

Abstract
Expression of estrogen receptor (ER), progesterone receptor (PR), and the human epidermal growth factor receptor 2 (HER2) can subdivide breast carcinomas into clinically meaningful classes. Cancers lacking expression of all three of these receptors (triple-negative breast cancer; TNBC) is of particular interest for molecular research because these tumors currently have no effective targets for therapy. Furthermore, TNBCs are relatively more prevalent among African-American women and can account for some of the health disparities associated with breast cancer. We approached a molecular understanding of how TNBC differs from ER+ breast cancer through a comprehensive gas chromatography (GC)-mass spectrometry (MS) and liquid chromatography (LC)/MS/MS-based and unbiased metabolomic analysis of a series of breast carcinomas from African-American patients. Remarkably, global metabolomic profiling of tumor tissues identified a total of 418 distinct metabolites, out of which 133 (31.8%) were shown to differ between the ER+ and TNBC tumors with statistical probability of p<0.05. Specific biochemical pathways affected included those reflecting general increases in energy metabolism and transmethylation in the TNBC tumors when compared to ER+ tumors. Additionally, biochemicals associated with increased proliferation, redox balance and the recently proposed oncometabolites, sarcosine and 2-hydroxyglutarate, were also detected at higher levels in the TNBC versus ER+ tumors. These studies demonstrate that TNBC tumors have metabolic signatures that distinguish them from ER+ tumors and suggest that distinctive metabolic characteristics of these tumors might offer new targets for treatment.
- Metabolomic
- triple-negative breast cancer
- African-American women
Breast cancer is a disease that shows striking diversity in terms of tumor histology, highly variable clinical behavior and response to therapy (1). This diversity probably reflects different molecular changes and possibly different cellular origins. African-American women have more advanced-stage disease at diagnosis and higher relative incidence rates of triple-negative breast cancer (TNBC), which lack estrogen receptor (ER) and progesterone receptor (PR) expression as well as human epidermal growth factor receptor 2 (HER2) amplification (2, 3). TNBCs tend to be higher in grade, have lymph node involvement at diagnosis and are biologically more aggressive. Clinically, TNBCs are associated with a relatively high rate of recurrence and distant metastasis, and poor overall survival (4). Clearly there is a need to better understand the basis of TNBC and to develop effective treatments for this aggressive subtype of breast cancer.
Although, extensive molecular and genomic studies (5-15) have been carried out to understand the complexity of breast cancer and to identify markers that can be therapeutically targeted, little is known about the global metabolomic alterations that characterize TNBC progression. Metabolic profiling is of special importance in cancer biology due to profound changes in central metabolism associated with many tumors as established early by biochemical studies (16, 17). Metabolites define the phenotype of all cells, accordingly, metabolite levels provide the most telling readout of cell function and dysfunction (18-20). The relation of the metabolome to phenotype may also offer increased insights into functional changes associated with pharmacological or nutritional intervention (21).
In the present study, we utilized a combination of gas chromatography-mass spectrometry (GC/MS) and liquid chromatography-mass spectrometry (LC/MS/MS)-based comprehensive and unbiased metabolomic profiling of tissue specimens from TNBC relative to estrogen receptor-positive (ER+) breast carcinomas in African-American patients. Starting with non-targeted analysis as an unbiased metabolomic approach, the possible biomarkers were selected by using statistically-based pattern analysis and then placed into the context of metabolic pathways and other research data (i.e. gene methylation data). Herein, we were able to identify several potent biomarkers for TNBC through qualitative and quantitative approaches that correlate with known and proposed mechanisms that differentiate TNBC from ER+ tumors.
This article has great significance in that together with genomics, transcriptomics, and proteomics, metabolomics can provide insights into understanding associated mechanisms and identifying pathways that are perturbed in TNBC that are potential targets for new therapies of these aggressive carcinomas.
Materials and Methods
Tissue samples and clinical data. For the study, we recruited and consented 30 African-American patients who were diagnosed with breast cancer (15 TNBC and 15 ER+) from the Department of Surgery at Howard University Hospital (HUH) and prior to beginning chemotherapy, radiation therapy or hormonal therapy. The fresh breast cancer tissue samples were collected along with clinical/pathological information with Institutional Review Board approval (IRB-12-CMED-39) from the Pathology Department at HUH and all samples were coded to preserve patient confidentiality. The mean age at diagnosis was significantly younger for the TNBC group compared with the ER+ group (53 versus 56 years, respectively). Patients in the TNBC group were more likely to have ductal carcinoma (100% versus 73%, p<0.05); grade III tumors (40% versus 20%, p<0.05); poorly differentiated tumors (100% versus 73%, p<0.05), and the mean tumor size was slightly larger (3.0 versus 2.6 cm, respectively). Only one-fifth of the TNBCs were <2.0 cm at presentation, whereas almost one-third of the ER+ carcinomas were <2.0 cm (100% versus 20%, respectively, p<0.05). The rate of node positivity was slightly higher in the TNBC group compared with the ER+ group (53% versus 40%, respectively). The recurrence rate was higher in patients with TNBC (60% versus 20%, p<0.05), whereas the survival rate was lower (47% versus 87%, p<0.05).
Sample preparation. Tumor tissue samples (100 mg each) were homogenized in a minimum volume of water and 100 μl withdrawn for subsequent analyses. Using an automated liquid handler (Hamilton LabStar, Salt Lake City, UT, USA), protein and nucleic acid were precipitated from the homogenates with methanol that contained multiple standards to report on extraction efficiency, a protocol optimized to allow maximum recovery of small molecules on multiple MS platforms. The resulting extract was divided into four equal fractions: one for analysis by UHPLC/MS/MS (positive mode), one for UHPLC/MS/MS (negative mode), one for GC/MS, and one banked for reserve. Samples were dried under nitrogen and vacuum desiccated (TurboVap®; Zymark, Hopkinton, MA, USA) to remove the organic solvent. Samples were then prepared for the appropriate instrument, either UHPLC/MS/MS or GC/MS.
Metabolite analysis. Unbiased, global metabolomic profiling analysis was performed by Metabolon (Durham, NC, USA) as previously described (22-25).
UHPLC/MS/MS. The LC/MS portion of the platform was conducted using a Waters ACQUITY UHPLC (Waters, Millford, MA, USA) and a Thermo-Finnigan linear trap quadrupole mass spectrometer (Thermo Fisher Scientific, Inc., Waltham, MA, USA), which consisted of an electrospray ionization source and linear ion-trap mass analyzer. The dried sample extract was reconstituted in acidic (50 μl of 0.1% formic acid in water) or basic (50 μl of 6.5 mM ammonium bicarbonate in water, pH 8) LC-compatible solvents, each of which contained eight or more injection standards at fixed concentrations to ensure injection and chromatographic consistency. Aliquots from either acidic or basic reconstitution were analyzed using (acidic) positive ion optimized conditions or (basic) negative ion optimized conditions in two independent injections using separate dedicated columns. Extracts reconstituted in acidic conditions were gradient eluted using water and methanol containing 0.1% formic acid, while the basic extracts, which also used water/methanol, contained 6.5 mM ammonium bicarbonate. The MS analysis scanned compounds with a mass/charge ratio of 99-1000 m/z and alternated between MS and data-dependent MS/MS scans using dynamic exclusion. Raw data files are archived and extracted as described below.
GC/MS. The samples destined for GC/MS analysis were re-dried under vacuum desiccation for a minimum of 24 hours prior to being derivatized to a final volume of 50 μl using equal parts bistrimethyl-silyl-triflouroacetamide and solvent mixture acetonitrile: dichloromethane:cyclohexane (5:4:1) with 5% triethylamine at 60°C for one hour. The GC samples were separated on a 5% phenyldimethyl silicone column with helium as the carrier gas, and a temperature ramp from 60°C to 340°C in a 16-minute period. GC-separated samples were analyzed on a Thermo-Finnigan Trace DSQ (Thermo Fisher Scientific, Inc.) fast-scanning single-quadrupole MS at unit mass resolving power with electron impact ionization and a 50-750 atomic mass unit scan range. The instrument was tuned and calibrated for mass resolution and mass accuracy on a daily basis. The information output from the raw data files were automatically extracted as discussed below.
Quality assurance (QA)/quality control (QC). For QA/QC purposes, additional samples were included with each day’s analysis. These samples included extracts of a pool created from a small aliquot of each experimental sample that served as “technical replicates” for the study, as well as process blanks. QC samples were spaced evenly among the injections and all experimental samples were randomly distributed throughout the run. A selection of QC compounds was added to every sample for chromatographic alignment, including those under test. These compounds were carefully chosen so as not to interfere with the measurement of the endogenous compounds.
Data extraction and compound identification. Metabolites were identified by automated comparison of the ion features in the experimental samples against a reference library of more than 2400 purified chemical standard entries that included retention time, molecular weight (m/z), preferred adducts, and in-source fragments as well as associated MS spectra (from LC- and GC-based platforms), and then curated by visual inspection for quality control using software developed at Metabolon (26). These systems are built on a web-service platform utilizing Microsoft’s NET technologies, which run using high-performance application servers and fiber-channel storage arrays in clusters to provide active failover and load-balancing.
Statistical analysis. Experimental samples and controls were randomized across a one-day platform run. Missing values (if any) are assumed to be below the level of detection. However, biochemicals that were detected in all samples from one or more groups but not in samples from other groups were assumed to be near the lower limit of detection in the groups in which they were not detected. In this case, the lowest detected level of these biochemicals was imputed for samples in which that biochemical was not detected. Following log transformation and imputation with minimum observed values for each compound, statistical analysis was performed using “R” (http://cran.r-project.org/) that is a freely available, open-source software package. Welch’s two-sample t-test was performed to identify biochemicals that differed significantly between experimental groups. Multiple comparisons were accounted for by estimating the false discovery rate using q-values (27).
Metabolite correlations with clinical parameters. Using the median-scaled imputed data, a correlation analysis was conducted between multiple clinical parameters, sarcosine, 2-hydroxyglutarate, and the other 409 measured metabolites, in order to determine the strength of the linear relationships between the metabolites and the various clinical parameters. A p-value was calculated testing the null hypothesis that the correlation between the variables was equal to zero. Storey Q values were calculated to estimate the proportion of false-positives.
Results
Global unbiased metabolomic analysis. Since the 1920s when Otto Warburg reported his observations that tumor cells preferentially metabolized glucose via anaerobic means, even in the presence of adequate oxygen (aerobic glycolysis), cancer cell metabolism has been a focus of many investigators. While cancer genomics has revealed many new mechanisms of malignant transformation and progression to metastasis, gene expression changes alone cannot fully account for differences that effectively distinguish cancer subtypes such as TNBC from ER+ tumors. Hence, we incorporated global, untargeted metabolomic analyses of tumor tissues in order to distinguish metabolic differences that may help elucidate mechanisms that contribute to the notably distinct clinical phenotypes observed between ER+ and TNBC subtypes. In the current study, global metabolomic profiling of tumor tissues identified a total of 418 distinct biochemicals, of which 133 (31.8%) were shown to differ with statistical significance (p<0.05) between the ER+ and TNBC tumors. A closer evaluation of specific biochemical pathways revealed general increases in energy metabolism, transmethylation, and coordinate elevations of biochemicals associated with increased proliferation and redox balance in TNBCs when compared to ER+ tumors.
Energy metabolism. Several energy metabolism pathways were shown to be significantly elevated in TNBC versus ER+ tumors including amino acids and γ-glutamino acids. Eight out of the 20 amino acids detected in this study were significantly elevated in TNBC samples compared to samples from patients with ER+ tumors, the most notably increased being aspartate (30.79-fold) and asparagine (16.81-fold), which reached statistical significance (p<0.05). Additionally, glutamate (3.13-fold), phenylalanine (3.25-fold), methionine (4.56-fold), and the branched chain amino acids isoleucine (3.34-fold), leucine (3.39-fold) (all p<0.05), and valine (7.77-fold; p=0.08) were also observed to be elevated in the TNBC compared to ER+ tumors.
One mechanism for intracellular transportation of amino acids is via the γ-glutamyl transpeptidase-mediated catalytic reaction of amino acids with glutathione to form γ-glutamyl amino acids (28), which then traverse the cell membrane to release amino acids intracellularly and in the process regenerate glutathione through the 5-oxoproline intermediate (Figure 1). Of the seven γ-glutamyl amino acids identified through metabolomic profiling in the current study, six were significantly elevated in TNBC compared to ER+ cancer. The relative concentrations of γ-glutamylvaline (7.90-fold), γ-glutamylleucine (8.44-fold), γ-glutamylisoleucine (8.67-fold), γ-glutamylmethionine (2.90-fold), γ-glutamylphenylalanine (5.91-fold) and γ-glutamyltyrosine (4.44-fold) were greater in TNBC versus ER+ breast cancer (Figure 1 and Table I). Additionally, out of the 48 dipeptides identified in this study (not shown), all were shown to be elevated in TNBC versus ER+ tumors and 39 of these reached statistical significance (p<0.05, range: 1.80- to 9.97-fold than in ER+). In fact, TNBC biochemical profiles showed elevated levels of every amino acid detected compared to levels in ER+ tumors (7 out of 20 statistically significant, p<0.05; 8 out of 20 trending significance, 0.05<p<0.1), with the exception of glutamine, which was slightly decreased in TNBC versus ER+ (p=0.53) (Table I).
Specific TCA cycle intermediates identified in TNBC samples were observed to be increased in a manner consistent with amino acid anaplerosis that parallels observations indicative of γ-glutamyl amino acid uptake and elevated amino acid levels in general versus ER+ levels. For the TCA cycle, TNBCs had increased levels of succinate (4.94-fold, p=0.06), fumarate (7.85-fold, p<0.05) and malate (3.91-fold, p=0.15), with the greatest increase observed with fumarate in relation to ER+ samples (Figure 2). This supports the idea of amino acid anaplerosis and specifically implicates the branched chain amino acids (BCAAs) valine and isoleucine, as well as aspartate, methionine and phenylalanine, all of which are potential contributors to the substantial increase observed in fumarate in the TNBC samples. Interestingly, a notable decrease in citrate was observed in TNBC samples (0.33-fold), although not statistically significant (p=0.34), which could be explained by an increase in Warburg metabolism (shunting glucose to lactate and limiting pyruvate incorporation into the TCA cycle via acetyl-CoA) and/or utilization of citrate in fatty acid synthesis, both of which are associated with cancer.

Pathway diagram showing the γ-glutamyl cycle and points of amino acid anaplerotic contribution with related box plots of levels of γ-glutamyl cycle intermediates and amino acids. p-Values are shown as insets for the comparison TNBC vs. ER+. Metabolites with significant elevation in TNBC vs. ER+ (p<0.05) are shown with solid arrow. NS: Not significant, p>0.1; GCS: γ-glutamylcysteine synthetase; GGT: γ-glutamyl transferase; GS: Glutathione synthetase. Box plots: +: Mean value; ---: Median value; o: Outliers; Top of box: Upper quartile; Bottom of box: Lower quartile; Top of whisker: Maximum of distribution; Bottom of whisker: Minimum of distribution. Sample cohorts are color coded: Light grey=ER+, Dark grey=TNBC. Data are shown as scaled intensity for each metabolite.
TNBCs were also shown to have increased glycolytic intermediates and lactate production compared to ER+ tumors (Table II). The metabolic profiles of TNBC tumors showed increases in several intermediates of glycolytic metabolism including glucose, glucose-6-phosphate, fructose-6-phosphate, fructose-1,6-bisphosphate, 2-phosphoglycerate, pyruvate and lactate (Figure 3). Although only differences between TNBC and ER+ relative concentrations for glucose-6-phosphate, pyruvate and lactate reached levels that were of trending significance (0.05<p<0.1), the observation that nearly all glycolytic intermediates were elevated in TNBC versus ER+ samples suggests an increase in Warburg metabolism in the TNBCs. These data are in good agreement with the decrease in citrate mentioned above (Figure 2). Increased glucose levels in TNBC tumors were also associated with increased markers of glycogen mobilization towards glucose metabolism: maltotetraose (11.23-fold, p=0.07), maltotriose (55.16-fold, p=0.14, maltose (23.58-fold, p=0.05) and glucose-1-phosphate (1.96-fold, p=0.42), although the variance within the TNBC group for these biochemicals was notably broad (data not shown).
TNBCs were shown to have increased amino acids and glycolytic precursors for TCA cycle energetics when compared to ER+ tumors, and therefore, it would be anticipated that the NAD+ requirements of TNBCs would be much greater than that of ER+ tumors. In fact, metabolomic profiles of TNBCs indicated two mechanisms of NAD+ production that are elevated over ER+ tumors; namely tryptophan metabolism to NAD+ and an increase in the NAD+ salvage pathway (Figure 4). TNBC samples contained elevated levels of tryptophan (3.06-fold, p=0.14) and its metabolic intermediates in NAD+ biosynthesis, kynurenine (6.90-fold, p<0.05) and quinolinate (7.99-fold, p<0.05). TNBCs also showed increased levels of the NAD+ salvage pathway biomarker nicotinamide (3.49-fold, p<0.05), further reflecting the increased energy demands of TNBC.
Redox balance. It is interesting to note that the most increased γ-glutamyl amino acids are the BCAA conjugates of valine, leucine and isoleucine, which are also elevated in TNBCs (Table I). As mentioned above, uptake of amino acids through the γ-glutamyl pathway is associated with regeneration of the redox mediator, glutathione. TNBCs had also increased reduced glutathione (GSH; 8.56-fold, p=0.14), 5-oxoproline (2.81-fold, p=0.19) and oxidized glutathione (GSSG; 1.58-fold, p=0.08) indicating a trend for increased glutathione synthesis, although these increases did not reach statistical significance (Figure 1). The majority of GSH synthesis intermediates resulting from methionine metabolism were significantly elevated in TNBC samples versus ER+ tumors, including methionine (4.56-fold, p<0.05), S-adenosylhomocysteine (SAH, 4.10-fold, p<0.05), homocysteine (3.93-fold, p<0.05) and cysteine (5.02-fold; p=0.07) (Table I). The fact that all of the detected components of this pathway were elevated in TNBCs over ER+ tumors strongly suggests a redox advantage in TNBC tumors.
Table of all identified glutathione-related biochemicals and amino acids. Biochemicals related to glutathione biosynthesis pathways and amino acids are shown with corresponding fold-change values and p-values for the specific comparison TNBC vs. ER+. p≤0.05 was accepted as significant; p-value trends of 0.05<p<0.10 identified biochemicals approaching significance. Platform: LC: Liquid chromatography; GC: Gas chromatography; HMDB: Human Metabolome Database.
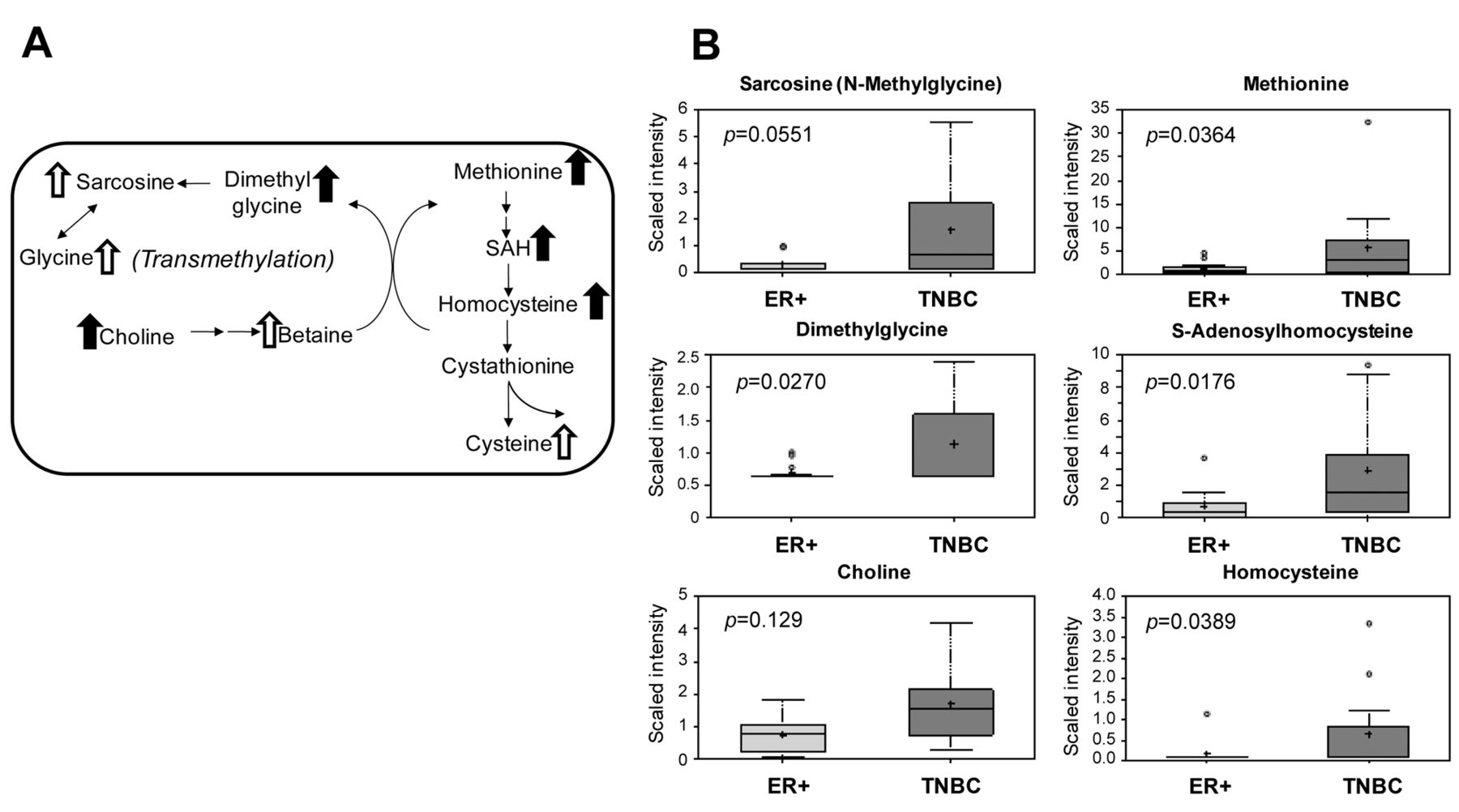
Transmethylation pathway. TNBCs showed increased levels of methionine (4.56-fold, p<0.05), which parallel increased methionine uptake through the γ-glutamyl pathway as mentioned above (Table I, γ-glutamylmethionine, 2.9-fold). Statistically significant elevations of SAH and homocysteine were also observed in TNBCs, potentially reflecting an increase in methionine accumulation and metabolism through multiple pathways. Accordingly, choline (2.26-fold, p<0.05) and dimethyl-glycine (1.62-fold, p<0.05), along with increases in betaine (p=0.27), sarcosine (4.64-fold, p=0.05) and glycine (3.89-fold, p=0.09), collectively indicated an increase in the transmethylation pathway in TNBC (Figure 5). Hypermethylation in the TNBC samples is further supported by statistically significant increases in the concentrations of 4-methylglutamate, 2-methylbutyroylcarnitine, dimethylarginine, N1-methyladenosine, N1-methylguanosine, N2,N2-dimethylguanosine, 5-methyluridine, methyl-alpha-glucopyranoside, 5-methylthioadenosine, and methylphosphate (trending significance, 0.05<p<0.1) (Table III).
Table of all identified methylation-related biochemicals. Biochemicals are shown with corresponding fold-change values and p-values for the specific comparison TNBC vs. ER+. p≤0.05 was accepted as significant; p-value trend of 0.05<p<0.10 identified biochemicals approaching significant. Platform: LC: Liquid chromatography; GC: Gas chromatography; HMDB: Human Metabolome Database; SDMA: Symmetrical dimethylarginine; ADMA: Asymmetric dimethylarginine.
Arginine, polyamine and collagen metabolism. Consistent with increased amino acid uptake and protein catabolism mentioned above, TNBCs had increased levels of several arginine metabolites, indicating changes in multiple tumor-relevant pathways. Intermediates of arginine incorporation into the urea cycle were significantly elevated including citrulline (16.22-fold, p<0.05) and the end-product urea (11.93-fold, p<0.05), suggesting a substantial increase in pro-inflammatory signaling via nitric oxide production and arginine metabolism for other biofunctional needs (Figure 6). Indeed, putrescine levels were also greatly elevated in TNBCs when compared to ER+ samples (15.09-fold, p<0.05), indicating an increased proliferative signaling associated with this polyamine (29). Intermediates of arginine incorporation into energy metabolism pathways were also elevated in TNBC samples. Creatine (2.50-fold) and creatinine (1.48-fold), metabolites of arginine that feed into energy metabolism, tended towards a significant increase in TNBC versus ER+ breast cancer, further reflecting the increased energy needs already mentioned above for aggressive TNBC (Figure 6). In addition to arginine metabolites, metabolic breakdown products of extracellular matrix (ECM, i.e. collagen) were also increased in TNBC samples, including proline (4.08-fold, p=0.05), trans-4-hydroxyproline (1.33-fold, p=0.43) and proline-hydroxy-proline (pro-hydroxy-pro, 1.22-fold, p=0.33); although the latter two did not reach statistical significance. These changes may reflect greater tissue remodeling in advanced/aggressive TNBC (data not shown).
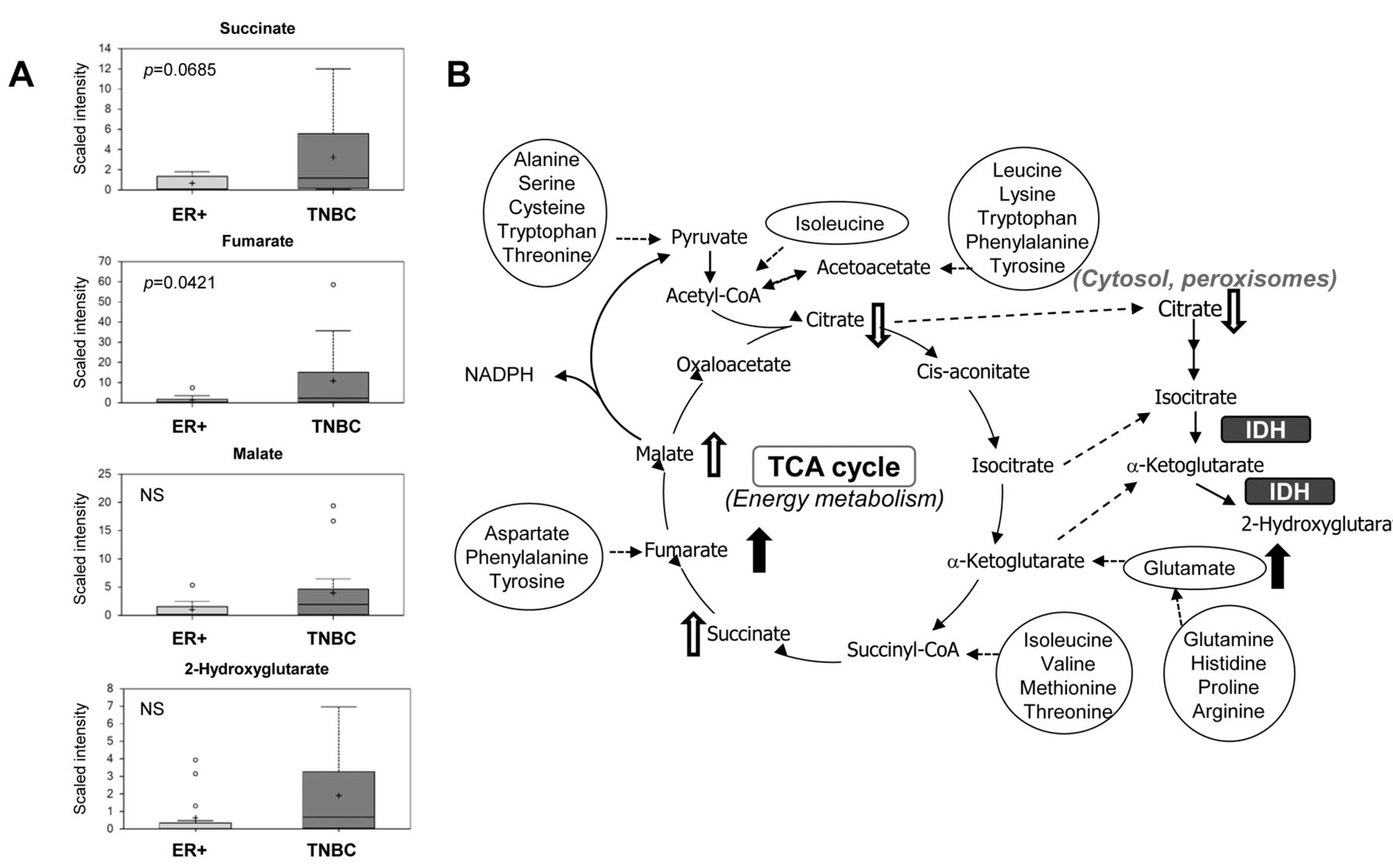
Pathway diagram showing the tricarboxylic acid (TCA) cycle and points of amino acid anaplerotic contribution with related box plots. A: Box plots showing TCA cycle intermediates. p-Values are shown as insets for the comparison of TNBC vs. ER+. NS: Not significant, p>0.1. B: Pathway diagram showing the TCA cycle and multiple points where amino acids contribute anaplerotically to pathway energetics. Metabolites with significant elevation in TNBC vs. ER+ (p<0.05) are shown with solid arrows; trending elevations or reductions shown with open arrow (0.05<p<0.1). IDH: Isocitrate dehydrogenase. Box plots: +: Mean value; ---: Median value; o: Outliers; Top of box: Upper quartile; Bottom of box: Lower quartile; Top of whisker: Maximum of distribution; Bottom of whisker: Minimum of distribution. Sample cohorts are color coded: Light grey=ER+, Dark grey=TNBC. Data are shown as scaled intensity for each metabolite.
Finally, TNBC samples had decreased lysolipids and increased glycerophospholipids, potentially indicating changes in their utilization and incorporation into membrane biosynthesis. Of the many biochemical pathways that was shown to be up-regulated in TNBC versus ER+ tumor samples, the only biochemical category that showed a prominent decrease in TNBC was grouped within the lysolipid sub-pathway. 2-Palmitoleoylglycerophosphoethanolamine, 2-oleoylglycero-phospho-ethanolamine, 2-linoleoylglycerophosphoethanolamine, 2-arachidonoyl-glycero-phosphoethanol-amine, 2-docosapentaenoylglycerophosphoethanolamine and 2-docosahexaenoyl-glycerophosphoethanolamine were all significantly decreased in TNBCs compared to ER+ tumors (data not shown). Additionally, the glycerophospholipids glycerol-3-phosphate (6.11-fold, p=0.11) and glycero-phosphorylcholine (3.04-fold, p<0.05) were both elevated in TNBC samples. Taken together, these data indicated that membrane remodeling, and perhaps biosynthesis, differs between ER+ and TNBC tumors. The fact that all the significantly decreased lysolipids in TNBCs are of the “2-acyl” isoform may suggest a phospholypase A2-specific (PLA2 versus PLA1) dysfunction associated with the formation of these lysolipids in TNBC, which could relate to inflammatory differences between the two tumor types.
Table of all identified glycolysis, glycogen metabolites and tricarboxylic cycle related (TCA) biochemicals and amino acids. Biochemicals are shown with corresponding fold-change values and p-values for the specific comparison TNBC vs. ER+. p≤0.05 was taken as significant; p-value trend of 0.05<p<0.10 identified biochemicals approaching significance. Platform: LC: Liquid chromatography; GC: Gas chromatography; HMDB: Human Metabolome Database.

Pathway diagram showing the glycolytic and glycogenolysis pathways with related box plots. A: Pathway diagram showing glycolysis from glucose to lactate and the affiliation with glycogenolysis. Intermediates with elevations of trending significance (0.05<p<0.1) when comparing TNBC to ER+ are shown with open arrows. B: Relevant box plots showing multiple intermediates of the glycolytic pathway. p-Values are shown as insets for the comparison TNBC vs. ER+. NS: Not significant, p>0.1; TCA: Tricarboxylic acid. Box plots: +: Mean value; ---: Median value; o: Outliers; Top of box: Upper quartile; Bottom of box: Lower quartile; Top of whisker: Maximum of distribution; Bottom of whisker: Minimum of distribution. Sample cohorts are color coded: Light grey=ER+, Dark grey=TNBC. Data are shown as scaled intensity for each metabolite.
Metabolite correlations with clinical parameters. In order to determine the strength of the linear relationships between the metabolites and the various clinical parameters, a correlation analysis was conducted between multiple clinical parameters, sarcosine, 2-hydroxyglutarate, and the other 409 measured metabolites. In our data set, although 2-hydroxyglutarate differentiated TNBC from ER+ tumors (3.02-fold, p=0.08), 2-hydroxyglutarate did not significantly correlate with death (3.21-fold than in survivors, p=0.18). Sarcosine, however, had a tendency to be correlated with mortality (3.84-fold than in survivors, p=0.07) and dimethylglycine (also in the transmethylation pathway) was significantly correlated with death versus survival (2.44-fold than in survivors, p=0.02) (Table IV). Interestingly, elevated dipeptides (almost every one detected) were also associated with death, which may indicate protein degradation associated with increasing amino acid utilization for fuel or from tissue remodeling (Tables V and VI). Moreover, changes in glucose and glycolytic intermediates also correlated with death, further emphasizing the potential role of metabolism in breast cancer prognosis and therefore aggressiveness.
Discussion
Possible explanations for higher mortality and lower survival rates from TNBC among African-American women include factors associated with lower socioeconomic status and later tumor stage at diagnosis (30, 31). However, several studies have now shown race-dependent differences in survival even after adjustment for variables such as stage at diagnosis, access to health care, treatment, co-morbid illness, and marital status (3,32-37). This might be explained by differences in the phenotypes of breast carcinomas seen across racial groups, and data indicate that African-American women have a relatively higher frequency of cancer that is high grade and ER-/PR- (37-41). Thus, there are indications that breast carcinomas in African-American patients are, in general, biologically more aggressive than those in Caucasian patients (42-44).
However, the mechanisms that underlie these health disparities associated with breast cancer in African-Americans remain largely unknown, particularly for TNBC, which is often refractory to conventional therapies. To study TNBC development and progression in African-American women, and to obtain insights into bioprocesses that could possibly be associated with perturbed metabolism, we conducted a GC/MS and LC/MS/MS-based comprehensive and unbiased metabolomic profiling of tissue specimens from TNBCs relative to ER+/PR+ breast carcinomas in African-American patients.
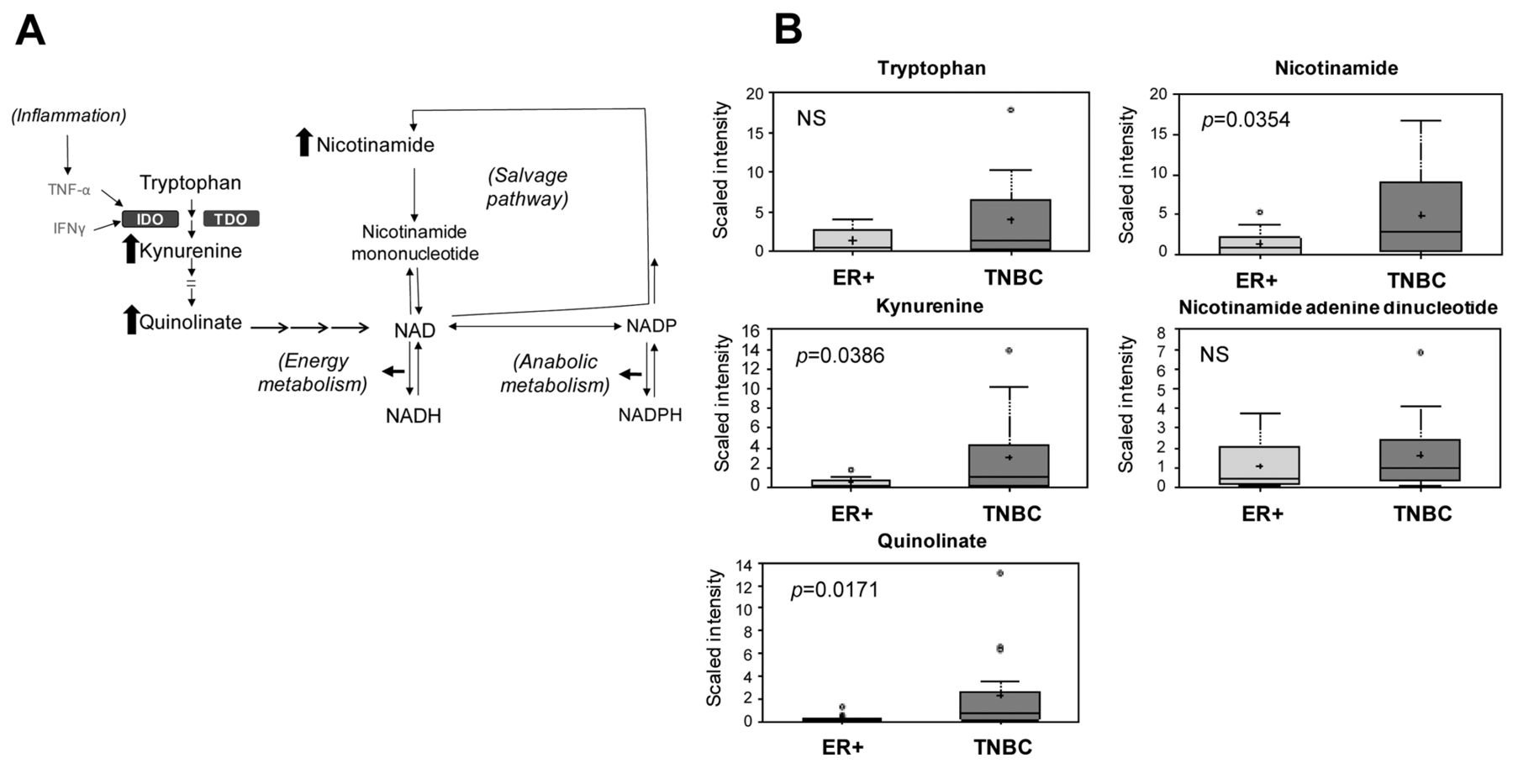
Pathway diagram showing multiple mechanisms for NAD+ synthesis with related box plots. A: Pathway diagram showing NAD+ synthesis from tryptophan (left side), and from the NAD-salvage pathway (right side). Metabolites with significant elevation in TNBC vs. ER+ (p<0.05) are shown with solid arrows. B: Relevant box plots showing multiple metabolites associated with NAD+ synthesis. p-Values are shown as insets for the comparison of TNBC vs. ER+. NS: Not significant, p>0.1; TNF-α: Tumor necrosis factor alpha; IFN-γ: Interferon gamma; IDO: Indoleamine 2,3-dioxygenase; TDO: Tryptophan 2,3-dioxygenase. Box plots: +: Mean value; ---: Median value; o: Outliers; Top of box: Upper quartile; Bottom of box: Lower quartile; Top of whisker: Maximum of distribution; Bottom of whisker: Minimum of distribution. Sample cohorts are color coded: Light grey=ER+, Dark grey=TNBC. Data are shown as scaled intensity for each metabolite.
In our study, global metabolomic profiling of tumor tissues identified a total of 418 distinct biochemicals, out of which 133 (31.8%) were shown to differ with statistical significance (p<0.05) between the ER+ and TNBCs. A closer evaluation of specific biochemical pathways revealed general increases in energy metabolism, transmethylation, with elevations of biochemicals associated with increased proliferation and redox balance in the TNBCs when compared to ER+ tumors. Interestingly, a notable decrease in citrate was observed in TNBC samples (0.33-fold that in ER+), although not statistically significant, which could be explained by an increase in Warburg metabolism (shunting glucose to lactate and limiting pyruvate incorporation into the TCA cycle via acetyl-CoA) or utilization of citrate in fatty acid synthesis, both of which are associated with cancer (45). Some of these observations, such as up-regulation of fatty acid synthesis, are consistent with previous reports using breast and prostate cell lines (42, 46). Borgan et al. were able to sub-classify invasive ductal carcinoma with ER+ disease, with the samples in one sub-group, designated A2, having significantly lower glucose and higher alanine levels than the other luminal A samples (14). This would suggest a higher glycolytic activity in these tumors. However, the data in our study are in agreement with our previous studies (47) and potentially support a metabolic foundation for gene hypermethylation in TNBC based on increased methionine uptake and acceleration of the transmethylation pathway. We previously showed increased gene methylation frequency in aggressive ER-/PR- breast cancer that distinguishes disease in African-Americans from Caucasian-associated disease in women less than 50 years of age. This is consistent with the current findings that identified significant elevations in several methylated nucleic acids and amino acids (48).
Consistent with our data, numerous metabolites such as glucose, lactate, lipids, choline, and amino acids, were shown to correlate with breast cancer (13, 49-55). Studies reported choline as one of the most prominent metabolites in cell biology and is invariably associated with increased activity of tumor cell proliferation in breast cancer. This might lead us to an explanation to the aggressiveness of TNBC.
Table of all identified methylation-related biochemicals and their correlation with death vs. survival. Biochemicals are shown with corresponding fold-change values and p-values for the specific comparison TNBC vs. ER. p≤0.05 was taken as significant correlation with death vs. survival; p-value trend of 0.05<p<0.10 identified biochemicals approaching significant correlation with death vs. survival. Platform: LC: Liquid chromatography; GC: Gas chromatography; HMDB: Human Metabolome Database; SDMA: Symmetrical dimethylarginine; ADMA: Asymmetric dimethylarginine.
Moreover, in the present study, elevation of the oncometabolite sarcosine in parallel with 2-hydroxyglutarate has been reported separately for other cancer types (24). The results of our study may indicate novel biomarkers that delineate TNBCs from ER+ tumors in African-American women. Sreekumar et al. screened 262 clinical samples related to prostate cancer for metabolites and defined sarcosine as a potential biomarker of interest (24). Sarcosine, an N-methyl derivative of the amino acid glycine, formed by the enzymes glycine N-methyl transferase or dimethylglycine dehydrogenase, and converted back into glycine via sarcosine dehydrogenase. As prostate cancer progresses towards metastatic disease, amino acid metabolism along nitrogen breakdown pathways increases. As determined in urine sediment and urine supernatant after digital rectal examination, Sreekumar et al. hypothesized that the prevalence of sarcosine increases with escalating severity of disease and this could lead us to an explanation to the aggressiveness of TNBC disease (24).
However, in 2011, Jentzmik et al. re-evaluated sarcosine in prostate cancer tissue samples and reported that sarcosine cannot be considered a suitable predictor of tumor aggressiveness or biochemical recurrence (56). The lack of metastatic tissue samples was their study limitation. Possible reasons for the contradictory results were discussed in two editorials (56, 57). But our data show that sarcosine was borderline correlated with death (3.84-fold survivor values, p=0.07). However, the small number of patients was a limitation of our study, hence we were unable to i) correlate the sarcosine concentration with the classic prognostic parameters of tumor grade, tumor stage and recurrence, and ii) estimate the potential of sarcosine as a predictor of tumor aggressiveness and the probability of cancer progression.
Table of all identified dipeptides and their correlation with death vs. survival. Biochemicals are shown with corresponding fold-change values and p-values for the specific comparison TNBC vs. ER. p≤0.05 was accepted as significant correlation with death vs survival; p-value trend of 0.05<p<0.10 identified biochemicals approaching significant correlation with death vs. survival. Platform: LC: Liquid chromatography; HMDB: Human Metabolome Database.
Tryptophan oxidation via the kynurenine pathway is an important mechanism of tumoral immunoresistance (58). Sakurai et al. evaluated the clinical significance of indoleamine 2,3-dioxygenase (IDO) expression in patients with breast cancer with bone metastasis (59). IDO activity was measured by the tryptophan/kynurenine ratio. They found that postoperative serum IDO levels of patients with breast cancer with a high number of bone metastases were lower than those of patients with a single metastasis. In addition, IDO activity increased in cases in which the number of metastatic lesions to the bone increased. These results suggest that expression of IDO in patients with breast cancer with bone metastasis may play a critical role in immunosuppression in these patients. It has also been suggested that interferon-gamma-mediated tryptophan breakdown acts to reduce substrate availability for NAD+ synthesis in cancer cells (60). In our study, TNBC samples contained elevated levels of tryptophan and its metabolic intermediates in NAD+ biosynthesis, kynurenine and quinolinate. TNBCs also had increased levels of the NAD+ salvage pathway biomarker nicotinamide. This may reflect the mechanism of immunoresistance of TNBCs and the role of kynurenine in TNBC progression and may be a potential marker of inflammation.
Table of all identified glutathione-related biochemicals, amino acids and their correlation with death vs. survival. Biochemicals are shown with corresponding fold-change values and p-values for the specific comparison TNBC vs. ER. p≤0.05 was accepted as significant correlation with death vs. survival; p-value trend of 0.05<p<0.10 identified biochemicals approaching significant correlation with death vs. survival. Platform: LC: Liquid chromatography; GC: Gas chromatography; HMDB: Human Metabolome Database.

Pathway diagram showing metabolic intermediates of the transmethylation pathway with related box plots. A: Pathway diagram showing the trans-methylation pathway and its affiliation with sarcosine (methylglycine). Metabolites with significant elevation in TNBC vs. ER+ are shown with solid arrows (p<0.05); trending elevations shown with open arrow (0.05<p<0.1). B: Relevant box plots showing multiple metabolites associated with the transmethylation pathway. p-Values are shown as insets for the comparison TNBC vs. ER+. SAH: S-Adenosyl-L-homocysteine. Box plots: +: Mean value; ---: Median value; o: Outliers; Top of box: Upper quartile; Bottom of box: Lower quartile; Top of whisker: Maximum of distribution; Bottom of whisker: Minimum of distribution. Sample cohorts are color coded: Light grey=ER+, Dark grey=TNBC. Data are shown as scaled intensity for each metabolite.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Pathway diagram showing metabolic intermediates of arginine metabolism with related box plots. A: Pathway diagram showing the several paths of arginine metabolism as they relate to nitric oxide synthesis, energy metabolism, the urea cycle, extracellular matrix (ECM) remodeling and proliferation. Metabolites with significant elevation in TNBC vs. ER+ (p<0.05) are shown with solid arrows. B: Relevant box plots showing multiple metabolites associated with arginine metabolism. p-Values are shown as insets for the comparison TNBT vs. ER+. NS: Not significant, p>0.1; ODC: Ornithine decarboxylase; iNOS: Inducible nitric oxide synthase; SAM-dc: S-Adenosyl methionine decarboxylated; NO: Nitric oxide; MTA: Methylthioadenosine. Box plots: +: Mean value; ---: Median value; o: Outliers; Top of box: Upper quartile; Bottom of box: Lower quartile; Top of whisker: Maximum of distribution; Bottom of whisker: Minimum of distribution. Sample cohorts are color coded: Light grey=ER+, Dark grey=TNBC. Data are shown as scaled intensity for each metabolite.
In our study, the intermediates of arginine incorporation into energy metabolism pathways were also elevated in TNBC samples. ODC, which facilitates the initial conversion of ornithine to putrescine in the polyamine synthesis pathway (61), has become a promising target for anticancer investigation and an independent prognostic factor for recurrence and death in patients with breast cancer (62). Activation of ODC and, consequently, increased concentrations of polyamines are associated with tumor promotion and progression (63). It has also been supposed that the ODC gene might act as an oncogene as its overexpression is essential for cell transformation (64); ODC activity has also been used as a biological marker for evaluating tumor growth and aggressiveness (65). Therefore, these changes may explain the mechanism that underlines the advanced/aggressiveness of TNBC.
Correlation of metabolites with clinical parameters such as cancer stage, ER and PR status contributes to the extent by which breast cancer can be detected early. Griskeodegard et al. showed a link between tumor metabolites and ER and PR status, with a predictive accuracy of 88% and 78%, respectively, indicating that the metabolic profile varies with the ER and PR status of the patient (66). Our data support these findings and inclusion of such parameters may help advance further development of early-detection metabolite profiles. Since we were unable to correlate the link between tissue sarcosine and clinical data, future studies will seek to validate the changes in sarcosine levels in different stages of breast cancer tissues and in sera, and estimate the potential of sarcosine as a predictor of tumor aggressiveness and the probability of cancer progression.
In conclusion, we identified many significant metabolic differences in the methanolic extracts of TNBC versus ER+ breast carcinomas that may directly impact the aggressive nature of TNBCs and the hypermethylation status of this tumor type, as previously shown by our studies and others. Through global biochemical profiling, we identified amino acid uptake and catabolism as having major potential influence on the aggressiveness of TNBCs versus ER+ tumors. Protein uptake and catabolism in the TNBC samples correlated with increased glutathione biosynthesis, NAD+ production, inflammatory and proliferative signaling, as well as potential increases in fumarate-mediated transcription factor hypoxia-inducible factor-1 changes: i.e. adaptation to hypoxia, angiogenesis, and metastasis. Additionally, increased uptake of methionine via the γ-glutamyl pathway correlated with increases in intermediates of the transmethylation pathway and the recently proposed oncometabolites sarcosine and 2-hydroxyglutarate. Although elevations of these oncometabolites have been reported separately for other cancer types, the results of this study may indicate novel biomarkers that delineate TNBCs from ER+ tumors in African-American women. The data also indicate Warburg metabolism in the TNBCs, as several biochemical intermediates of the glycolytic pathway were increased, including lactate. Elevated lactate and proline (and proline metabolites) also suggest elevated ECM remodeling in TNBCs compared to ER+ tumors. Finally, a robust utilization of arginine correlated with increased metabolites linked to nitric oxide-mediated inflammatory signaling, cell proliferation, energy metabolism and ECM remodeling. We recommend future studies on distinct breast cancer subtypes include non-cancerous tissues as standard control and for increased statistical power in order to separate metabolic differences and assess the effects of ER status on breast cancer more thoroughly, which could be quite interesting given our limited understanding of the role of ER loss in the racial health disparities associated with breast cancer. Furthermore, another limitation of this study is that it was performed using only African-American patients, and that future studies including a Caucasian arm may help elucidate potential mechanisms associated with racial differences related to this disease through the use of metabolomic profiling. To our knowledge this is the first metabolomics study that combines the information-rich analytical methods of GC, LC and MS to provide insights into understanding associated mechanisms and identifying pathways that are perturbed in TNBC and which therefore represent potential targets for new therapies of these aggressive carcinomas.
Disclosure
We have no personal or financial conflict of interest and have not entered into any agreement that could interfere with our access to the data on the research, or upon our ability to analyze the data independently, to prepare manuscripts, and to publish them.
Acknowledgments
This work was supported by Howard University College of Medicine Bridge Funds, and Pilot Study Awards Program (BFPSAP, U400042), and in part by the Howard University Cancer Center/Johns Hopkins Cancer Center Partnership grant (2U54 CA091431-06) funded grant by National Cancer Institute, National Institutes of Health.
- Received September 9, 2014.
- Revision received October 20, 2014.
- Accepted October 27, 2014.
- Copyright© 2014, International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved