

Abstract
Background: Despite optimal primary treatment of ovarian cancer, overall prognosis is poor due to recurrences. While steroid hormone receptors are frequently expressed, the role of estrogen receptor (ER) in ovarian carcinogenesis, response to treatment or prognosis has not been established. We analyzed the gene-expression in response to estradiol (E2) and genistein (Gen) in ovarian cancer cells. Materials and Methods: Cell lines (Br-1, UL-1; Oy-1), treated with E2 (10 nM) or Gen (5 μM), were used for gene expression profiling. RT-PCR and Western immunoblotting were used to further analyze gene expression data. Results: Twenty-four genes were differentially regulated in ovarian cancer cell lines. C3, CLU, COL6A1, DLC1, NME1, NRIP1, PTEN, RAC2, S100A2 were down-regulated with E2 in Br-1 and UL-1 cells. MK167, SERPINB5, SLC7A5, CDK1NA, LCN2, PLAU, PHB2, CTSB, EGLN2, ERBB2, HMGB1, ID2, ITGB4, TOP2A were up-regulated in Oy-1 cells with E2 and/or genistein. ERBB2 and ID2 (E2 and Gen), LCN2, PHB2 and HMGB1 (Gen) were down-regulated in Br-1 cells. ERα and ERβ were detected in all cell lines at different levels. Conclusion: Variable response of ovarian cancer cells to E2 and Gen was observed. Study of ERs including splice variants, co-regulatory molecules are necessary to understand the relevance of receptors.
- Ovarian cancer
- gene expression
- estrogen receptor
- genistein
- estradiol
Ovarian carcinoma (OC) is the most lethal of all gynecological malignancies. Annually, it results in over 16,000 deaths in the US. Because the symptoms of ovarian cancer are non-specific, the majority of patients do not present until their disease has metastasized (1, 2). Involvement of steroid hormones, mainly estrogen and progesterone, has been associated with ovarian cancer. Estrogens are major regulators of growth and differentiation in normal ovaries and potentially play a role in the development and progression of ovarian carcinoma, but the mechanisms of action remain unclear (3). The actions of estrogens on ovarian tissue are mediated by two estrogen receptor (ER) variants, ERα and ERβ (4). While ERβ is predominantly expressed in the normal ovary, ERα is expressed to a much greater extent in malignant ovarian tumors (5). Estradiol directly regulates the transcription of a number of genes involved in many aspects of cellular function such as DNA repair, extracellular matrix, apoptosis and signal transduction (6). The prognostic significance of tumor ER and progesterone receptor (PR) status in OC remains controversial. The role of ER regulation in ovarian carcinogenesis requires further investigation including selective contribution of ERα and ERβ.
First line treatment for ovarian cancer consists of surgical cytoreduction followed by chemotherapy (7, 8). Endocrine therapies have proven effective in other hormonally sensitive cancers with relatively little toxicity. Experimental studies using preclinical models have demonstrated that ovarian cancers that express high levels of ERα are growth responsive to both estrogens and antiestrogens (9). This raises the possibility of therapeutic potential in targeting the ERα in this disease. The anti-estrogen tamoxifen has been used to treat chemo-resistant ovarian cancer in addition to aromatase inhibitor Letrozole (9). Forty to 60% of ovarian cancers express ERα, but only a small proportion of patients (7-18%) respond clinically to antiestrogen treatment (10). ER negative and PR positive ovarian cancers, which accounted for approximately 10% of all tumors, showed significantly superior prognosis when compared with all other combinations of ER and PR expression statuses. However, in another large study, elevated expression of ER and PR, alone and in combination, point to a favorable outcome of patients with OC (11). While ER overexpression might imply a poorer prognosis of ovarian cancer, no significant differences between the survival curves of ER positive and ER negative patients were observed (12).
In addition to estrogen, ovarian tissue is subjected to molecules with estrogenic properties such as genistein which is the main component of soy products and is consumed in supplements (13, 14). Genistein possesses weak estrogenic properties and has weak affinity for the estrogen receptor. Genistein's structural similarity to endogenous estrogen, its ability to bind to both ERα and ERβ, with preferential binding to ERβ has been well documented. It has also become apparent that genistein can regulate the growth of ER negative cell lines indicating that it can function independently of the ER. Genistein's effect on the regulation of gene expression in ovarian cancer has not been well characterized.
The purpose of this study was to determine gene expression, that is specifically involved with the estrogen receptors in ovarian cancer cells in response to estradiol and genistein.
Materials and Methods
Culture and treatment of ovarian cancer cells. Ovarian cancer cell lines (Br-1, Oy-1 and Ul-1) were established in our laboratory from tumor samples obtained from University of Louisville Hospital under an approved IRB protocol. All these cell lines had been in culture for approximately 60 passages at the time these experiments were performed. Cells were cultured in RPMI medium supplemented with 10% charcoal-treated fetal bovine serum. Cells were treated with E2 (0.1 nM) or genistein (5 μM) for 48 hours.
Oligoarray analysis. Gene expression profiling was performed by using pathway-focused oligoGE arrays containing 113 genes (SABiosciences, Frederick, MD, USA). Total RNA was isolated from cultures with the designated treatments using Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's protocol. Following cRNA amplification and labeling with biotin 16-UTP, hybridization was performed. Array imaging and data analysis were performed by GE array expression analysis suite 2.0. Gene expression following correction for background and housekeeping genes is presented as fold over untreated controls.
Real-time PCR (RT-PCR). RT-PCR studies were carried out to validate the data derived from microarray analysis. Two micrograms of total RNA was used to generate cDNA by reverse transcription. RT-PCR was performed in triplicate reactions using X-SYBR Green Master Mix in LightCycler II (Roche) with the appropriate primers for CDKN1A, ERBB2 and TOP2A (SABiosciences). The template cDNA was diluted from the reverse-transcribed product by 4-fold for use in the PCR. Fold changes compared to controls were calculated and statistical significance was determined by a paired Student t-test with p<0.05 regarded as significant. The housekeeping gene GAPDH was used as normalizer gene, because it displayed the lowest variation in expression levels as judged by microarray analysis and the smallest standard deviation in threshold cycle (Ct) values when compared with other known normalizer genes.
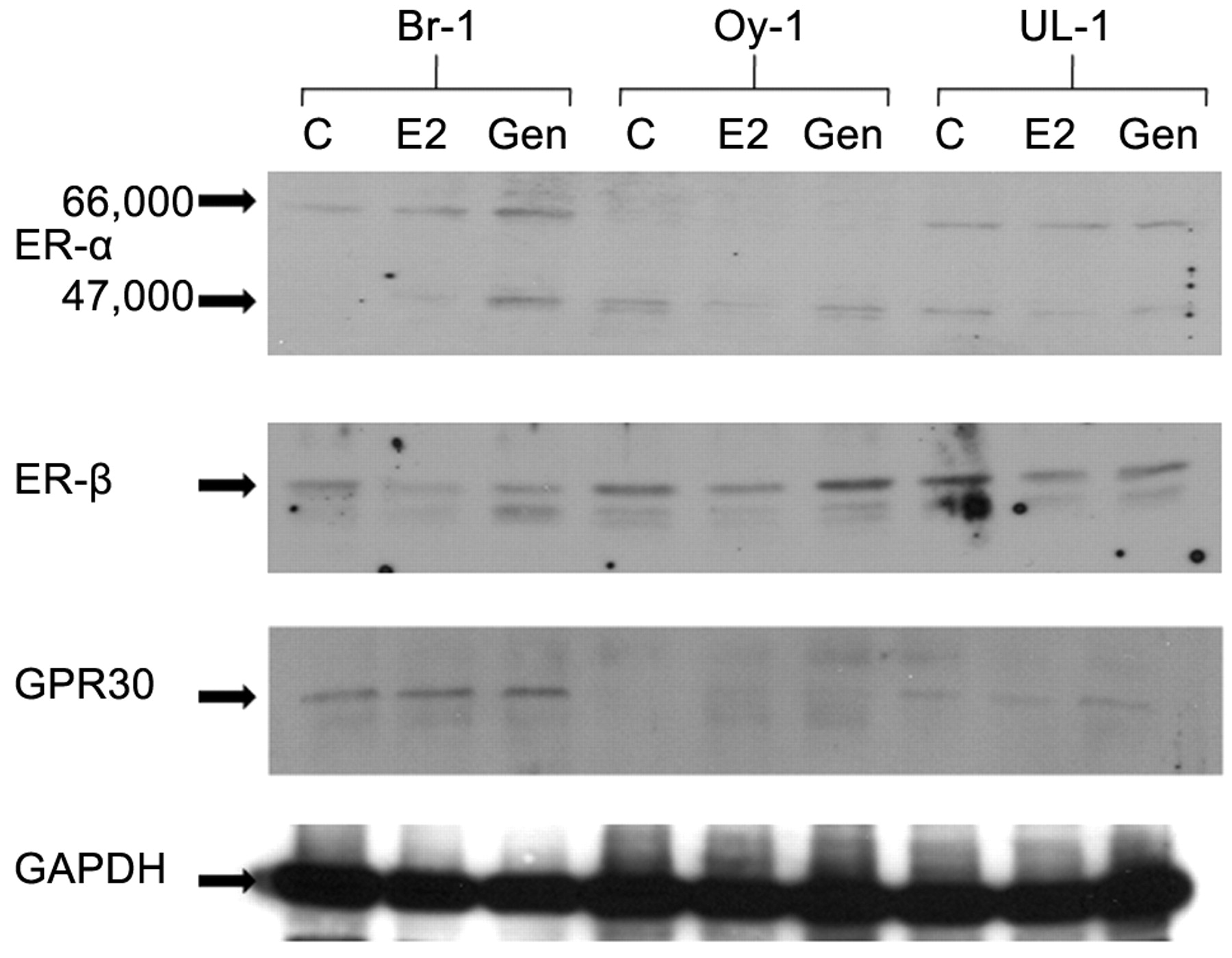
Western immunoblotting. Cells (treated, control and vehicle control) in log phase were washed and lysed in 2% SDS, 10% glycerol, and 50 mM Tris (pH 6.8). Protein concentrations were determined using Bio-Rad assay. Equal amounts of protein (30 μg) were separated on 8-16% gradient polyacrylamide gels. Transfer to nitrocellulose was performed in Bio-Rad transblot apparatus. The blots were blocked with 5% non-fat dry milk and antibodies were used at 1 μg/ml. Primary antibodies to ER-α, ER-β (both mouse monoclonals) and GPR30 (goat polyclonal) were utilized (Santa Cruz Biotechnology). Appropriate secondary antibody (peroxidase-conjugated anti-mouse or goat IgG) was incubated for 45 minutes. Enhanced chemiluminescence (ECL) was used to visualize the specific binding of the antibodies. These experiments were repeated three times and a representative blot is being presented.
Genes displaying a 2.0 fold or greater reduction in expression compared to untreated controls following treatment with 10 nM E2 and 5 μM genistein.
Genes displaying a 2.0 fold or greater increase in expression compared to untreated controls following treatment with 10 nM E2 and 5 μM genistein.
Genes that were differentially regulated by ovarian cancer cell lines (2.0 fold or greater change compared to controls) following treatment with 10 nM E2 and 5 μM genistein.

RT-PCR confirmation of selected genes that were differentially regulated in ovarian cancer cells. Br-1, Oy-1 and UL-1 cells were treated with E2 (10 nM) or genistein (5 μM) for 48 hours. A: CDKN1A; B: ERBB-2; C: TOP2A.
Results
Gene expression analysis. Microarray analysis was performed on focused microarrays to study the response of ovarian cancer cells and selected genes involved in estrogen receptor signaling. Twenty-four of the genes on the microarray were differentially regulated by the ovarian cancer cell lines (Tables I, II and III). Genes that were down-regulated by either or both Br-1 and UL-1 are shown in Table I. C3 was down-regulated by both E2 and Gen treatments in Br-1 and UL-1 cells. CLU, COL6A1, DLC1, NME1, NRIP1, PTEN, RAC2 and S100A2 genes were down-regulated with E2 in UL-1 cells. While COL6A1 were down-regulated by genistein in both Br-1 and UL-1 cells, only DLC1 in UL-1 cells and NRIP1 in Br-1 cells were affected in a similar fashion. No significant change in gene expression was seen in Oy-1 cells with these nine genes. Three genes, MK167, SERPINB5 and SLC7A5 were up-regulated by genistein or E2 treatment of Oy-1 and not Br-1 or UL-1 cells (Table II). Twelve genes were differentially regulated by the three cell lines (Table III). Up-regulation of CDK1NA, LCN2, PLAU and PHB2 was observed with Oy-1 cells in response to E2 and genistein, and CTSB, EGLN2, ERBB2, HMGB1, ID2, ITGB4 and TOP2A with genistein only treatments. All of these genes except ID2 were down-regulated with E2 treatment of UL-1 cells, while CDK1NA, EGLN2, ERBB2, FOSL1 and ITBG4 were down-regulated with genistein only. FOSL1 was the only gene up-regulated with Br-1 cells in response to E2 treatment. ERBB2 and ID2 were down-regulated by both E2 and genistein in Br-1 cells. LCN2, PHB2 and HMGB1 were inhibited by genistein only.
RT-PCR confirmation of microarray data. Transcription of the three differentially regulated genes was confirmed by RT-PCR (Figure 1). The trends in significant up-regulation or down-regulation were similar to that obtained from microarray data.
Western immunoblot analysis of ER receptors. Br-1 and UL-1 expressed similar levels of ERα, while expression of this receptor was barely detectable in Oy-1 cells (Figure 2). Increased expression of high MW ERα was demonstrated in Br-1 with E2 and Gen treatments. Low molecular weight ERα receptor was detected in Oy-1 cells with slight decrease in expression with E2, and no difference with genistein treatment. Low molecular weight ERα was increased in Br-1 cells and decreased in UL-1 cells with E2 and genistein treatments.

{kind=link}
{kind=link}
Western immunoblot analysis of the expression of ER-α, ER-β and GPR30 in Br-1, Oy-1 and UL-1 cells. Cells were treated with E2 (10 nM) or genistein (5 μM) for 48 hours. C: untreated control; E2: estradiol; Gen: genistein.
With respect to ERβ, all cell lines expressed this receptor with Oy-1 and UL-1 slightly higher levels than Br-1. ERβ levels were increased with genistein treatment of Oy-1 cells, and decreased in Br-1 and UL-1 cells.
GPR30 levels were highest in Br-1 cells, less in UL-1 cells and barely detectable levels in Oy-1 cells. Treatment with E2 and genistein did not result in significant changes in Br-1 and UL-1 cells, however slight induction was seen in Oy-1 cells with both treatments.
Discussion
Gene expression analysis was performed to study the effects of E2 and genistein on ovarian cancer cells at physiologically achievable levels. This study included multiple cell lines isolated from Stage IIIC ovarian cancers, while previous studies were performed with one cell line exposed to multiple treatments (9). Our data demonstrate that 24/113 genes were differentially regulated in the three ovarian cancer cell lines. The effect on gene expression suggests that some cells respond to E2 in a manner that induces gene expression (Oy-1) and others are inhibited, such as seen with UL-1 and Br-1, even though some selectivity is observed. At least in this study, ERα (66 kDa) expression is inversely related to induction of the gene expression, however, the role of ERα-46 needs further study. It is of interest that among down-regulated genes are those with tumor suppressive ability such as PTEN, NME1 and DLC1.
The actions of estrogen and estrogenic compounds have been traditionally described to occur through one of the two classical nuclear ERs. They function as ligand-dependent transcription factors and bind directly to estrogen response elements in the promoter regions of genes and recruit additional transcriptional co-regulators through ER activation domain interactions (15). Both ERα and ERβ mediate the transcriptional activity of estrogen by influencing the regulatory molecules co-activators and co-repressors that in turn activate or repress the transcriptional activity of genes responsive to estrogen (16). There are currently 49 known ER-interacting coactivators and 18 known ER-interacting corepressors. Their potential role in our findings as well as others needs to be further investigated. Both ERα and ERβ genes undergo alternate splicing, giving rise to variant proteins such as ERα-46 for ERα which was expressed in all three ovarian cancer cell lines in our study. The role of the diverse splice variants in the regulation of E2 induced cell proliferation or cancer progression is not known (17). In breast cancer cells, in addition to a predominant full-length 66 kDa form, the ERα gene also produces a 46 kDa isoform, lacking N-terminal domains. ERα-46 has been shown to suppress not only the activation function -1 (AF1) activity of the full length ERα but also the transcription of target genes. However, whether the presence of ERα-46 has any association with anti-tumor treatment remains unknown (3). Additionally, E2 activated non-genomic pathways activated by ER ligands, such as the mitogen-activated protein kinase (MAPK)/epidermal growth factor receptor (EGFR) pathway, has been recently described (17,18). Estrogen mediated rapid signaling events results in the activation of cellular kinases, synthetases and ion channels. A structurally unrelated ER GPR30 located predominantly at the cell surface is involved in estrogen signaling (19,20). Genistein has been shown to exhibit high affinity and selectivity for GPR30. Although the genomic and nongenomic actions of estrogen have been studied separately, these actions are interdependent and should be considered synergistic in the molecular response to E2 (17). GPR30 was expressed at different levels by the three ovarian cancer cell lines, the least being demonstrated in Oy-1 cells.
Our data demonstrate the expression of specific genes in response to E2 and genistein. Among these, clusterin was identified as one of the genes that was significantly up-regulated in short term survivors (21). In addition, high clusterin expressing cells were associated with resistance to paclitaxel. Lipocalin 2, up-regulated in Oy-1 cells, and S100A2, down-regulated in UL-1 cells, are among genes that are up-regulated in ovarian serous papillary carcinomas (22). Topoisomerase II and HER-2 gene amplification and expression have been demonstrated in ovarian cancer (23).
DLC-1 and DLC-2 are emerging tumor suppressor genes. Lower expression of DLC-1 and DLC-2 is seen in high percentage of tumors, suggesting that deficiency of either DLC gene facilitates dissemination of carcinoma cells to secondary sites (24). NME1 (nm23-H1) is a tumor suppressor gene and is expressed in normal cells. Change in nm23-H1 expression and the PIK3/AKT pathway are involved in carcinogenesis, development, invasion and metastasis of ovarian cancers. Expression of nm23-H1 was negatively correlated with tumor stage and grade. Estrogen reduced nm23-H1, which ultimately resulted in increased cell migration and invasion (25). Similarly PTEN is a tumor suppressor gene and its reduced expression is a poor prognostic factor for disease free survival in patients with epithelial ovarian cancer (26).
Previous studies with an ER-positive ovarian cancer cell line demonstrated that PLAU, VIM, BIGH3, CDH6, FN1, CASP4, KRT4, KRT7, KRT13, TRAM and NGAL were down-regulated and TFF1, TFF3, TRAP1, TFAp4, MYC, CTSD, IL17BR, TOP2A, CCNB1, CCNB2, PDZK1 and UBE2C were up-regulated by E2. The modulation of these genes was reversed by the anti-estrogen tamoxifen and was ERα dependent. Similarly, genistein treatment at 5 μM of an endometrial cell line induced multiple changes in gene expression including some implicated in oncogenesis (13, 14).
Many ovarian cancers express high levels of ERα. Lee et al reported 86% of ovarian tumor specimens stained positive for ER, 50% positive for PR, and 45% positive for both receptors (26). Among the various ovarian cancer subtypes, ERα immunostaining was found in 97% of serous adenocarcinomas, 100% of endometrioid adenocarcinomas, 70% of mucinous adenocarcinomas and none of the clear cell carcinoma specimens. In contrast, ERβ immunopositivity was found in all ovarian cancer subtypes (38% of clear cell carcinoma, 41% of serous adenocarcinoma, 30% mucinous adenocarcinomas and 75% of endometrioid adenocarcinoma). One may speculate that ER overexpression might imply a poorer prognosis of ovarian cancer; however, this was not observed in a recent study and no significant differences were seen between the survival curves of ER positive and ER negative patients (12). Another study demonstrated that an elevated expression of ER and PR, alone and in combination, point to a favorable outcome for patients with ovarian cancer. This is the largest study describing the relationships between ER and PR expression in ovarian tissue and also with respect to examination of the prognostic values of the receptor status in ovarian cancer (11). In epithelial ovarian carcinoma, the prognostic significance of tumor ER and PR status remains controversial (10). Use of antiestrogen Letrozole in ovarian cancer has consistently demonstrated a significant level of activity and the larger clinical studies have linked response with ERα expression and other biomarkers. Subgroups of ovarian cancer are sensitive to hormones and will respond to endocrine therapy. To optimize these approaches further, it will be important to define characteristics within tumors that confer sensitivity (27).
Based on our findings, more in-depth understanding of the action of estrogens on normal and malignant ovarian tissue is warranted to understand the role played in carcinogenesis, as well as their therapeutic potential.
Footnotes
- Received March 11, 2009.
- Accepted March 30, 2009.
- Copyright© 2009 International Institute of Anticaner Research (Dr. John G. Delinassios), All rights reserved