

Abstract
KAI1, also known as CD82, is a candidate metastasis suppressor gene and has been indicated in the disease progression of certain solid tumours, including those of breast cancer. The present study aimed to investigate the importance of KAI1as a potential metastasis suppressor in breast cancer cells. MDA-MB-231 and MCF-7 sublines with different patterns of KAI1 expression were created by way of anti-KAI1 transgene or transfection of KAI1 expression construct. Cell adhesion was markedly increased in cancer cells showing increased expression of KAI1 (MCF-7KAI1EXP, p=0.021 vs. control cells), while it was significantly reduced in the KAI1 knockout subline, MDA-MB-231KAI1KO (p=0.002 and 0.0004, respectively). Significant increase of cell migration of MCF-7KAI1EXP cells (p=0.024 vs. control) and restricted motility of MDA-MB-231KAI1KO cells (p=0.003) were observed. Furthermore, MCF-7KAI1EXP cells also showed reduced cell invasion (p=0.022), while MDA-MB-231KAI1KO cell line showed a significant increase in invasion (p=0.0063 and p=0.007, respectively). KAI1 did not affect cell growth. It is concluded therefore that KAI1 plays an important role in cell adhesion, invasion and migration of breast cancer cells, in vitro, and is a potential metastasis suppressor gene in breast cancer.
- Kai-1
- CD82
- breast cancer
- invasion
- cell migration
Metastasis is the main complication in solid tumours. In recent decades research has focused on understanding the metastasis process, disease progression and to explore potential therapeutic targets. The cascade of metastasis has been found quite complicated and systematic, which encompasses the involvement of cancer cells, immune evasion, adhesion, invasion, motility, chemoattraction, cytoskeleton rearrangement, cell survival, gene rearrangement and unknown molecular factors. In breast cancer, metastasis has been frequently observed in bones, lungs and brain. With advances in the field of cancer research, a family of genes has been identified which were found to be responsible for the suppression of metastasis and are termed as metastasis suppressor genes (MSGs) (1). These genes play their role by inhibiting the spread of primary tumour cells, either by arresting them at the primary site, or their invasion, adhesion or extravasations in the surrounding environment.
The role of KAI1 as a candidate metastasis suppressor was first identified in prostate cancer cells (2). Dong et al. identified the location of the KAI1 (Kang Ai in Chinese) gene on chromosome 11p with 10 exons and 9 introns spanning around 80 kb (3). KAI1 exists in two isoforms, with 267 residues in isoform-1 and 242 residues in isoform-2 (4), playing a role in growth inhibition and invasion. As yet, no significant mutation has been identified in this gene in relation to oesophageal and breast cancer patients (5, 6). Regulation of KAI1 is co-ordinated with many transcriptional binding factors such as AP1 (activator protein 1), AP2 and p53 binding motifs on promoter region. A direct relationship between p53 protein and KAI1 upstream binding region suggests that either the loss of p53 consensus binding region or a change of p53 function (commonly observed in many types of cancer) leads to down regulation of KAI1 protein (7-9). Similarly, the antagonistic effect of β-catenins and reptins on co-activator Tip 60 also shed light on the regulatory mechanism of KAI1 protein (10).
Loss or reduced expression of KAI1 has been observed in the tissues of metastatic breast tumours when compared to benign and early stage breast tumour samples (11). Down-regulation, both at transcript and translational levels, has also been observed in relation to brain metastasis in breast cancer progression (12). We have recently shown that KAI1 expression is reduced in ductal carcinoma of the breast and that high levels of KAI1 are associated with a longer survival (13). The aim of the present study was to investigate the effect of KAI1 on growth, adhesion, invasion and cellular migration in two breast cancer cell lines, using a series of in vitro cell models, in order to elucidate the cellular impact of KAI1 on breast cancer cells.
Primer sequences used in the present study.
Materials and Methods
Cell lines and culture conditions. MDA-MB-231 and MCF-7 cell lines were purchased from the American Type Culture Collection (ATCC, Maryland, US) and maintained in Dulbecco's modified Eaglels medium (DMEM) supplemented with 10% foetal calf serum (PAA Laboratories Ltd, Somerset, UK), streptomycin and penicillin. The cells were incubated at 37°C, 5% CO2 and 95% humidity.
Generation of MCF-7 and MDA-MB-231 cells displaying altered levels of KAI1. KAI1 expression was either enhanced in MCF-7 cells, or suppressed in MDA-MB-231 cells through transfection of a pEF6/V5-His-TOPO (Invitrogen, Paisley, UK) containing either a KAI1 mammalian expression construct or ribozyme transgenes which specifically recognised and cleaved KAI1 mRNA transcript. These techniques have been widely implemented by our laboratories and are outlined elsewhere (14-17). The purified plasmids were sequenced in order to verify the presence and sequence of the construct (MWG, Milton Keynes, UK). MCF-7 cells were transfected with either the expression plasmid that had been sequence verified or an empty pEF6 control plasmid. Similarly, MDA-MB-231 cells were transfected with plasmids containing the ribozyme transgenes or an empty pEF6 control plasmid. Cells were then subjected to selection with blasticidin. MDA-MB-231 cells containing the control pEF6 plasmid were designated as MDA-MB-231Ct and those containing the ribozyme transgenes were labelled MDA-MB-231KAI1KO1 and MDA-MB-231KAI1KO2. Similarly, MCF7 cells containing the control pEF6 plasmid were labelled MCF7Ct whereas those containing the mammalian expression sequence were termed MCF7KAI1EXP1.
RNA isolation and cDNA synthesis. Total RNA was extracted from cells grown to confluence in a 25 cm2 tissue culture flask using TRI reagent purchased from Sigma (Dorset, UK). cDNA was synthesised using an Enhanced Avain Reverse Transcriptase-PCR-100 kit with anchored oligo(dt) primers (Sigma). β-Actin was analysed to assess cDNA quality. Specific KAI1 primer sets (Table I) were subsequently used to assess the efficacy of both the ribozyme transgenes and the KAI1 mammalian expression constructs in the MCF-7 and MDA-MB-231 cell sets, by RT-PCR (reverse transcriptioin polymerase chain reaction).
Quantitative-Polymerase Chain Reaction (Q-PCR). Real-time quantitative PCR was also used to assess KAI1 transcript levels present in the MCF-7 and MDA-MB-231 cells as previously reported (17). Briefly, the iCycler IQ system was used to detect and quantify the number of KAI1 transcripts in each of the control and transfected cells. Results are given as number of transcripts/μl based on an internal standard and the results were further normalised using the expression of GAPDH in these samples. The Q-PCR technique used the Amplifluor system (Intergen Inc., New York, US), Q-PCR Master Mix (ABgene, Surrey, UK) and a universal probe (Uniprimer™, Intergen) to record the fluorescence emitted by the z-sequences. Conditions for Q-PCR were: an initial 15 minute 95°C period followed by 60 cycles of 95°C for 15 seconds, 55°C for 60 seconds and 72°C for 20 seconds. Full details of primers used are given in Table I.
SDS-PAGE and Western blotting. Cell pellets were lysed with a SDS lysis buffer for 1hour. This was followed by removal of insolubles after centrifugation at 13,000 rpm. Cell lysates were later quantified using the Bio-Rad DC Protein Assay kit (Bio-Rad Laboratories, CA, USA). Denatured samples with equal quantity of protein were separated using SDS PAGE gels. Proteins were blotted onto nitrocellulose membrane, which was subsequently blocked in 10% milk and subjected to specific antibody probing. GAPDH was used as internal control. KAI1 antibody, raised in rabbit (Santa Cruz Biotechnologies, UK) and GAPDH, raised in goat (Santa Cruz Biotechnologies), were used for probing at a concentration of 1:100 and 1:500 respectively. Anti-rabbit and anti-goat secondary antibodies were obtained from Sigma and used at a 1:1000 concentration.
Immunocytochemical staining of KAI-1. A 16-well chamber slide was used for immunocytochemical staining with 30,000 cells from the entire panel of MCF-7 and MDA-MB-231 breast cancer cell lines seeded in each well separately. Chamber slides were placed in an incubator at 5% CO2 and 37°C for one day. Following fixation and blocking, cells were incubated with anti-KAI1 antibody at 1:100 dilution. Immunocytochemical staining was carried out following the protocol given elsewhere (18).
In vitro cell function assays
Growth assay for breast cancer cell lines. The growth of the breast cancer cell lines was assessed using a colorimetric-based method (19). At the desired time points, plates were fixed and stained with crystal violet. Cell density and growth were measured on a Bio-Tek EL ×800 multiplate reader (Bio-Tek Instruments Inc., Winooski, VT, USA).
Adhesion assay for breast cancer cell lines. This was based on a previously published protocol (17). In brief, Matrigel™ basement membrane matrix purchased from BD Biosciences, UK, was used as adhesion sheet at 5 μg/well. 40,000 cells were added per well. After 40 minutes incubation, the wells were vigorously washed to remove unbound cells. Adherent cells were then fixed and stained with crystal violet. Stained cells were later counted in a number of random fields under a ×40 objective.
Migration assay for breast cancer cell lines. A scratch wound of approximately 300 μm was artificially introduced into a monolayer of breast cancer cells and cell movement towards the wound was measured over 90 minutes. Images of the wound were taken at a number of time points and Optimas 6.0 software was used to determine the distance migrated as previously described (20, 21).
Invasion assay for breast cancer cell lines. Invasion assays were undertaken using inserts (ThinCert™ of 8 μm) from Greiner bio-one, Germany, in 24-well tissue culture plates. Each insert was first coated with Matrigel. After rehydration, 45,000 cells were seeded into each insert. After three days of incubation, inserts were fixed with formalin and stained in crystal violet. Cells stained with crystal violet were counted and observed under the microscope for possible invasion as described elsewhere (15).
Statistical analysis. Experimental procedures were repeated independently at least three times. In all assays the transfected breast cancer cell lines were compared with pEF6 plasmid controls (cells containing closed pEF6 plasmid only) using a two-sample, two-tailed t-test. The data values given represent the mean value±SEM, and values of p≤0.05 were considered to be statistically significant.
Results
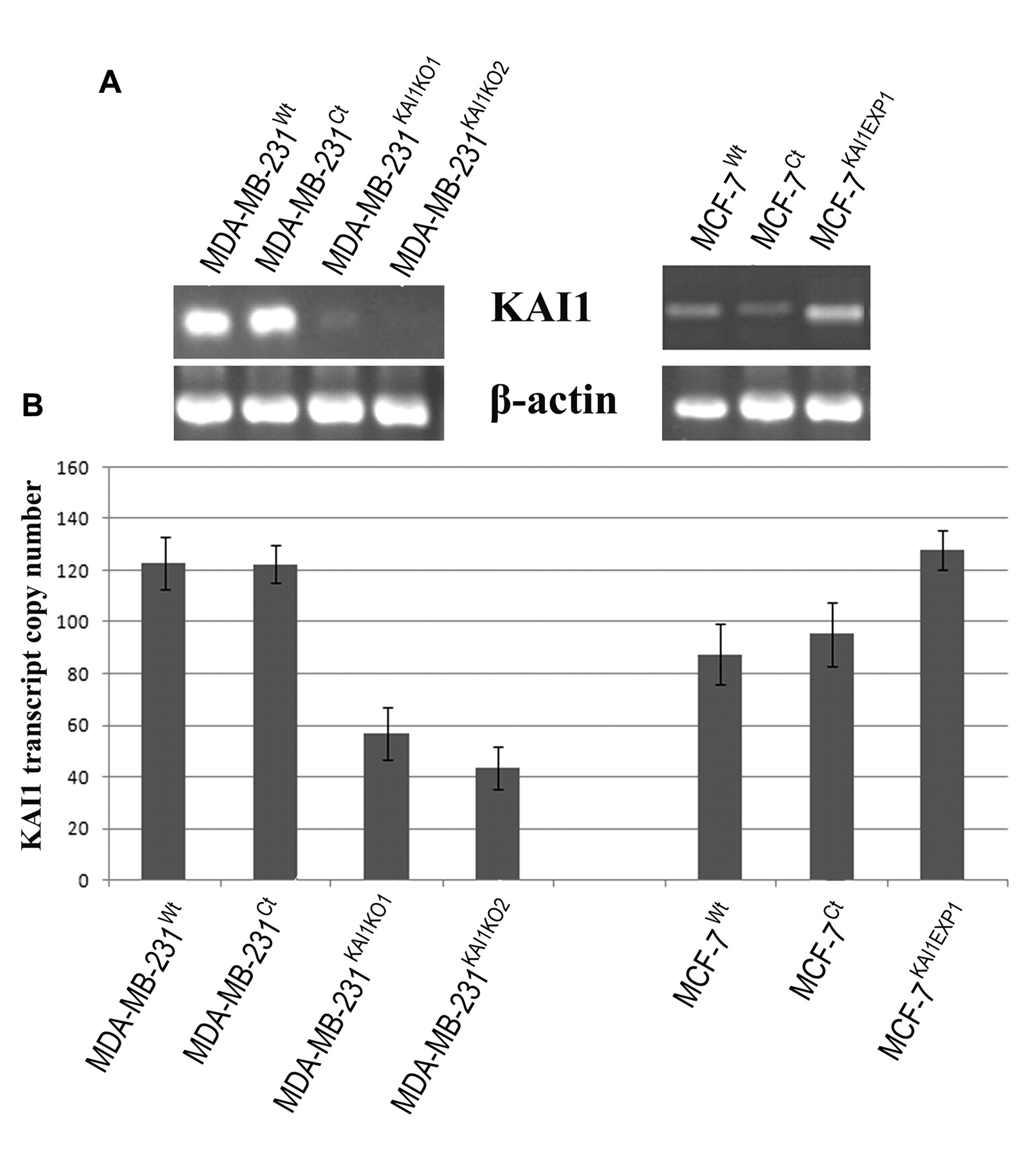
Generation of breast cancer cell lines with altered expression patterns of KAI1. Increased expression of KAI1 transcript levels in MCF-7KAI1EXP1 (cells containing the KAI1 expression plasmid) in comparison to MCF-7Wt and MCF-7Ct (cells containing a closed pEF6 plasmid only) was observed as shown in Figure 1A. Strong reduction of KAI1 transcript levels was observed in both MDA-MB-231 KAI1 ribozyme 1 and 2 compared to wild-type and plasmid control MDA-MB-231 cell lines as shown in Figure 1A. Real-time PCR for KAI1 also confirmed successful forced expression of KAI1 in MCF-7KAI1EXP1 and knockout of KAI1 in MDA-MB-231KAI1KO1/2 at the transcript level. Real-time PCR results were normalised against GAPDH expression and are shown in Figure 1B. The efficiency of the ribozymes and forced expression construct to alter KAI1 expression was similarly assessed at the protein level using both immunocytochemisty (ICC) and Western blot analysis. ICC results show an increased brown staining indicative of KAI1 expression in the MCF-7KAI1EXP1 cells in comparison to MCF-7Wt and MCF-7Ct cells. Similarly, a reduction in KAI1 protein levels in MDA-MB-231KAI1KO1/2 cells was seen in comparison to MDA-MB-231Wt and MDA-MB-231Ct cells (Figure 2A). KAI1 protein, with size ranging from 46-70 kDa was also observed at marginally increased levels in MCF-7KAI1EXP1 cells while a decrease and complete loss of KAI1 protein was seen in both MDA-MB-231KAI1KO1 and MDA-MB-231KAI1KO2 cells respectively as shown in Figure 2B.
Effect of KAI1 on the growth and adhesion of breast cancer cells. Manipulated expression of KAI1 expression in the MDA-MB-231 and MCF-7 breast cancer cell lines did not seem to have any significant effect on cell growth over 3- and 5-day periods (Figure 3 A and B). No significant differences between any of the cells transfected with the ribozyme transgenes or the expression construct and their respective pEF6 controls were observed (p>0.05) at either time point. Knockout of KAI1 resulted in a decreased ability of MDA-MB-231 cells to adhere to an artificial Matrigel basement membrane (Figure 3 C and D). The number of adherent cells observed for both the MDA-MB-231KAI1KO1 (p=0.002) and the MDA-MB-231KAI1KO2 (p=0.0004) cell lines were found to be significantly decreased compared to the MDA-MB-231Ct cell line. In addition, enhanced expression of KAI1 significantly increased the adhesive nature of the MCF-7 cell line with the MCF-7KAI1EXP1 (p=0.021) displaying a significant increase in adhesion as compared to the MCF-7Ct cell line.
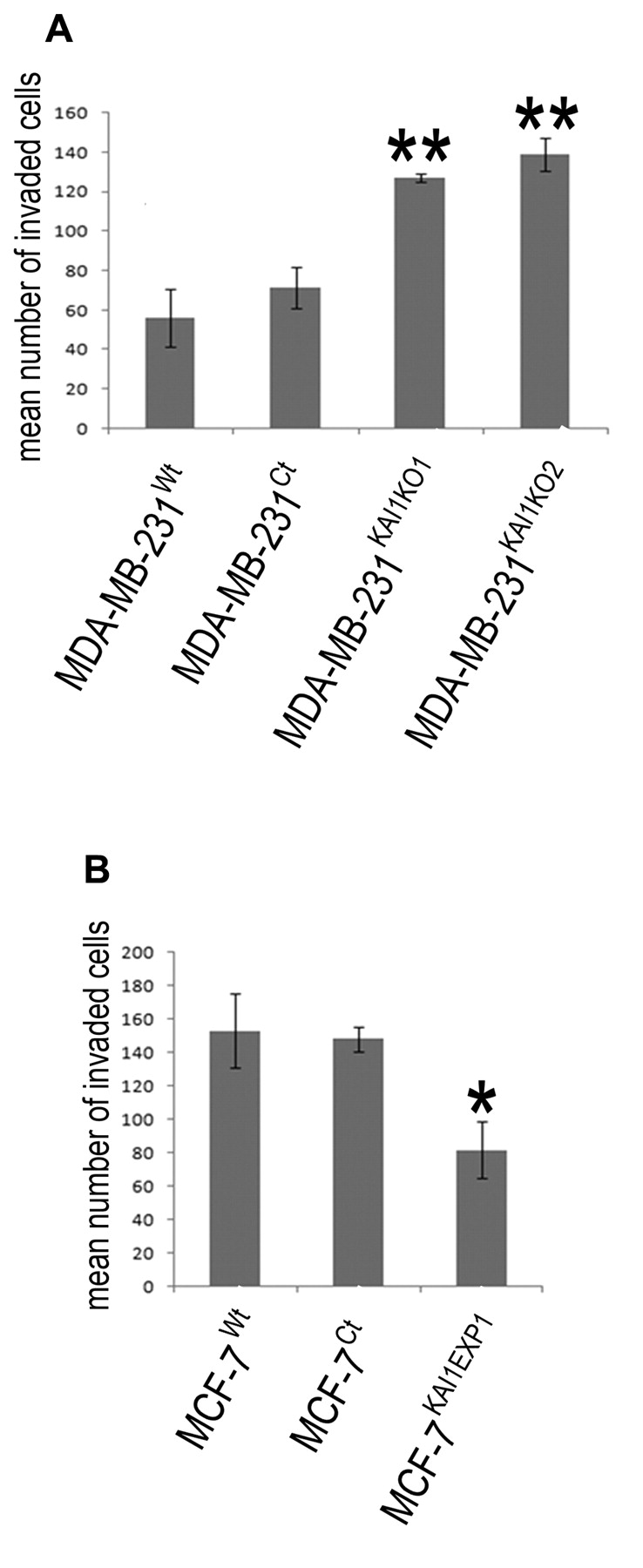
Effect of KAI1 on breast cancer cell motility and invasion. Alteration in the levels of KAI1 was seen to alter the migratory capacity of both breast cancer cell lines (Figure 4). The distance migrated over 90 minutes of both MDA-MB-231KAI1KO1 (p=0.003) and MDA-MB-231KAI1KO2 (p=0.003) cells was significantly less than that of the MDA-MB-231Ct cell line. In keeping with this, enhancing the expression of KAI1 in the MCF-7 cell line, was found to significantly decrease the migratory capacity of the MCF-7KAI1EXP1 cell line over 90 minutes compared to the MCF-7Ct cell line (p=0.024). Alteration of KAI1 levels in these two breast cancer cell lines was also seen to be associated with an alteration in their invasive capacity (Figure 5). In MDA-MB-231 cells transfected with the KAI1 ribozymes, a significant increase in cellular invasion was seen compared to that of the MDA-MB-231Ct cell line (MDA-MB-231KAI1KO1 p=0.0063, MDA-MB-231KAI1KO2, p=0.007). Following a similar trend, when KAI1 levels were raised in the MCF-7KAI1EXP1 cell line, a decrease in invasiveness compared to the control cells was observed (p=0.022). Together these data strongly imply a role of KAI1 in cancer cell invasion.

Confirmation of alteration of KAI1 expression in the MDA-MB-231 and MCF-7 breast cancer cell lines. A. RT-PCR showed (KAI1KO1 and 2) a reduction in the expression of KAI1 in MDA-MB-231 cells containing the KAI1 ribozymes (KAIKO-1 and -2) in comparison to wild-type (wt) and control (Ct) cells. RT-PCR also showed a substantial increase in KAI1 expression in MCF-7 cells following transfection with the expression construct (KAI1EXP1). B. Q-PCR results confirming the RT-PCR data and indicating knockout of KAI1 in MDA-MB-231 cells and the forced expression of KAI1 in the MCF-7 cell line.

Protein confirmation of KAI1 knockout or forced expression. A. ICC staining indicated a reduced level of KAI1 in MDA-MB-231 cells containing the ribozyme transgenes for KAI1 (KAI1KO-1 and -2). Enhanced expression of KAI1 is also visible in MCF-7 cells containing the expression plasmid (KAI1EXP1). This is further confirmed using Western blot analysis (B) where the knockout of KAI1 in the MDA-MB-231 and the forced expression of KAI1 in the MCF-7 cell lines are also apparent.
Discussion
During metastatic spread, cancer cells dislodge from the primary tumour, invade the surrounding extracellular matrix and penetrate into the blood or lymphatic circulation. This systematic spread of cancer cells from the primary tumour to secondary sites is still an area which requires further investigation. KAI1 has been indicated as having a potential role in regulating the metastatic process. Investigation related to prognostic impact of KAI1 in invasive ductal breast carcinoma patients has already been reported as having a significant correlation with TNM staging (13).
KAI1, also termed as CD82, belongs to the tetraspanin family and localises on the cell membrane (22). As reported earlier, alteration in the levels of KAI1 is attributed more toward expressional dysregulation rather than mutations in the coding region (6). KAI1 has a transmembrane localisation with several interactions with integrin (23), chemokines (24), epidermal growth factor receptor (EGFR) (25) and other tetraspanins (26). These interactions are responsible for cell-cell interaction, cell motility, adhesion and cell signalling. Thus, down-regulation of KAI1, both at the transcript as well as translational level, in clinical cases ultimately leads to poor prognosis.
One significant change of cell function after modification of KAI1 expression is that of cell adhesion, as shown in the present study. Increased adhesion of MCF-7KAI1EXP1 with Matrigel has shown a direct correlation of KAI1 with cell adhesion molecules while reduced attachment of cells along the membrane has been observed in MDA-MB-231KAI1KO. KAI1 co-localisation with integrins leads to increase cellular adhesion (27). Integrins are a family of heterodimeric transmembrane proteins which act as receptors for extracellular matrix proteins (28, 29). KAI1 actually works as a linker molecule between integrins and PKC (protein kinase C), induction of phosphorylation of which are involved in cell adhesion and migration (30). KAI1 also has an ectopic effect on adhesion by strengthening the interactions between E-cadherin and β-catenin, in doing so, KAI1 may reduce the chances of cellular dissemination from the primary tumour (31).

In vitro assays assessing the role of KAI1 in growth and adhesion of breast cancer cell lines. A. Knockout of KAI1 did not appear to have any significant effect on the growth of MDA-MB-231KAI1KO1/2 cells over 3- and 5-day periods of incubation. B. Similarly, no significant effects were observed on the growth rates of MCF-7 cells containing the KAI1 expression plasmid (KAI1EXP1). KAI1 did appear to play a role in cell adhesion to a Matrigel membrane. C. Knockout of KAI1 due to either ribozyme transgene significantly reduced the number of adhesive cells when compared to the pEF6 control cells. D. In keeping with this, forced expression of KAI1 in the MCF-7 cell line also significantly increased the adhesive nature of this cell line compared to control cells. **p>0.01, *p>0.05.
In a few earlier studies, it was shown that expression of KAI1 significantly reduced cell adhesion to basement membrane as observed in different cancer cell lines. Our findings, as in colorectal carcinoma cell lines, is consistent with its role in suppressor of metastatic progression (32). These contrasting results may be attributed due to the different natures of the cell lines used as well as the level of integrins at various stages of cell cycle (33). In another study conducted on T-cells co-stimulation, it was reported that binding affinity of intercellular adhesion molecule and leukocyte function associated antigen-1 (LFA-1) is in fact correlated with over-expression of KAI1 (34).

In vitro migration/wounding assay assessing the role of KAI1 in cell motility. A. Reduced KAI1 levels, through the transfection of either ribozyme transgene, was associated with an increase in MDA-MB-231 migration over a 90 minute period (both at p>0.01 compared to pEF6 control). B. Forced expression of KAI1 was associated with a decreased rate of migration in the MCF-7 cell line over 90 minutes (p>0.05 compared to pEF6 control).
The role of KAI1 molecule on cell migration was also assessed by time lapse video recording and showed a direct correlation of KAI1 with cellular motility. MCF-7KAIEXP1 cells showed reduced cell migration towards a wound induced in the near vicinity. In contrast, MDA-MB-231KAI1KO1/2 cells showed increased cellular motility in comparison to MDA-MB-231 wild type and control (empty plasmid) cells. These findings are also in accordance with the feature observed in other MSGs studied in relation to different cancer cell lines used such as nm23 (non-metastatic gene 23), KISS1 (Kisspeptin-1), BRMS1 (breast cancer metastasis suppressor-1) and E-cadherin.
Invasion is the most devastating feature of mammary tumour cell metastatic spread. The cellular invasion process is regulated by the coordinated expressional dysregulation of several factors including mitogen-activated protein kinases (MAPKs). MAPKs ultimately lead to the activation of matrix metalloproteinases (MMPs) and extracellular matrix (ECM)-degrading enzymes. EGFR also influences expression levels of these MMPs and TIMPs via MAPK signal pathway. EGFR binding with its ligand results in increased cellular motility while its over-expression is often correlated with poor survival of the patients. KAI1 interaction with EGFR leads to its internalisation, thereby altering the cells invasive ability (35). Here, we observed that KAI1 forced expression in the MCF-7 cell line reduced cellular invasion. Similarly, an increased cellular invasion across Matrigel membrane was seen in MDA-MB-231 cells containing the KAI1 ribozyme transgenes, showing direct correlation of KAI1 to cell invasion.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
In vitro invasion assay demonstrating the role of KAI1 in cellular invasion. A. Knockout of KAI1 was associated with an enhanced invasive capacity of MDA-MB-231 cells whereas the forced expression of KAI1 (B) was associated with a reduced ability of MCF-7 to invade through a Matrigel membrane. **p>0.01, *p>0.05.
KAI1 does not alter the growth pattern of cancer cells: no significant effect on cellular division or cell proliferation was observed up to 5 days in three independent sets of growth assays. This lack of effect by KAI1 on cell growth was also in accordance with other MSGs including Drg-1, BMRS, RhoGDI2 and NM23. Lack of KAI1 influence on the growth pattern may be attributed to variations in cancer cell type as well as in the microenvironment as reported in the context of another putative MSG (Drg-1) (36, 37). Although Drg-1 did not regulate growth of either prostate or colon cancer cells, it did induce a strong arrest of bladder cell growth both in vivo as well as in vitro assay. Likewise, the HGF/cMET MSG regulating dual has an inhibitory effect on the growth of liver and colorectal cancer cell growth (38, 39), but has not effect on a number of other cancer cell types (40).
Conclusion
Expressional variations of KAI1 molecule in in vitro-based assays strongly indicate the potential importance of KAI1 as a metastatic suppressor in breast cancer cell lines. Thus, regulation of KAI1 is strongly suggested to be a potential target for reducing cancer metastasis.
Acknowledgements
We are extremely thankful to the Higher Education Commission of Pakistan for providing funds for supporting Mr Malik.
Footnotes
- Received April 7, 2009.
- Revision received May 26, 2009.
- Accepted June 24, 2009.
- Copyright© 2009 International Institute of Anticaner Research (Dr. John G. Delinassios), All rights reserved