

Abstract
The current study investigated the expression of claudins 1, 5 and 16 in human clear cell renal cell carcinoma (CCRCC) and the impact of claudin-16 on kidney cancer cells. Levels of claudin transcripts were assessed using quantitative reverse transcription-polymerase reaction and proteins were examined by immunohistochemical methods. Human kidney epithelial cell HEK293 and human kidney cancer cell line UMRC2 were also used in the study. Human kidney tissues expressed high levels of claudin-1 and in particular claudin-16, but were negative for claudin-5. Late-stage kidney carcinomas tended to express low levels of claudin-1 and claudin-16. UMRC2 cells, highly positive for claudin-16, were transfected with an anti-claudin-16 transgene, which resulted in significant reduction in both transepithelial resistance and cell matrix adhesion compared to control cells. When HEK293 cells, negative for expression of claudin-16, were forced to express claudin-16, the cells showed a marked increase in transepithelial resistance and to some degree in cell matrix adhesion. In conclusion, the present study showed that kidney tissue and CCRCC tissues highly expressed claudin-16 and claudin-1, and that claudin-16 has a profound impact on barrier function and cell adhesion in kidney cancer cells.
- Claudin-1
- claudin-5
- claudin-16
- clear cell renal cell carcinoma
- paracellular permeability
Tight junctions are the most apical cellular structure in epithelial and endothelial cells. Tight junctions create a regulated paracellular barrier to the movement of ions, solutes, macromolecules and cells, such as immune cells, between epithelial and endothelial cells, and to a certain degree act as signaling pathways that communicate cell position, and limit growth and apoptosis. Thus, this cellular structure has two exclusive functions: firstly by acting as a tissue fence by forming an apical/basolateral intramembranous diffusion barrier which prevents the mixing of membrane proteins; and secondly by acting as a gate by controlling the breadth and selectivity of diffusion along the paracellular pathway (1-3). The tight junctions are formed between two neighbouring cells and at the cellular level by three groups of proteins, namely the membrane integral proteins (including occludins, claudins, and Junctional Adhesion Molecules (JAMs), membrane associated/subcoat proteins such as zonula occludens (ZOs), and tight junction-associated proteins.
Out of these proteins, the membrane integral proteins are perhaps most intriguing as they offer direct protein-protein interactions between two cells. Claudins, or the claudin protein family members, are the most interesting of these (4, 5). First discovered by Furuse et al. in chicken liver (6), subsequent research has identified that the human claudin protein family has 24 members. These proteins, 22-27 kDa in size, have four transmembrane domains where the N- and C-terminal domains are orientated towards the cytoplasm. Claudins from different cells form homotypic and heterotypic interactions which form a key part of paracellular barriers.
Tight junctions play very important roles in maintaining distinct tissue space and in controlling paracellular permeability. This is particularly so for endothelial cells (for the blood-tissue space), intestinal epithelial cells (for gut permeability) and kidney (for the blood–tissue–water system) (7-9).
The role of tight junctions in cancer development and progression has been explored in recent years, although a firm causal relationship is yet to be established. For example, it has been shown that integral membrane proteins occludin and certain claudins are reduced in epithelial-derived malignant conditions and are linked to disease progression and clinical outcome of the patients. Claudin-20 has been found to be increased in aggressive breast tumors and, being associated with poor patient survival and metastatic disease, demonstrates its possible use as a target for therapy and as a marker for determining treatment regimens (10). Moreover, a review by Webb et al. summarized that new functions for claudins outside tight junctions were supported by the discovery that the disruption and reduction/loss of tight junctions structure occurred during tumour progression (11). Moreover, there is a distinct expression of claudin subtypes (12). Unlike claudin-5 and claudin-16 which are significantly reduced in human breast cancer, a number of claudins are up-regulated in cancer (13, 14). Claudin-4 is up-regulated in breast cancer as are claudin-3 and -7 (15,16). Claudin-4 overexpression has also been shown to promote gastric cancer metastasis through increased invasion of gastric cancer cells (17). Claudin-1 overexpression is associated with advanced clinical stage and invasive pathological characteristics of oral squamous cell carcinomas (18). Interestingly, a recent study showed that in renal cell cancer, positive expression was detected in 62%, 67%, 45%, 55%, 7% and 35% of cases for claudins 1, 2, 3, 4, 5 and 7, respectively. High expression of claudin-2 was observed in 20% of cases, while high expression of other claudins was less frequent. Claudins were compared to classical prognostic factors. Claudin-1 and claudin-2 were significantly associated with lower-grade and higher-grade tumors, respectively. None of the claudins was significantly associated with tumor stage or patient survival (note claudin-16 was not investigated in the study) (19).
Claudins 1, 3, 4, 7, and 8 have been reported in human kidney tissues, with claudins 3,4,7, and 8 found to be both membranous and cytoplasmic, whereas claudin-1 is predominantly membranous (20), and may be useful markers in distinguishing renal cell carcinomas from oncocytomas (21). Fritzsche et al. showed claudin-1 to be highly expressed in papillary renal cell carcinoma compared with clear cell carcinoma (76-86% vs. 29.9% respectively) (22). The authors reported that expression of claudin-1 was associated with unfavourable outcome of the patients. Claudin-7 and claudin-8 are seen in the distal nephron epithelium. In renal cell carcinoma, claudin-7 and claudin-8 were found to be reduced in chromophone renal cell carcinoma (23).
Of particular interest, in terms of expression of claudins in the kidney is claudin-16, also known as paracellin-1, which was identified in 2000 from an inherited disease known as hypercalcaeuria and hypomagnesaemia symdrome (24, 25). The study identified that the disease was caused by the mutation of then a new protein, subsequently named as paracellin-1 and claudin-16, which results in the loss of control of the paracellular re-absorption of both calcium and magneseum. As a consequence, patients suffer uncontrolled loss of calcium and magneseum through the urine. Clinically, this is manifested as the development of kidney stones at an early age and low serum levels of calcium and magnesium. Despite the importance of claudin-16, the expression of this molecule in kidney cancer has not been studied as far as we are aware.
In the present study, we investigated the pattern of expression of claudin 1, 5, and 16 in clear cell renal cell carcinoma (CCRCC) and also explored the role of claudin-16 in the biological behaviour of renal cancer cells.
Materials and Methods
Materials. Antibodies against claudin-1(SC-17658), claudin-5 (SC-28670) and claudin-16 (SC-10178) were purchased from Santa Cruz Biotechnologies Inc. (Santa Cruz, CA, USA). A universal ABC kit for immunohistochemical staining was used from Vector Laboratories Inc. (Burlington, CA, USA).
Cells and tissues. The human kidney cancer cell line, UMRC2 and immortalized kidney epithelial cell HEK293 were purchase from the Americal Type cell Culture (ATCC)/LGC standard (Teddington, Middlesex, UK). Cells were routinely maintained in DMEM-F12 supplemented with 10% foetal cal serum and antibiotics. Human CCRCC tissues (n=23) were obtained from the Department of Urology, Beijing ChaoYang Hospital of Capital Medical University, PRC. The tissues were immediately collected after surgery and stored in liquid nitrogen until use. Ethics approval was obtained from the Capital Medical University Chaoyang Hospital Research Ethics Committee and consent obtained from the patients.
Extraction of RNA and preparation of cDNA from cells and tissues, qualitative and quantitative analysis of claudin transcript. Fresh frozen tissues were first homogenised in a Trizol RNA extraction buffer (Sigma-Aldrich, Poole, Dorset, UK) using a handheld homogenizer. Cell pellets were also lysed in the same buffer. Total RNA was extracted and purified by following the manufacturer’s instructions. cDNA was synthesised from equal amounts of RNA by using a first-strand DNA systhesis kit (BioRad, Hemel Hemstead, England, UK). Routine polymerase chase reaction (PCR) was carried out to detect the present of claudin transcript by using the respective primers for claudins-1, 5 and 16 (Table I). Quantitative analysis of claudin transcript was carried out using Amplifor™-based technologies, in which a 6-carboxy-fluorescine-tagged Uniprimer™ (Biosearch Technologies, Inc., Novato, CA, USA) was used as a probe together with a pair of target specific primers, to one of which was added a Z-sequence (actgaacctgaccgtaca). An internal standard was used as control and Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as the housekeeping control for both PCR and quantitative PCR. The quantification was carried out using Stepone-plus quantitative PCR unit (ABI, Paisley, Scotland, UK).
Immunohistochemical analysis of claudins. Paraffin-embedded CCRCC was sectioned, dewaxed and prepared for immunohisto-chemical analysis. Briefly, slides were blocked using horse serum buffer before the primary antibody (made in 1% Tris balanced solution/bovine serum albumin) was applied and the sections incubated for 30 minutes. Antibody localisation was then identified by a standard stretavidin-biotin peroxidase technique using Vector Elite ABC Kit by incubating the sections with a relevant biotinylated secondary antibody for 30 minutes, followed by incubation with the avidin-biotin complex (ABC) reagent provided in the kit for a further 30 min. The final reaction product was developed for 10 min with 3,3’-diaminobenzidine substrate (0.005%) serum (Dako Ltd.®, High Wycombe, UK). The sections were then rinsed in TBS, followed by tap water, and then counterstained with Ehrlich’s haematoxylin solution (BDH-Merck, Poole, UK) for 30 sec, and then washed again in tap water for 5 minutes. Finally, sections were dehydrated through a graded series of alcohol solutions (BDH-Merck) for 5 min in each and mounted in DPX medium (BDH-Merck) before mounting under a coverslip. The staining pattern and staining intensity of the claudins were observed under the microscope.
Primers used for polymerase chain reaction (PCR), quantitative PCR and amplification of claudin coding sequence.
Creation of kidney cancer cell lines with differential expression of claudin-16 and the impact of claudin-16 expression on cellular behaviour. The construction of a human claudin-16 expression vector and anti-claudin-16 ribozyme transgenes were carried out as we previously reported (26). Kidney cancer cell line UMRC2, which was positive for claudin-16 expression, was transfected with anti-claudin-16 transgene (26) and HEK293 cells, which were negative for claudin-16, were transfected with claudin-16 expression vector (26) by way of electroporation. The expression of claudin-16 was examined 24 hours after transfection and the cells were used within 96 hours, owing to the fact that the kidney cancer cells used were slow growing and survived the selection process with difficulty. Transepithelial electric resistance was assessed using the EVOM system volt-ohmmeter (World Precision Instruments, Aston, Herts, UK), equipped with a pair of STX-2 chopstick electrodes (27,28). Briefly, HEK293 or UMRC2 cells were seeded into transwell inserts with 0.4 μm pore size (Greiner Bio-One Ltd., Stroudwater Business Park, Stonehouse, England, UK). The cells were then allowed to reach full confluence, after which fresh medium was replaced for further experiments. Electrodes were placed at the upper and lower chambers and resistance measured with the volt-ohmmeter.
Electric cell-substrate impedance sensing (ECIS)-based cellular adhesion assays. An ECIS-Zθ instrument with a 96-well station and wounding module (Applied Biophysics Inc., Troy, NJ, USA) was used for the cell adhesion assays (29). Cell modelling was carried out using the ECIS RbA modelling software, supplied by the manufacturer (30). The 96W1E ECIS arrays (Applied Biophysics Inc.) were used, which had 96-wells in each array allowing multiplex assays. ECIS measures the interaction between cells and the substrate to which they are attached via gold-film electrodes placed on the surface of culture dishes. The adhesion was tracked immediately after placing the cells into the arrays. Impedance and resistance of the cell layer were immediately recorded for a period of up to 30 h over a range frequencies (1,000 to 64,000 Hz). Data were analysed using the software supplied by the manufacturer.
Statistical analysis. This was carried out using Sigma Plot (version 11; Systat Software Inc., San Jose, CA, USA). Student t-test was to calculate the significance for normally distributed data.
Results
CCRCC differentially expressed claudins. We quantitatively analyzed the levels of claudin transcripts in the CCRCC tissues and the paired normal tissues. As shown in Table II, kidney tissues generally expressed higher levels of claudin-16 than of claudin-1 and claudin-5, with levels of claudin-5 being the lowest. Both claudin-1 and claudin-16 transcript levels were observed to be higher in tumor compared to normal tissues, although the difference was not statistically significant. Grade 2 and grade 3 tumors had higher levels of claudin-1 and claudin-5 transcript than did grade 1 tumours. The levels when grade 2 and grade 3 tumors were combined wee significantly higher than those of grade 1 tumors (p=0.0135 for claudin-1 and p=0.012 for claudin-5). Claudin-16, although demonstrating high levels in grade 2 tumors, did not have a statistical link with tumour grade, nor with TNM staging.
Immunohistochemical staining. As shown in Figure 1, normal kidney tissues stained positively for claudin-1 (left panel) and claudin-16 (right panel), but very weakly for claudin-5 (middle panel). In CCRCC tissues, staining for both claudin-1 and claudin-16 was similar to that of normal tissues. Staining for claudin-5 remained negative.
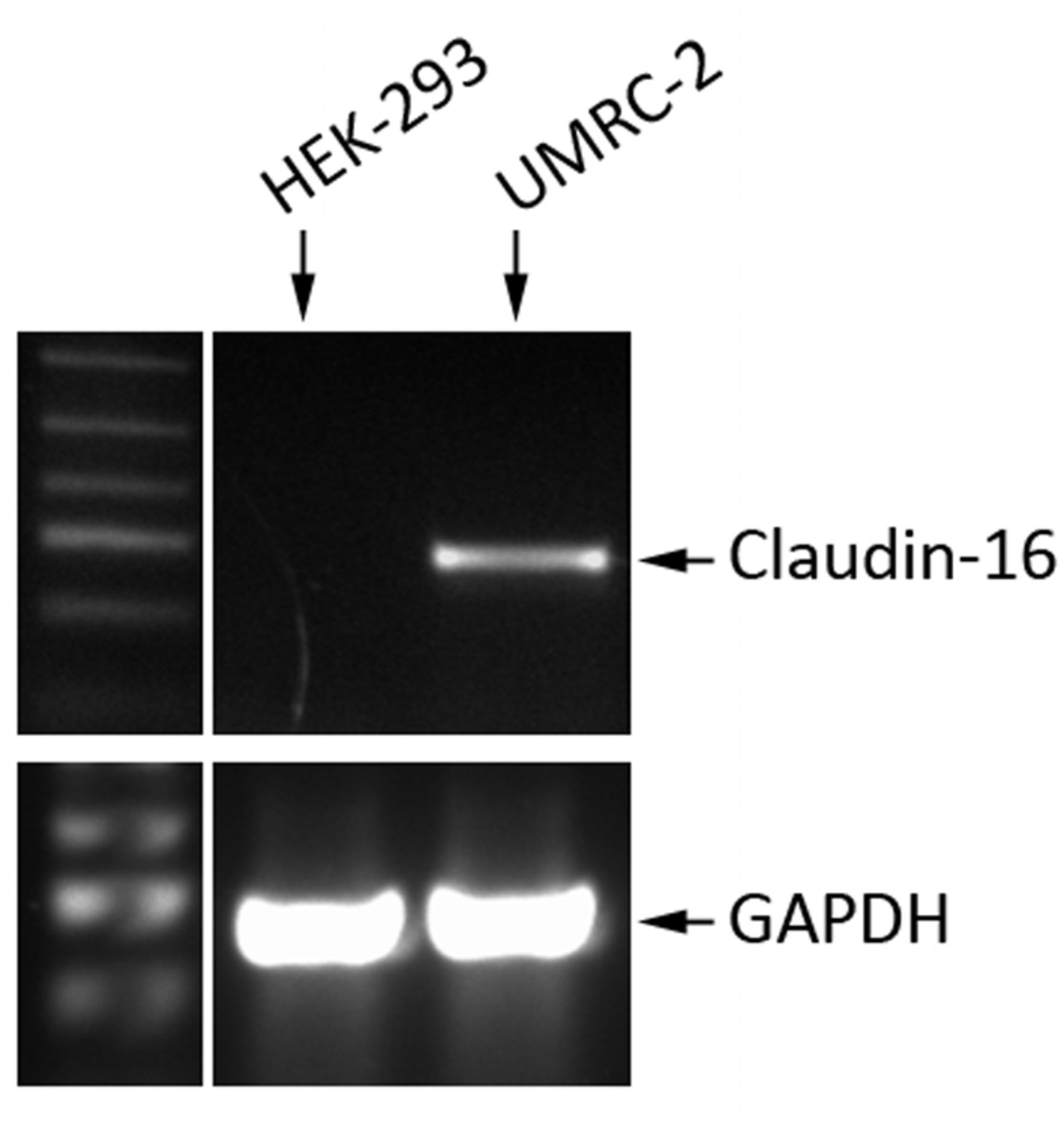
Genetically manipulating expression of claudin-16 had an impact on the paracellular permeability of kidney epithelial and RCC cells. Owing to its high level in kidney tissues, we examined the expression of claudin-16 in kidney cell lines, HEK293 and UMRC2, which are an immortalised normal foetal kidney epithelial cell line and a renal cell carcinoma cell line, respectively. As shown in Figure 2, UMRC2 cells had high levels of claudin-16 transcripts, whereas HEK293 was largely negative for claudin-16 expression.

Immunohistochemical staining of claudin-1 (A), claudin-5 (B) and claudin-16 (C) of clear cell renal cell carcinoma tissues (left) and residual normal kidney tissues (right in each panel). Claudin-1 and claudin-16 are positively stained in both tumor and normal tissues. Claudin-5 staining is negative. Top panel: ×100 magnification; Bottom panel: ×400 magnification.
Being highly expressed in kidney tissues and in kidney cancer cells, we used the two cell lines to evaluate the impact of claudin-16 on biological functions. HEK293 cells were transiently transfected with claudin-16 expression vector and UMRC2 with anti-claudin-16 transgene, thus creating claudin-16 overexpressing HEK293 cells (HEK293-CL16exp) and claudin-16 knockdown UMRC2 cells (UMRC2-CL16KD).
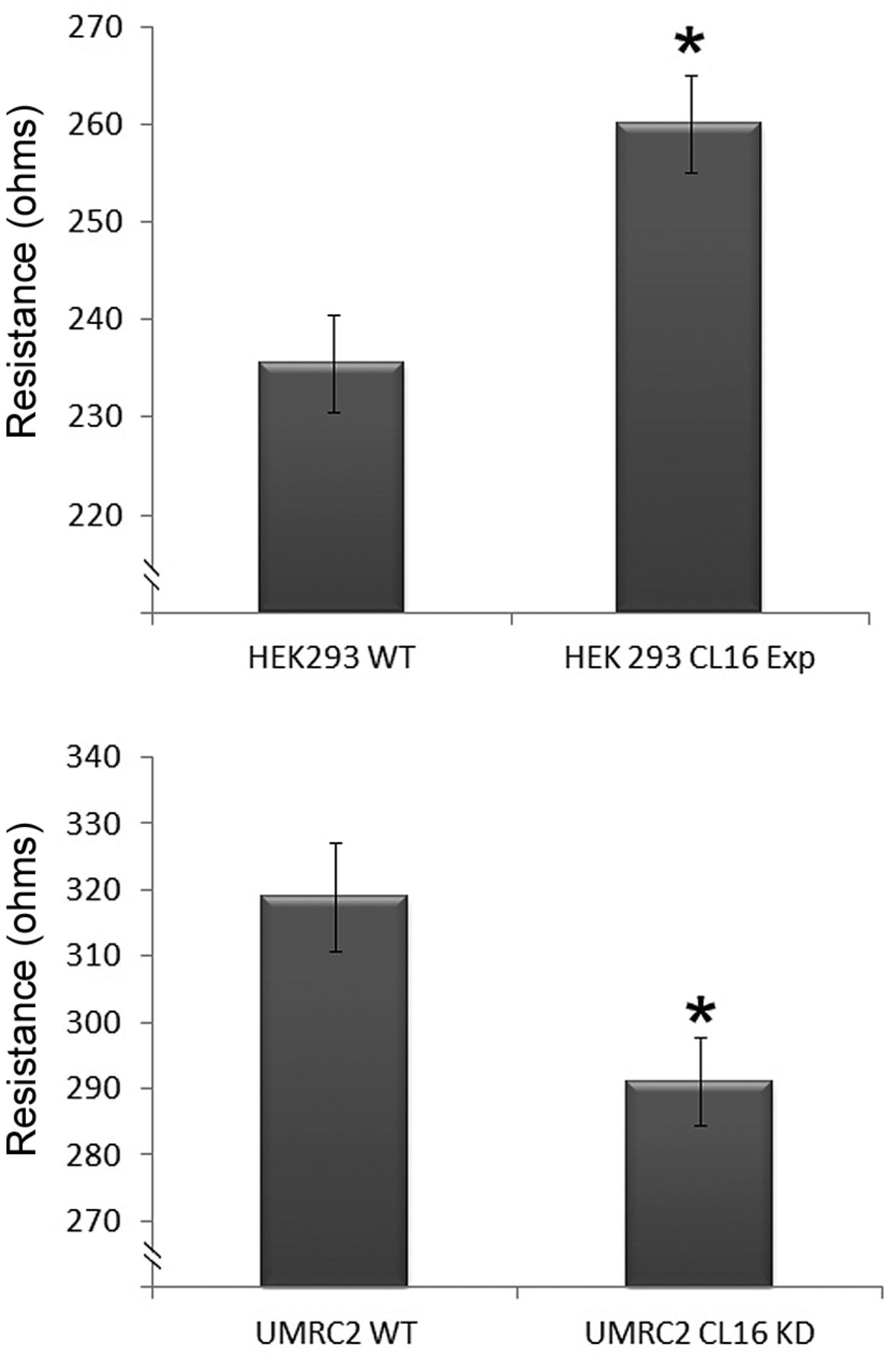
Using these genetically modified cells, we tested and compared the paracellular permeability to that of the control non-trasfected cells. As shown in Figure 3, HEK293-CL16exp cells had significantly higher transepithelial electric resistance (top). In contrast, knockdown of claudin-16 in UMRC2 cell resulted in an opposite effect (bottom). Using ECIS, we further demonstrated that knocking-down claudin-16 markedly reduced the adhesion of UMRC2 cells when compared with the wild-type cells (Figure 4A). This pivotal effect was seen over a broad range of frequencies evaluated in this study (1,000 Hz - 64,000 Hz) (Figure 4). Although overexpression of claudin-16 increased the adhesion of HEK293 cells (Figure 4B), the effect was nonetheless less prominent than that observed on UMRC2 cells.

Expression of claudin-16 in human kidney cell lines. UMRC2 cells strongly expressed claudin-16. HEK293 was negative for expression of this claudin.
Relative transcript levels of claudin-1, -5 and -16 in clear cell renal cell carcinoma.
Discussion
Claudins are an important group of transmembrane proteins controlling paracellular permeability and barrier function, and are essential components of tight junctions. They have also been shown to play important roles in the development and, in particular, the spread of cancer cells in breast (31-33), urinary bladder (34,35), colorectal (36-38), oesophageal (39), gastric (40, 41), prostate (42) and lung (43-45) cancer. There have been limited reports on the expression of claudin-1,-7 and -8 in CCRCC (20-23).
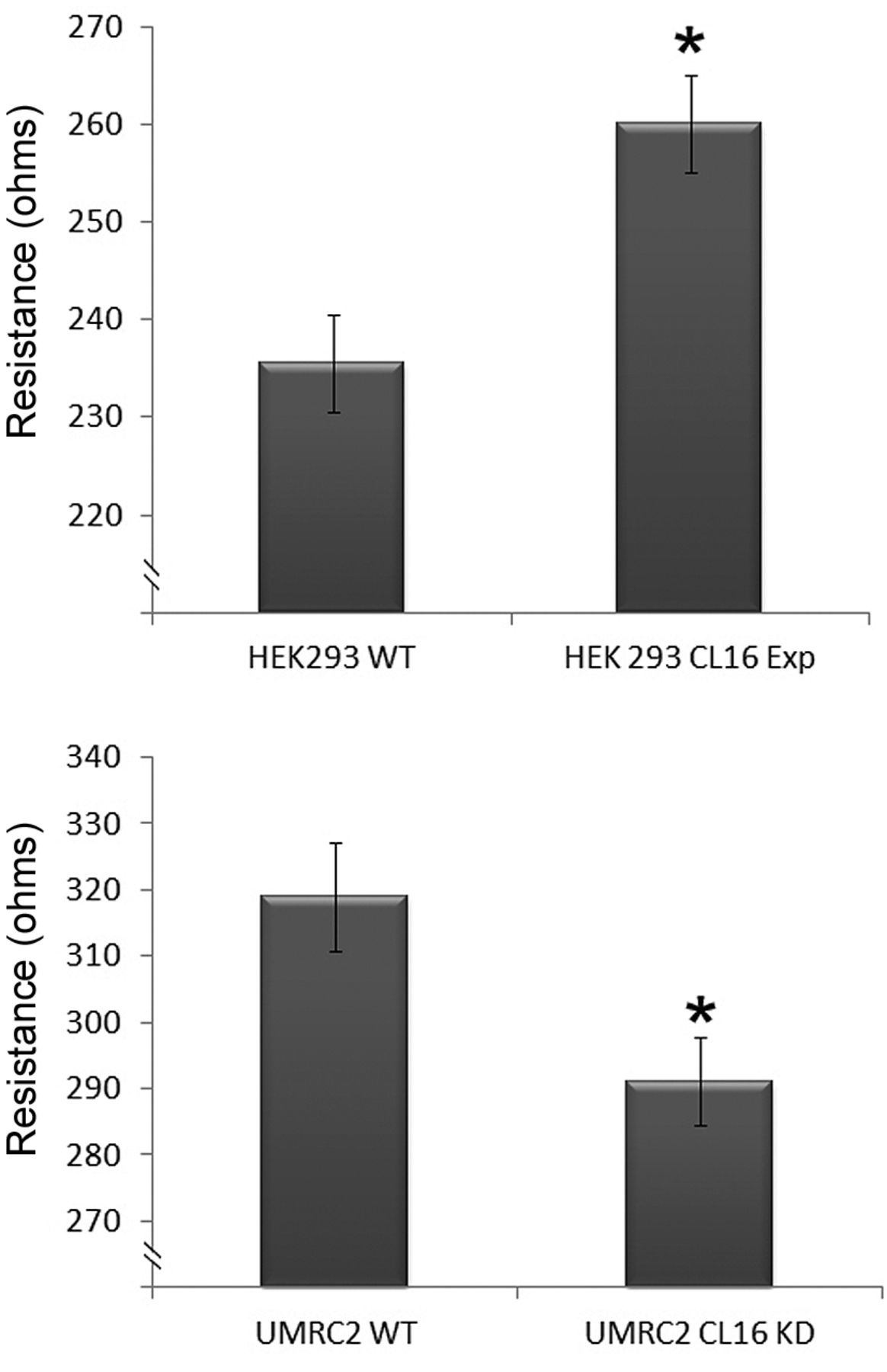
Effects of genetically manipulating the expression of claudin-16 on transepithelial electric resistance (TER) in HEK293 (top) and UMRC2 (bottom) cells. Overexpression of claudin-16 in HEK293 cells significantly increased TER. In contrast, knockdown of claudin-16 in UMRC2 recal carcinoma cells reduced TER of the cell. *p<0.05 vs. control cells.
In the present study, we showed that claudin-1 and claudin-16 are highly expressed in kidney tissues. Claudin-16 expression is particularly high in kidney tissues. In contrast, claudin-5 is virtually undetectable in kidney tissues. The observation on claudin-16 is very interesting. In patients with hypercalciuria and hypomagnecaemia syndrome, mutation of the claudin-16 protein results in increased permeability to both calcium and magesium in the ascending tubule of the loop of Henle of the kidney. Thus, it is not surprising that the levels of claudin-16 are high in kidney tissues. However, the current study has provided information for the first time that the molecule is also present in CCRCC and that the level is reduced in late stage tumours, namely TNM3, compared with TNM1 tumours. The present cohort is small, which has prevented us from reaching a stronger conclusions regarding the link between claudin-16 and long-term patient outcome. We aim to address this issue in future studies.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Manipulation of claudin-16 impacts the adhesion of kidney cancer UMRC2 (A) and HEK293 cell (B) cells. A: Knockdown of claudin-16 (UMRC2-CL16KD) markedly reduced the adhesion of the cancer cell line compared to wild-type cells (UMRC2-WT). This effect was seen over a broad range of frequencies monitored (ii: UMRC2-WT; iii: UMRC2-CL16KD). B: Effect of overexpression of claudin-16 on HEK293 cells. There was a modest effect on the cell adhesion after induced overexpression of claudin-16.
Although the findings on claudin-1 are not surprising and are in line with those previously reported, the observation that claudin-5 is largely undetectable in normal kidney and CCRCC tumors is interesting. Expression of claudin-5 is rather ubiquitous and frequently seen in epithelial cells, and in particular in vascular endothelial cells. The current study indicates that claudin-5 may play a far lesser role in normal kidney and RCC, in contrast to other tumour types, namely breast and colorectal cancer.
The role of claudin-16 in kidney cancer cells is not clear. Herein, by using a pair of cell lines, negative and positive for claudin-16, respectively, we were able to create sublines that differentially expressed claudin-16. Our results clearly demonstrated that the level of claudin-16 is inversely associated with barrier function, namely knockdown of claudin-16 in UMRC2 cells resulted in cells with reduced resistance, and forced expression of claudin-16 in HEK293 cells caused the opposite. The same was largely seen in the ECIS. Thus, this would suggest that claudin-16 in RCC acts in a control mechanism for barrier function. Together with the observations of the clinical cohort, claudin-16 may act as a suppressor mechanism in the development and progression of CCRCC. Clearly, more work is required to further explore how this cell function may be linked to the clinical course of the tumour type.
In conclusion, the current study demonstrated that claudin-1 and claudin-16 are predominately expressed in human kidney tissues and CCRCC. Claudin-16, which is highly expressed in these tissues, plays an important role in the barrier properties of RCC cells. Both claudin-1 and claudin-16 may play an important in the biology and clinical course of RCC.
Acknowledgments
The Authors wish to thank Cancer Research Wales, the Albert Hung Foundation, Yiling Foundation and the Welsh Life Science Fund (Ser Cymru) for supporting their work. Dr Wenjin Men is a recipient of Cardiff University’s China Medical Scholarship.
- Received October 24, 2014.
- Revision received November 5, 2014.
- Accepted November 7, 2014.
- Copyright© 2015, International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved