

Abstract
Background/Aim: In this study we evaluated the proteomic profile of PC-3 cells treated with novel, 3-N-alkyloxyestradiol derivative, 3-[2-diisopropylamino]-ethoxy-D1,3,5 (10)-estrien-17-one (DI) (USPTO #7,687,486). Materials and Methods: The growth inhibitory potential of DI was determined by the National Cancer Institute (NCI) Developmental Therapeutics Program. 2-D gel electrophoresis and mass spectrometry were employed to identify differentially expressed proteins after treatment with DI. Results: Growth inhibitory (GI50) results showed that DI inhibited the growth of PC-3 and DU-145 cells, at 13.9 μM and 30.8 μM, respectively. Out of the proteins differentially expressed, five were selected for identification with four of those being successfully identified. The identified proteins play a role in protein folding, cell motility, carbohydrate biosynthesis, and carbohydrate degradation. Conclusion: Our studies resulted in the identification of targets associated with the glycolytic pathway and cell motility which have been implicated in the development and progression of many cancers.
- Prostate cancer
- SERMs
- selective estrogen receptor modulators
- NCI-60
- proteomics
- GRP78
- triosephosphate isomerase
- GAPDH
Over the last decade there have been huge strides in early detection methods and understanding of the molecular mechanisms fueling the development and progression of prostate cancer, specifically the castration and hormone-resistant forms. Although, these advances have led to the discovery of new molecular targets and therapeutic options, there still exist significant challenges regarding resistance to existing therapies, a need for new targets, more tolerable therapy options and a deeper understanding over existing molecular mechanisms and targets. According to Hanahan and Weinberg, there are very distinct hallmarks of cancer, all of which are regulated by complex signaling pathways and result from genomic instability (1). These hallmarks include characteristics that translate to the uncontrolled growth of cells and their ability to invade surrounding and distant tissues (1, 2).
3-[2-diisopropylamino]-ethoxy-D1,3,5 (10)-estrien-17-one (DI) is a novel selective estrogen receptor modulator that is a combination of an aminoalkyloxy group and the rigid structure of naturally-occurring, 17-β-estradiol, which is endogenous in the human body (3). DI been shown to have activity comparable to that of 4-hydroxytamoxifen (HO-Tam), the active metabolite of tamoxifen, in MCF-7 human breast cancer cells (3). DI has been screened by the National Cancer Institute (NCI) Therapeutics program against 60 human tumor cell lines regarding their anti-proliferative potential (See Figure 1 for DI/NCI-60 Screening Results). Due to our laboratory’s keen interest in prostate cancer, the present study focuses solely on DI-treated prostate cancer cells and their proteomic profile.
The present study used proteomics to investigate the underlying molecular targets associated with the growth inhibitory effects of the 3-N-alkyloxyestradiol (3) derivative, D1, in the androgen-independent PC-3 prostate cancer cell line. Further elucidation of underlying mechanisms will provide a better understanding over how DI modulates the expression of proteins and signaling networks that play a major role in prostate carcinogenesis.
Materials and Methods
Chemicals. The 3-N-alkyloxyestradiol derivative, 3-[2-diisopro-pylamino]-ethoxy-D1,3,5 (10)-estrien-17-one (DI) was synthesized and provided by Dr. John Cooperwood (Florida A&M University, Tallahassee, FL, USA). Propidium iodide, DMSO (dimethyl sulfoxide), D-Glucose and Ethanol was obtained from Sigma-Aldrich (St. Louis, MO, USA). RPMI 1640, penicillin-streptomycin antibiotic solution (10,000 UI/mL), 0.25% Trypsin-EDTA solution and phosphate buffer (PBS) were all obtained from Invitrogen/Life Technologies (Carlsbad, California, USA). Fetal bovine serum (FBS) was obtained from Atlanta Biologicals (Flowery Branch, GA, USA). Quick Start™ Bradford Reagent, 1X (BioRad, Hercules, CA) and Ready 2-D Starter Kit were obtained from BioRad (Hercules, CA, USA).
PC-3 cells were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum and penicillin-streptomycin antibiotic solution (10,000 UI/mL), at 37°C in a 5% CO2 humidified atmosphere. A stock solution was prepared by dissolving 3-[2-diisopropylamino]-ethoxy-D1,3,5 (10)-estrien-17-one (DI) in 100% DMSO (Sigma-Aldrich) with subsequent dilutions made in cell culture medium. In our experimental conditions, PC-3 cells were treated for 48 h with DI. Cells were detached with 0.25% Trypsin-EDTA solution washed and collected.
NCI developmental therapeutics program screening. DI was assayed at a single high concentration (10-5 M) in the full NCI 60 cell panel. Compounds that satisfy threshold inhibition criteria progress to the full five-concentration assay. The results are expressed as the percent growth (GI50) of treated cells at the test concentration of 10-5 M following 48 h incubation. The compound was dissolved in dimethyl sulfoxide (DMSO) and evaluated using five concentration of 10-fold dilutions (10-8-10-4 M) for 48 h.
Two-dimensional gel electrophoresis. Total proteins were extracted from control and treated groups by incubation with lysis buffer in -80°C. The protein pellets were re-solubilized in rehydration solution (7 M urea, 2 M Thiourea,4% CHAPS, 1% DTT, 1% Biolyte, 0.1% Protease Inhibitor Cocktail, HPLC-grade water, bromophenol blue) and kept at -80°C until further analysis. The total amount of proteins was determined using the Quick Start™ Bradford Reagent, 1X (Bio-Rad, Hercules, CA, USA) per the manufacturer’s instructions. The reagents provided in the Ready 2-D Starter Kit (Bio-Rad) were used to carry-out both dimensions. 300 μg of proteins were rehydrated into 11-cm immobilized ReadyStrip™ (Bio-Rad) IPG Strips (pH 3-10) overnight. The first dimension was run using the Protean IEF system (Bio-Rad) at room temperature with the following settings: step 1 at 250 V for 20 min; step 2 at 8000 V for 2.5 h; step 3 at 8000 V for up to 25,000 Vh. Upon completion of the first-dimensional separation, the strips were equilibrated using the Equilibration Buffers provided in the Ready Prep 2-D kit. The second-dimensional electrophoresis was performed running the samples on CRITERION TGX, Any kD, IPG+1 (pre-cast gels) using the Criterion System (Bio-Rad) in 1X Tris-Glycine-SDS running buffer with the following settings: step 1 at 200 V, constant for 1 h. After electrophoresis, the gels were washed three times at five min each. Then BioSafe Coomassie Stain was added and gels were allowed to stain overnight.
Differential Proteomic Evaluation: After staining, the gels were washed and scanned using the Versadoc Imager (Bio-Rad). PDQuest 2-D Analysis Software (Bio-Rad) was utilized for spot detection and matching. The quantity of each spot was normalized as a percentage of the total quantity of all spots in that gel. Statistical analysis involved the comparison of data from three repeated experiments using the Students t-Test. Only spots that showed significant differences to a level of 90% were selected for mass spectrometry analysis.
Protein Digestion, Desalting, MALDI-TOF/TOF. Protein spots were manually excised from the polyacrylamide gels and kept in sterile 1.5 mL Eppendorf tubes. Excised spots (gel plugs) were washed with destaining solution (100 mM ammonium bicarbonate) for 10 min, then dehydration solution (100% acetonitrile) for 10 min, liquid removed, then again with the dehydration solution, followed by discarding the liquid. This was repeated once. Then reduction solution (10 mM DTT) was added for 30 min followed by dehydration solution for 10 min, then alkylation solution (55 mM iodoacetamide with 100 mM ammonium bicarbonate) for 45 min. Trypsin (Protea Biosciences, West Virginia, USA) (10 ng/mL) was then added into gel plug and incubated overnight at 37°C. The following day 10% acetonitrile/0.2% trifluoroacetic acid was added to stop the digestion. The supernatant was transferred to a new tube and the gel plugs were re-extracted with 60% acetonitrile/0.1% trifluoroacetic acid. The supernatant was combined with the first extraction. Samples were dried using a speedvac and resuspended in 5% acetonitrile/0.1% trifluoroacetic acid and purified using Agilent C18 Tips (Bond Elux Omix) (Agilent, USA). The eluted proteins were mixed with MATRIX (Protea Bioscience, West Virginia, USA) solution and spotted on a MALDI plate using dry droplet method in triplicates. Samples were analyzed using ABI 4800 MALDI TOF/TOF Analyzer and generated peptides were blasted with MASCOT search algorithm to identify the possible proteins. The function of the identified proteins was determined using UniProtKB/Swiss-Prot protein database.
Results and Discussion
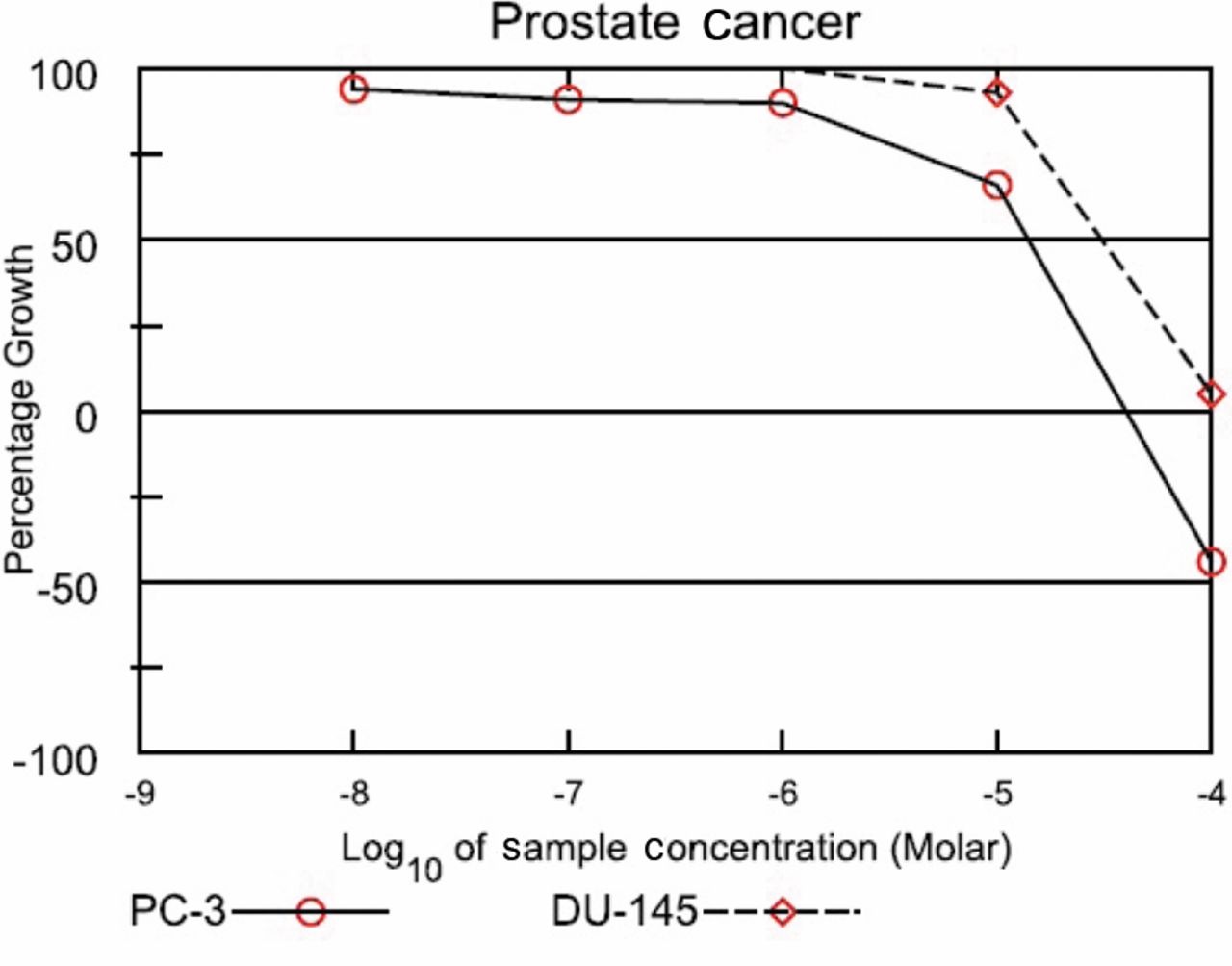
DI was screened by the NCI Developmental Therapeutics Program and growth inhibitory (GI50) results showed that the compounds inhibited the growth of two androgen-independent metastatic prostate cancer cell lines, PC-3 and DU-145, at 13.9 μM and 30.8 μM, respectively (Figure 2). 2-D gel electrophoresis was employed to identify proteins that were differentially expressed after treatment with the DI (Figure 3). Out of the proteins shown to be differentially expressed between treated and control groups, five were shown to be statistically significant, thus chosen for identification. Four of those chosen for identification were successfully identified and as described in the UniProtKB/Swiss-Prot protein database, play a role in protein folding, cell motility, carbohydrate biosynthesis, and carbohydrate degradation (Table I).
Glucose-regulated protein of 78 kDa (GRP78/Heat shock 70 kDa protein 5/BiP) is a protein that resides in the endoplasmic reticulum and plays a major role in polypeptide translocation, protein folding, and is a maker and initiator of endoplasmic reticulum stress, all of which are functions important to the viability of a cell (4, 5). It is up-regulated as a result of stress in the endoplasmic reticulum. GRP78 has been found to be up-regulated in breast tumor, hepatocellular carcinomas, gastric tumors, and oesophageal adenocarcinomas (6). In prostate cancer, its expression has been shown to be up-regulated as cells transition to metastatic castration-resistant forms (7). Intracellar GRP78 is associated with the pro-survival and anti-apoptotic characteristics of the protein, while the cell surface/extracellular GRP78 binds pro-apoptotic ligands. As described by Shrestha-Bhattarai and Rangnekar, endoplasmic reticulum stress results in the binding of PAR-4 to GRP78 and thus the activation of caspase 8 and 3 dependent apoptotic signaling and other downstream effector proteins (8). Further evidence of the involvement of GRP78 in apoptosis was shown in prostate cancer cells by Chiu et al. They reported that tanshinone IIA and n-butylidenephthalide increased expression of the GRP78/BiP, inhibited cell growth and induced apoptosis in prostate cancer cells (5, 9). Our results show that GRP78 was up-regulated in PC-3 cells in response to treatment with DI, providing evidence indicating that the possible mechanism driving its growth-inhibitory effects may be linked to apoptosis-dependent signaling.

National Cancer Institute Developmental Therapeutic Program Dose Response Curves for 3-[2-diisopropylamino]-ethoxy-D1,3,5 (10)-estrien-17-one (DI).
Actin cytoplasmic-1 (β-actin), commonly used as a housekeeping gene/protein, is involved in cell migration, cell division, embryonic development, wound healing and immune response, specifically providing protrusive forces that aid in the forward movement of migrating cells (10-15). Our results showed that the expression of β-actin increased in our samples as the concentration of DI increased. Research has shown that beta actin plays a role in cancer pathogenesis and has been found to be differentially expressed in many cancers, including prostate cancer (10, 15-18).
Triose phosphate isomerase (TPI), a glycolytic pathway enzyme, was shown to have decreased expression as a result of DI treatment, especially in the 25-μM treatment. This enzyme is essential in the process of energy production and glycolysis, where it catalyzes the inter-conversion of triose phosphate isomers dihydroxyacetone phosphate (DHAP) and D-glyceraldehyde-3-phosphate (GAP) (19). TPI has also been shown to be down-regulated in PC-3 treated with phyllanthus, which resulted in the suppression of proliferation and induction of apoptosis via signaling pathways including MAPKs, PI3K/Akt, NFκB, and Hypoxia (2).
Identified proteins differentially expressed in PC-3 cells after DI treatment.
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) is a glycolytic enzyme that is essential in glycolysis and energy production, breaking-down glucose for energy and carbon molecules. Recently it has been associated with such non-glycolytc roles transcription activation, cell death (initiation of apoptosis), cytoskeletal organization, endocytosis, exocytosis and DNA replication and repair (20, 21). Due to its key role in glycolysis, overexpression of GAPDH is commonly associated with cancer (22, 23). Studies have shown that GAPDH is differentially expressed in many cancers, including lung, renal, breast, prostate, liver, colorectal, bladder, pancreatic, gastric, melanoma and glioma (17, 21, 24-31). Our proteomic analysis results of SERM-treated PC-3 cells show a decrease in the expression of GAPDH with increasing SERM concentration. This suggests that SERM-induced growth inhibition in PC-3 cells could be mediated through the glycolytic or one of the above-described non-glycolytic mechanisms of GAPDH.
Conclusion
In the present study, the NCI screening of DI has shown its ability to inhibit the growth of DU-145 and PC-3 prostate cancer cells. We have employed 2-D gel electrophoresis to examine the proteomic profile of the PC-3 cells after treatment with DI. Our studies resulted in the identification of targets associated with the glycolytic pathway and cell motility. It appears that the 3-N-alkyloxyestradiol derivatives, specifically DI, are depleting cells of energy and affecting cell movement, both of which are essential for survival and progression of cancer cells. All identified proteins have been previously implicated in the carcinogenesis of many types of cancers, including prostate cancer. Taken together, these experimental studies provide compelling evidence of the growth inhibitory and possibly anti-metastatic properties of the 3-N-alkyloxyestradiol derivatives, thus endorsing this class of compounds worthy of further investigation.
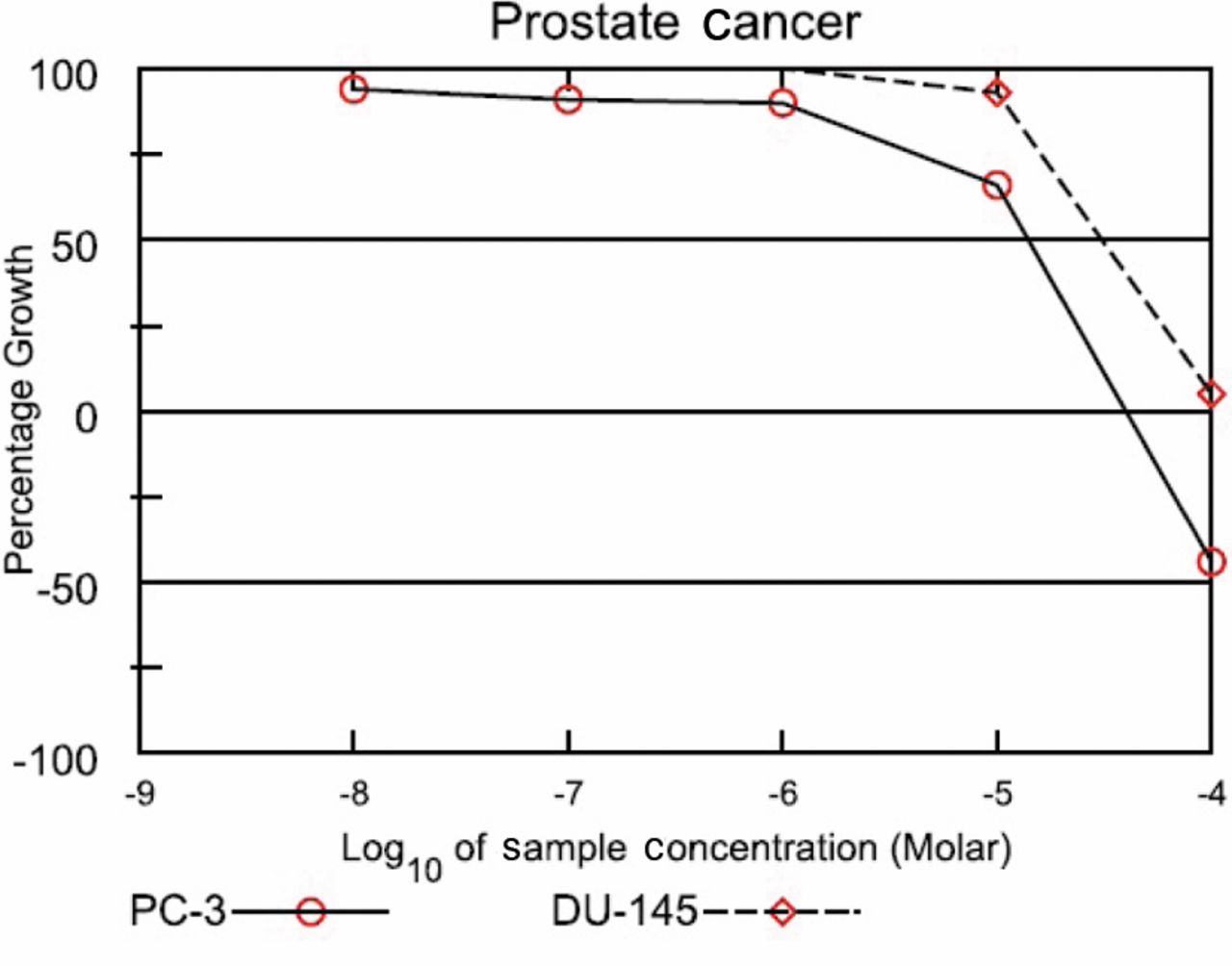
National Cancer Institute (NCI) Developmental Therapeutics Program Growth Inhibitory Curve. The compound, DI, was assayed at a single high concentration (10-5 M) in the full NCI 60 cell panel, results from the PC-3 and DU-145 cell lines are shown. Compounds that satisfy threshold inhibition criteria progress to the full five-concentration assay. The results are expressed as the percent growth (GI50) of treated cells at the test concentration of 10-5 M following 48 h of incubation (unpublished results). The compound was dissolved in dimethyl sulfoxide (and evaluated using five concentration of 10-fold dilutions (10-8-10-4 M) for 48 h.

{kind=link}
{kind=link}
{kind=link}
Representative 2-D gel images of PC-3 cells. PC-3 cells were treated with 10 μM, 25 μM and 40 μM of DI for 48 h followed by 2-DE analysis. Red circles indicate differentially expressed protein spots selected for mass spectrometry analysis. Selection was based on the following criteria: all spots that were statistically significant to a level of 90% (Student’s t-Test). Of the five protein spots of interest, we were able to identify four, using MALDI TOF/TOF mass spectrometry, in combination with MASCOT search algorithm.
Conflicts of Interest
The Authors declare that they have no financial or non-financial competing interests.
Acknowledgments
This research was supported by a grant from NIH NCRR RCMI program (G12RR 03020) and the National Institute on Minority Health and Health Disparities, NIH (8G12MD007582-28 and 1P20 MD006738-01).
- Received May 7, 2014.
- Revision received June 17, 2014.
- Accepted June 18, 2014.
- Copyright© 2014, International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved