


Abstract
Background: Cancer stem cells (CSCs) in gliomas are associated with resistance to radio- and chemotherapy, based on O6-methylguanine-DNA methyltransferase (MGMT) hypermethylation and the Multidrug resistance (MDR) system activation. Materials and Methods: Samples from 21 glioblastomas (GBMs) were put in culture with growth factors or serum in order to obtain neurospheres or adherent cells. Both were genetically and immunohistochemically characterized for ATP-binding cassette, sub-family B (MDR/TAP), member 1 (ABCB1), ATP-binding cassette, sub-family C (CFTR/MRP), member 1 (ABCC1) and MGMT expression together with primary tumors. Results: ABCB1 expression was positive in endothelial cells of primary tumors. ABCC1 expression was variably positive in tumor cells and positive in neurospheres, and less expressed in adherent cells. MGMT was methylated and unmethylated in primary tumors and in neurospheres, respectively, and unmethylated in adherent cells. Conclusion: Methylation is an epigenetic event affecting progenitors before the separation of the two glia lineages and maximally the future initiating cells. ABCB1 expression is limited to endothelial cells, whereas ABCC1 expression could mark a minority of tumor cells approaching a stem-like status.
Brain tumor stem cells (BTSCs) are characterized by proliferation capacity, renewal potential, clono- and tumorigenicity, and responsibility for tumor recurrence and resistance to therapies (1-4). They are also called glioma-initiating cells (5, 6) and are interpreted in the context of the known glioblastoma multiforme (GBM) heterogeneity. The existence of BTSCs is based on a hypothesis, which still provokes endless discussions. They are supposed to derive from the transformation of embryonic or adult neural stem cells (NSCs) but also from uni- or multipotent progenitors (7-9). They share stemness antigenic properties with NSCs and genetic alterations with primary tumors. An alternative hypothesis is that they derive from the de-differentiation of tumor cells (3, 10, 11), the opposite of the differentiation in normal cytogenesis that reaches a stem cell-like status (12). Therefore, the definition of BTSCs on the basis of a functional competence would be more appropriate than the one based on their being just a cell type (13). It must be pointed out that for other types of glioma, for example oligodendrogliomas, the origin from stem cells has not been contemplated (14-16).
The methodology of cell culture can influence the expression of BTSCs, which could even be regarded as a simple product of in vitro culture (17-18). There is an ongoing debae between researchers who rely upon neurosphere (NS) formation in culture conditions as expression of BTSCs, and those who obtain flat cells with the use of a coated matrix. This can be summarized by the expression “going round or going flat” (19-20). It is not just a semantic question, since it concerns the significance to be attributed to the cells in culture. In a previous article, in line with previous findings (6, 21), we observed that NS share genetic alterations with primary tumors typical of primary GBMs, whereas adherent cells (AC) do not, and on the contrary, exhibit differentiation antigens (12). AC could be interpreted as differentiating cells progressively losing their stemness properties (12).
As a typical characteristic, BTSCs exhibit resistance to chemo- and radiotherapy based on CD133-positive cells and on the activation of the multidrug resistance (MDR) system, hypermethylation of the O6-methylguanine-DNA methyl-transferase (MGMT) gene, poly [ADP-ribose] polymerase 1 (PARP1) and DNA repair (22-24). On this basis, we wanted to verify the behaviour of primary tumors, NS and AC in relation to the expression of genes/proteins of cell resistance such as ATP-binding cassette, sub-family B (MDR/TAP), member 1 (ABCB1), and ATP-binding cassette, sub-family C (CFTR/MRP), member 1 (ABCC1), and in relation to epigenetic events, such as promoter hypermethylation of MGMT.
Materials and Methods
Patients. The investigation was carried out on 21 primary hemispheric GBMs partially resected at the Department of Neuroscience, Neurosurgical Unit, University of Turin and at Maggiore della Carità University Hospital of Novara. The histological diagnosis of GBM was performed according to WHO guidelines (25). The mean age of the patients was 63.5 years (range=26-80 years). Twelve patients were males, and nine were females.
A viable sample from the dissected tumor was divided into three serial slices: one was stored at −80°C for molecular genetics and frozen sections, one was fixed in buffered-formalin and embedded in paraffin (FFPE), and one was washed, minced and enzymatically dissociated for expansion in culture. A peripheral blood sample was obtained from each patient and stored at −20°C. All patients provided informed consent allowing for the use of tumor specimens for molecular and cell culture studies. The study was approved by the Ethics Committees of the University Hospital of Turin and Novara.
Histology and immunohistochemistry (IHC). Hematoxylin and eosin (H&E) staining was performed on 4-μm paraffin sections.
IHC was performed using a Ventana Full BenchMark® automatic immunostainer (Ventana Medical Systems Inc., Tucson, AZ, USA). UltraView™ Universal DAB Detection Kit was used as detection system. The antibodies used were mouse anti-ABCB1 (AB4336, 1:200; Chemicon International Inc., Temecula, CA, USA) and mouse anti-MRP1 (MAB4147, 1:150; Chemicon). Heat-induced epitope retrieval (HIER) was performed in Tris-EDTA, pH 8 (Ventana Medical Systems, Tucson, AZ, USA). Negative controls were obtained by omitting the primary antibody.
The analysis of MGMT protein expression was performed by a conventional streptavidin-biotin protocol as previously described (26) using a mouse anti-MGMT antibody (MAB16200, 1:100; Chemicon). The staining was evaluated on the basis of its distribution, intensity and percentage of positive cells, according to a previous scheme (26).
In vitro cultures. Tumor tissue was processed as previously described (27). Culture conditions were the following: for NS, Dulbecco's modified Eagle's medium (DMEM)/F-12 with 10 ng/ml basic fibroblast growth factor (bFGF) and 20 ng/ml epidermal growth factor (EGF); for AC, DMEM with 10% fetal bovine serum (FBS). Both cultures were maintained at 5% O2/CO2. GBM reference cell lines were U87-MG and 010627 (Galli R, DIBIT San Raffaele, Milan, Italy) for both AC and NS.
All cell cultures were periodically checked for Mycoplasma contamination (e-Myco™ Mycoplasma PCR Detection kit; iNtRON Biotechnology, Gyonggi, Korea).
Differentiation assays were performed by mitogen withdrawal and the addition of 3% FBS to the culture. Clonogenicity assay and growth rates were performed for both NS and AC as already reported (12).
Immunofluorescence. Immunofluorescence staining was performed on NS and AC. Goat anti-rabbit fluorescein isothiocyanate (FITC)-conjugated IgG and rabbit anti-mouse tetramethyl rhodamine isothiocyanate (TRITC)-conjugated IgG antibodies were used. Cell nuclei were stained with 4’,6-diamidino-2-phenylindole (DAPI). Primary antibodies were the same used for immunohistochemical analysis. Observations were made on a Zeiss Axioskop fluorescence microscope (Karl Zeiss, Oberkochen, Germany) equipped with an AxioCam5MR5c and coupled to an Imaging system (AxioVision Release 4.5; Karl Zeiss).
DNA extraction. Genomic DNA (gDNA) was extracted from frozen tumor samples according to a standard phenol-chloroform protocol. Before DNA extraction for each sample, only tumor areas previously identified as proliferating by H&E staining and microscopic examination were selected. gDNA from cell lines and peripheral blood was isolated using the QIAmp DNAMini Kit (Qiagen, Hamburg, Germany) and a salting-out procedure, respectively. The genetic analysis described below was carried out on primary tumors, NS and AC.
MGMT promoter hypermethylation status. The promoter hypermethylation status of the MGMT gene (GenBank sequence NM_002412) was determined by methylation specific polymerase chain reaction (MS-PCR) followed by capillary electrophoresis as previously reported (26). Sodium bisulphite modification was performed with the MethylEasy™ Exceed Rapid DNA Bisulphite Modification Kit (Human Genetic Signatures Pty Ltd., Macquarie Park, Sydney, Australia) (26). CpGenome™ Universal Methylated DNA (Chemicon) and normal lymphocyte DNA were used as methylated and unmethylated controls, respectively. The primer sequences for MS-PCR and the amplification conditions have already been reported (28). After electrophoresis on an ABI® 3130 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA), data were collected using GeneMapper v4.0 software for fragment analysis (Applied Biosystems). The MS-PCR products for the methylated and unmethylated allele corresponded to a 81 and 93-base pair peak, respectively. A height ratio >0.1 between peaks for methylated and unmethylated allele was scored as evidence of the methylated status of the MGMT gene (mean of two replicates) (26).
Western blotting analysis. Frozen tissue and cultured cells (both NS and AC) were homogenized in a lysis buffer supplemented with Protease and Phosphatase Inhibitor Cocktails (Sigma Aldrich Co., St. Louis, MO, USA). Tissues were sonicated with three 10-s bursts. Whole protein extracts were quantified by BCA™ Protein Assay Kit (Pierce Biotechnology, Rockford, IL, USA), equal amounts were resolved in 10% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) gels and transferred onto polyvinylidene difluoride (PVDF) membranes (Immobilon-P; Millipore, Bedford, MA, USA). Blots were probed with a mouse anti-MGMT antibody (MS-470-P0; NeoMarkers, Fremont, CA, USA) and with the same antibodies used in immunohistochemistry for ABCB1 and ABCC1 analysis. Membranes were then incubated with the appropriate horseradish peroxidase (HRP)-conjugated secondary antibodies and proteins were detected by enhanced chemiluminescence (Millipore). An anti-α-tubulin antibody was used to normalize sample loading and transfer. Band intensity was quantified by densitometry using NIH Image J software (RSB, NIMH, Bethesda, MD, USA).
Results
Histology. Slices from the 21 tumor samples used for histological analyses were serial to those used for cell culture. The following phenotypic features characterized 7/10 (70%) samples generated by NS and 3/13 (23%) samples generated by AC: high cell density, high number of mitoses, a Ki-67/MIB-1 labeling index (LI) between 25% and 30%, circumscribed necroses with pseudo-palisading, high vessel density, and perivascular cuffings of tumor cells.
Immunohistochemistry of primary tumors. Positive staining for MGMT was evaluated according to the already published scheme (26, 29), and was observed in 11/21 cases (52.3%) (Figure 1a, c). ABCB1 was positive in endothelial cells but negative in tumor cells (Figure 2a). ABCC1 exhibited positive variable cytoplasmic immunostaining in tumor cells (Figure 2c) in all cases; sometimes it was also positive in endothelial cells.
Immunofluorescence. In NS, MGMT was either expressed or not (Figure 1b, d). NS and AC were negative for ABCB1 expression (Figure 2b). In contrast, ABCC1 expression was positive in all the NS (Figure 2d) and weakly positive in AC, except for two cases.
In vitro culture assay. NS were obtained in 10 cases (CV1, CV7, CV10, CV13, CV17, CV20, CV21, NO3, NO4 and NO6) with DMEM/F-12 with bFGF and EGF. AC were obtained in 13 cases (CV1, CV2, CV3, CV4, CV6, CV8, CV9, CV10, CV17, CV21, NO2, NO3 and NO6) with DMEM and FBS. Four cases (CV5, CV11, CV15 and CV16) did not produce any cells in culture. In one case (CV1), AC were developed together with NS in DMEM/F-12 with bFGF and EGF, and in one case (CV10), NS grew from AC when transferred to a low-adherent culture plate with DMEM/F-12 medium and mitogens. Four cases (CV17, CV21, NO3 and NO4) generated both NS and AC in their respective medium conditions.
NS showed clonogenicity, differentiation upon serum addition and variable growth rates, as already demonstrated (12).
Molecular genetics. The match of NS and AC with the respective primary tumors was verified by genotyping of 13 microsatellite markers, as reported elsewhere (12).
By MS-PCR, the MGMT hypermethylation status was successfully determined in all primary tumors and in the matched cells. MGMT promoter hypermethylation was identified in 10/21 (47.6%) primary tumors (CV2, CV4, CV6, CV10, CV11, CV15, CV16, CV17, CV20, NO4). Among primary tumors generating either NS or AC, 7/17 (41.2%) cases (CV2, CV4, CV6, CV10, CV17, CV20, NO4) exhibited MGMT hypermethylation.
MGMT molecular genetics and immunohistochemistry.
Among these primary tumors with hypermethylation, 4/7 (57.1%) (CV10, CV17, CV20 and NO4) gave rise to hypermethylated NS and 3/7 (42.6%) (CV2, CV4, CV6) to AC. All AC were unmethylated, with the exception of cells CV10, U87-MG and 010627 which were completely hypermethylated. There were 11/21 (52.4%) unmethylated primary tumors: four generated unmethylated NS (CV1, CV13, NO3, NO6), seven unmethylated AC (CV1, CV3, CV8, CV9, CV21, NO2, NO3) and two generated hypermethylated NS (CV7, CV21).
Among NS, MGMT promoter was hypermethylated in 6/10 (60%) cases (CV7, CV10, CV17, CV20, CV21, NO4). Hypermethylated NS originated from hypermethylated primary tumors with the exception of two cases (CV7 and CV21) (Table I). Four out of six NS (CV10, CV20, CV21, NO4) (66.7%) were homozygously hypermethylated, whereas two cases (CV7 and CV17) had both MGMT hypermethylated and unmethylated alleles (Figure 3). NS of reference cell lines, U87-MG and 010627, both carried hypermethylated MGMT.
In contrast to NS, in MS-PCR each hypermethylated and matched primary tumor always displayed evidence of a signal for the MGMT unmethylated allele as a consequence of the presence of unmethylated tumor and normal cells in the sample.

O6-methylguanine-DNA methyltransferase (MGMT) expression in nuclei of primary tumors by immunohistochemistry, ×200 (a). MGMT expression in neurospheres by immunofluorescence, ×400 (b). Nuclei of MGMT-methylated primary tumors negative by immunohistochemistry, ×200 (c). Neurospheres negative by immunofluorescence, 4’,6-diamidino-2-phenylindole (DAPI), ×400 (d).
Western blotting analysis. MGMT expression in cell lines was consistent with the MGMT promoter hypermethylation status. Because of contamination, a band for MGMT was variably found in all the primary tumors. In NS, the band was not found in two cases (CV10 and CV20). In two cases (CV7 and CV17), a small band was found and this corresponded to the finding of both methylated and unmethylated MGMT alleles by MS-PCR. A band was constantly found in all cases of AC (Figure 4a).
ABCB1 was never found in NS nor AC (Figure 4b). ABCC1 exhibited a positive band in NS and was lacking in AC (Figure 4c).
Discussion
MGMT promoter hypermethylation does not correlate with malignancy and it is more frequent in low-than in high-grade gliomas (29-37). In GBMs, it occurs in 32-72% of cases (26, 30, 31, 38, 39), but it is more frequent in secondary GBMs (sGBMs) than in primary GBMs (pGBMs) (29, 32, 33, 40-42); in our series, the frequency was 75% (29) in sGBMs and 25% and 31.5% in anaplastic astrocytomas and pGBMs, respectively.
In the present series of GBMs, which is composed of pGBMs only, the percentage of MGMT hypermethylation in primary tumors was 47.6%, whereas that in NS was 60%. It has been demonstrated that in primary tumors, MGMT methylation ranges from 10 to 90% and in NS is 100%; both the frequency and methylation density are greater in NS than in primary tumors, which are associated with no or low MGMT expression (43). In our material, methylated NS derived from methylated tumors and unmethylated NS derived from unmethylated tumors. Only in two cases did methylated NS derive from an unmethylated primary tumor; probably, in the latter, the methylated MGMT allele was undetectable. This means that hypermethylation is not linked to cell stemness. Since NS, which correspond to glioma-initiating cells, exhibit a high methylation density with both methylated alleles and since hypermethylation occurs both in astrocytomas and oligodendrogliomas (43), it can be deduced first that hypermethylation is an early epigenetic event (44) affecting progenitors before the separation of the two lineages, and second that hypermethylation maximally affects the future initiating cells.

ABCB1 expression in endothelial cells by immunohistochemistry, ×200 (a). No ABCB1 expression in neurospheres by immunofluorescence, DAPI, ×400 (b). MRP1 expression in tumor cells by immunohistochemistry, ×200 (c). ABCC1 expression in neurospheres by immunofluorescence, ×400 (d).
According to a widely accepted hypothesis, in glioma development, tumor-initiating cells differentiate, losing stemness properties (1), and the number of MGMT methylated and unmethylated tumors does not vary between low- and high-grade gliomas. The percentage of unmethylated cells in each tumor group is roughly similar, with slightly higher values in GBM due to contamination (29). This way, the epigenetic hypermethylating event should affect progenitors independently of their fate and the rapidity of their transformation (45). Consequently, NS behave like primary tumors. The higher percentage of methylated sGBMs in comparison with pGBM may be due to the more rapid transformation of the latter.
In AC, non-methylation was more prevalent, inconsistent with the finding on NS, but consistent with the prevalence of unmethylated primary tumors among those generating AC. It could be a stochastic finding associated with the low number of cases, or may be attributed to the generation of AC from a tissue slice contiguous but not identical to the one that underwent MS-PCR.
Even if stemness was considered as a functional status (13, 45), proliferation is one of its features and, therefore, it is not astonishing that a hypermethylating event takes place more frequently in proliferating cells, i.e. progenitors which maintain at least partially stemness properties. Our findings could be consistent with the observation that CD133-positive stem cells exhibit higher levels of MGMT mRNA, which activates DNA repair (46). As a matter of fact, the activation of ataxia-telangiectasia-mutated (ATM) and Checkpoint 1/Checkpoint 2 (Chk1/Chk2) and then DNA repair is another property of CD133-positive cells (47).
As for the MDR system, one clear-cut result is that ABCB1 was not expressed in tumor cells nor in NS and AC. This is in agreement with the occurrence of ABCB1 in the endothelial cells (48, 49), but not with the literature observations on gliomas, which showed that it was active in tumor cells (50, 51), even though much less than ABCC1 (52).

Analyses of the O6-methylguanine-DNA methyltransferase (MGMT) gene by methylation specific polymerase chain reaction (MS-PCR) and capillary electrophoresis in primary tumors of glioblastoma multiforme and respective neurospheres (NS) and adherent cells (AC). Case NO3: MGMT-unmethylated primary tumor, NS and AC; case NO4: MGMT-hypermethylated primary tumor and NS, MGMT-unmethylated AC; case CV17: MGMT-hypermethylated primary tumor and NS, MGM-unmethylated AC. The 81-base pair peak refers to the methylated allele and the 93-base pair peak to the unmethyalted allele. Arrow indicates the GeneScan™ 500 LIZ™ Size Standard (Applied Biosystems).
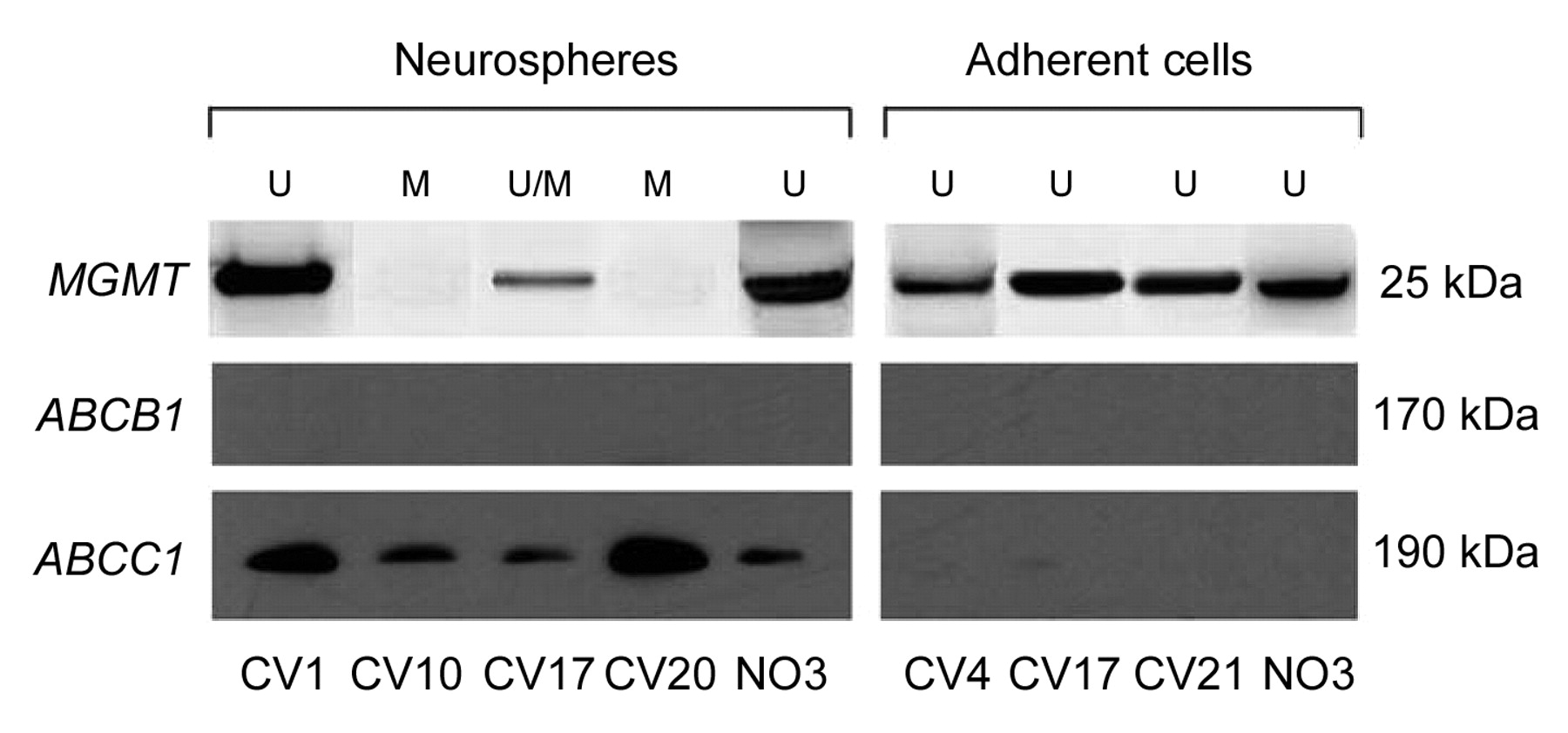
Western blotting analysis for O6-methylguanine-DNA methyltransferase (MGMT), ABCB1 and ABCC1 expression in neurospheres and adherent cells. U, Unmethylated MGMT promoter; M, methylated MGMT promoter.
ABCC1 expression in NS is quite in line with their resistance to radio- and chemotherapy as a feature of stemness, consistent with the above mentioned observations (46, 47). In primary tumors, the number of ABCC1 positive cells is variable and could represent a minority of cells that are resistant, probably because they are near a stem-like status. As a matter of fact, in AC ABCC1 expression decreases (53-55).
Acknowledgements
This work was supported by a Grant from Compagnia di San Paolo, Turin.
- Received April 12, 2012.
- Revision received June 29, 2012.
- Accepted June 29, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.