-2 expression using antisense cDNA sensitizes cancer cells to radiation-induced apoptosis (21). Cellular redox deregulation has been studied for its potential to increase caspase-3 activity and induce apoptosis. Taking this into account, the resveratrol-induced caspase-3 activation and apoptosis observed in the present study could be attributed to the peroxiredoxin-2 down-regulation.
Resveratrol also down-regulated maspin, which is a tumor suppressor that blocks multiple stages of carcinogenesis. Recently, Yin et al. (24, 25) reported that the down-regulation of maspin increased basal ROS levels and H2O2-induced ROS generation. They also found, using two yeast hybrids, that maspin protected against intracellular oxidative damage through a direct interaction with GST. Among all the major human GST isoforms, the mu isoform has been identified as a candidate maspin-interacting protein. In this study, 1D-PAGE showed that resveratrol influenced GST mu 3 expression (Table II). Tumor suppression by maspin may be attributed to GST-dependent redox homeostasis. Therefore, maspin down-regulation sensitizes cancer cells to resveratrol-induced apoptosis by generating excessive amounts of ROS.
Protein expression profile of resveratrol-induced proteins from 1D-PAGE and LC MS/MS analysis.
Several studies have shown that the prooxidant property of resveratrol requires the presence of Cu(II) (14, 26). Resveratrol has a potent metal-reducing activity, which is often related to ROS generation (27). Copper and iron are two redox active metals that catalyze free radical formation, as in the Fenton reaction. The reduction step in this process is thought to be influenced by metal-reducing polyphenolic compounds. Resveratrol up-regulated transferrin receptor 1 (Table II), and intracellular iron has been reported to regulate expression of this protein (28). The present proteomic data therefore suggested that resveratrol induced endogenous iron mobilization and subsequent ROS generation, which further indicated that the polyphenol is a prooxidant. Additionally the down-regulation of thymidine kinase activity is related to imbalances in dNTP levels and subsequent apoptosis (29) and most of the proteins obtained from the 2D-PAGE analysis are associated with apoptosis.
In the present study, resveratrol significantly inhibited cancer cell growth with marked ROS generation and DNA damage. Proteomic analysis showed that the polyphenol suppressed the expression of antioxidant proteins that protect cells from ROS or cellular oxidative damage in SK-HEP-1 cells. By repressing these proteins, resveratrol may increase the susceptibility of cancer cells to oxidative stress. The prooxidant action of resveratrol could be one of the important mechanisms underlying its anticancer effects. Because the liver contributes significantly to the bioavailability of dietary resveratrol (30), we suggest that this polyphenol is a potential anticancer compound in hepatic cancer cells. Our proteomic findings, therefore, may provide more effective and specific therapeutic targets for chemopreventive agents that promote cell cycle deregulation or apoptosis in hepatic cancer cells.
Acknowledgments
This study was supported by partial funding from Han Cell INC. (2007-1-1).
- Received July 8, 2009.
- Revision received September 4, 2009.
- Accepted September 7, 2009.
- Copyright© 2009 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved