


Abstract
Background: 20-Hydroxyeicosatetraenoic acid (20-HETE), a metabolite of arachidonic acid (AA) produced by the CYP4A and CYP4F enzyme families has been reported to induce mitogenic and angiogenic responses both in vitro and in vivo, and inhibitors of this pathway reduced growth of brain and kidney tumors. Materials and Methods: Real-Time PCR, western blot and immunohistochemistry were used to compare the expression of CYP4A/F mRNA and protein levels in human cancer tissue samples versus normal controls. Liquid chromatography/mass spectrometry analysis (LC-MS/MS) was performed to measure 20-HETE formation in tumor homogenates. Activation of Ras in human proximal tubule epithelial cells (HRPTEC) treated with stable agonist of 20-HETE was measured using a Ras pull-down detection kit. Results: The expression of CYP4A/4F genes was markedly elevated in thyroid, breast, colon, and ovarian cancer samples in comparison to matched normal tissues. Furthermore, the levels of the CYP4F2 protein and of 20-HETE were higher in ovarian cancer samples compared to normal control tissues. A stable 20-HETE agonist induced activation of the small-GTPase Ras in HRPTEC cells. Conclusion: The present finding of elevated expression of CYP4A/F enzymes in human cancer tissue suggests that 20-HETE inhibitors and antagonists may be useful in the treatment of cancer.
Lipid mediators, collectively called eicosanoids, are derived from the metabolism of arachidonic acid (AA). The three major classes of eicosanoids are prostanoids, products of cyclooxygenases, 5, 12, 15-HETEs and leukotrienes products of lipoxygenases, and EETs and HETEs products of CYP mono-oxygenases. Collectively, eicosanoids are hormone-like messengers that can signal in an autocrine and a paracrine fashion resulting in mitogenic and anti-mitogenic actions. Mitogenic signaling of these lipid mediators confers cell growth, invasion, angiogenesis, resulting in enhanced resistance of cancer cells to chemotherapeutic agents (1, 2). Therefore, to increase chemotherapy sensitivity in cancer cells, combination therapy strategies are needed that will target both pro-survival pathways and simultaneously inhibit overproduction of eicosanoids in tumor cells. Recent studies have implicated HETEs, notably 20-HETE (20-hydroxyeicosatetraenoic acid) in cell growth and cancer development. The tumor-promoting effects of 20-HETE catalyzed by ω-hydroxylation of AA by the CYP4A/F enzymes has been documented in numerous studies (3-5). 20-HETE induced cell proliferation and DNA synthesis in vascular smooth muscle and proximal tubule cell cultures as well as angiogenic effects by stimulating endothelial cell proliferation, which was accompanied by increased vascular endothelial growth factor (VEGF) expression and release (3, 5, 8). More recent studies by Guo et al. reported that HETE0016, a selective inhibitor of CYP4A/F enzymes (9, 10) inhibited the formation of 20-HETE resulting in suppression of both 9L gliosarcoma and U251 glioma cell proliferation and tumor growth (6, 11). Similarly, the same group demonstrated that overexpression of the 20-HETE synthase CYP4A1 into U251 glioma cells resulted in hyperproliferation and more aggressive tumor growth (12). Additionally, inhibition of the synthesis and actions of 20-HETE with HET0016 and WIT002 (20-HETE antagonist) inhibits the proliferation of renal adenocarcinoma cells in vitro and the growth of kidney cancer tumors in an animal model in vivo (7). However, very little has been done to determine if the expression of CYP4A/F enzymes at either the mRNA or protein levels is elevated in human cancer samples.
Materials and Methods
Cell culture. Human renal proximal tubule epithelial cells (HRPTEC), purchased from Cambrex Bio Science Inc. (Walkersville, MD, USA), were grown in renal epithelial cell basal medium (REBM) supplemented with 0.5% fetal bovine serum, 0.1% mEGF, 0.1% insulin, 0.1% hydrocortisone, 0.1% GA-1000, 0.1% epinephrine, 0.1% T3 and 0.1% transferrin (all obtained from Lonza, Walkersville, MD, USA). The cells were maintained at 37°C in a humidified incubator containing 5% CO2.
Drugs. A stable 20-HETE agonist 5(Z),14(Z)-20-HEDE (WIT003) was synthesized by Dr. John R Falck (University of Texas Southwestern). Indomethacin (a nonselective cyclooxygenase (COX) inhibitor) was purchased from Sigma-Aldrich (St. Louis, MO, USA), arachidonic acid and NADPH were from Cayman chemicals (Ann Arbor, MI, USA), and EGF was from BD Biosciences (San Jose, CA, USA).
Real-Time PCR. TissueScan oncology survey cDNA panels were obtained from OriGene (Rockville, MD, USA) and quantitative PCR analysis was performed using iQ SYBR Green (BioRad Laboratories, Hercules, CA, USA) and Mx3000P real-time PCR system (Stratagene, Santa Clara, CA, USA). 20-HETE producing CYP450 isoform specific primer sequences were as follows: CYP4A11-F (ATGAAGTGTGCCTTCAGCCA), CYP4A11-R (AAG GCATTCCTCACACGGG), CYP4A22-F (AATGGGAAGAGCTCCTTGGC), CYP4A22-R (AAGGCATTCCTCATACAGC), CYP4F2-F (AAGCACCCAGAATACCAGGA), CYP4F2-R (TCATGCACATGGTCAGGAAG), CYP4F3-F(CTGTCGGCAGGAGGTACAAG), CYP4F3-R (CCTCAGGCTCTCCTTAATGC). PCR reactions contained 2-3 ng cDNA per well, based on normalization to β-actin and 0.1 μM final primer concentration in 25 μl. Reaction cycling parameters were as follows: 1 cycle at 95°C for 3 min, 45 cycles at 95°C for 10 s, 58°C for 45 s. The relative expression of the CYP isoforms was compared using the delta CT method.
Western-blot. The OncoPair Insta-Blot ready to use PVDF western blot membrane containing denatured protein lysates from diseased and adjacent normal tissue was obtained from Imgenex (San Diego, CA, USA). The Insta-Blot PVDF membrane was wetted with 100% methanol to rehydrate, then blocked in 5% nonfat dry milk for 1 h and incubated with an isoform-specific CYP4F2 primary antibody (HPA014048, Sigma Aldrich) overnight at 4°C. Following incubation with the primary antibodies, membranes were immunoblotted with goat anti-rabbit-HRP conjugated secondary antibody (BioRad) incubated with ECL+ (Amersham, Pittsburg, PA, USA) and exposed to film to detect chemilluminescence signal.
Immunohistochemistry. Formalin-fixed paraffin-embedded immunohistochemistry slides for adenocarcinoma of ovary (papillary serous) and normal ovary were obtained from Cytomyx (Division of Origene Technologies, Rockville, MD, USA). Slides were stained with a CYP4F2 primary antibody (dilution 1:25) and further incubated with goat anti-rabbit-HRP conjugated secondary antibody. DAB (3,3’-diaminobenzidine) was used as chromogen and sample slides were further counterstained with hematoxylin and eosin (Dako-Cytomation, Carpinteria, CA, USA).
CYP4A/F metabolism of AA in cancer tissue. Frozen human ovarian cancer and normal tissues (500 mg) were obtained from Proteogenex (Culver City, CA, USA). The frozen tissue was ground to a fine powder using a liquid nitrogen-cooled mortar (Fisher, Pittsburg, PA, USA) and homogenized in a buffer containing 250 mM sucrose, 1 mM EDTA, 1 mM KH2PO4, 9 mM K2HPO4 and 0.1 mM PMSF by a mechanical homogenizer, followed by sonication. The nuclear and mitochondrial fractions were removed by centrifugation and 1 mg/ml of the lysate was incubated for 1 h at 37°C in the presence of 100% O2 with exogenous AA (40 μM), 1 mM NADPH and 2 μM indomethacin (to prevent metabolism of 20-HETE by COX). The reaction was stopped by acidification with formic acid to pH 3.5 and eicosanoids were extracted with ethyl acetate after being spiked with 2 ng of 20-HETE-d6 internal standard. The organic phase was dried under nitrogen gas. The eicosanoids were separated by HPLC on a Betabasic C18 column (150×2.1 mm, 3 μM; Thermo Hypersil-Keystone, Bellefonte, PA) at a flow rate of 0.2 ml/min using an isocratic elution with 51:9:40:0.01 mixture of acetonitrile: methanol:water:acetic acid for 30 min followed by a step gradient to 68:13:19:0.01 acetonitrile: methanol:water:acetic acid for 15 min. The effluent was ionized using negative-ion electrospray and peaks eluting with a mass to charge ratio (m/z) of 319>245 (20-HETE), 319>301 (HETEs and EETs), 337>319 (DiHETEs), or 325>251 (internal standard) were monitored using an Applied Biosystems API 3000 LC–MS/MS. The area ratio of ion abundance in the peaks of interest to internal standards were compared with standard curves generated over a range from 0.2 to 10 ng for 20-HETE and from 1.0 to 10 ng for other metabolites in order to quantify the production of 20-HETE and other eicosanoids.
Active-Ras GTPase pull-down assay. HRPTEC cells were plated in 100-mm dishes and were grown in renal epithelial basal (REBM) media supplemented with growth factors up to 80% confluence. Prior to experiment, the cells were serum-starved in REBM basal medium containing no growth factors for 24 h. The cells were stimulated with 20 μM 5(Z),14(Z)-20-HEDE (WIT003); a stable 20-HETE agonist (13) and were immediately lysed in a buffer containing protease inhibitors. 900 μg protein was taken for an active Ras pull-down assay (Thermo Scientific, Logan, UT, USA). To pull-down active Ras, cell lysates were incubated with a GST fusion protein of the Ras effector protein binding domain (RBD) of Raf1, immobilized on gluthione agarose resin. Non-specific proteins were washed away, samples were eluted by 2× SDS buffer, and then separated by 4-20% SDS–PAGE, transferred to a PVDF membrane and probed for active Ras with anti-Ras antibody at 4°C overnight. Goat anti-mouse–HRP conjugated antibody (BioRad) was used as a secondary antibody and the detection was carried out with ECL+ chemiluminescent substrate, followed by exposure to X-ray film.
Results
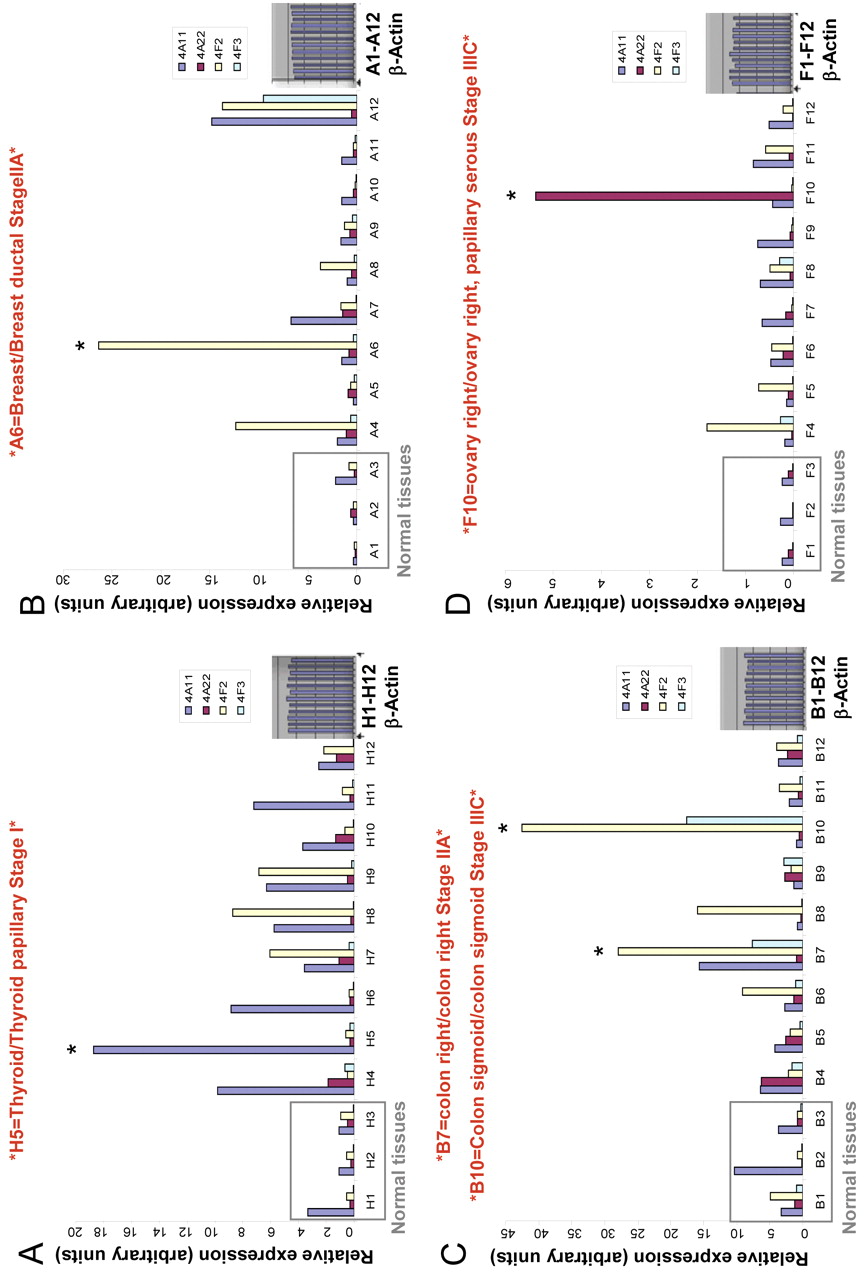
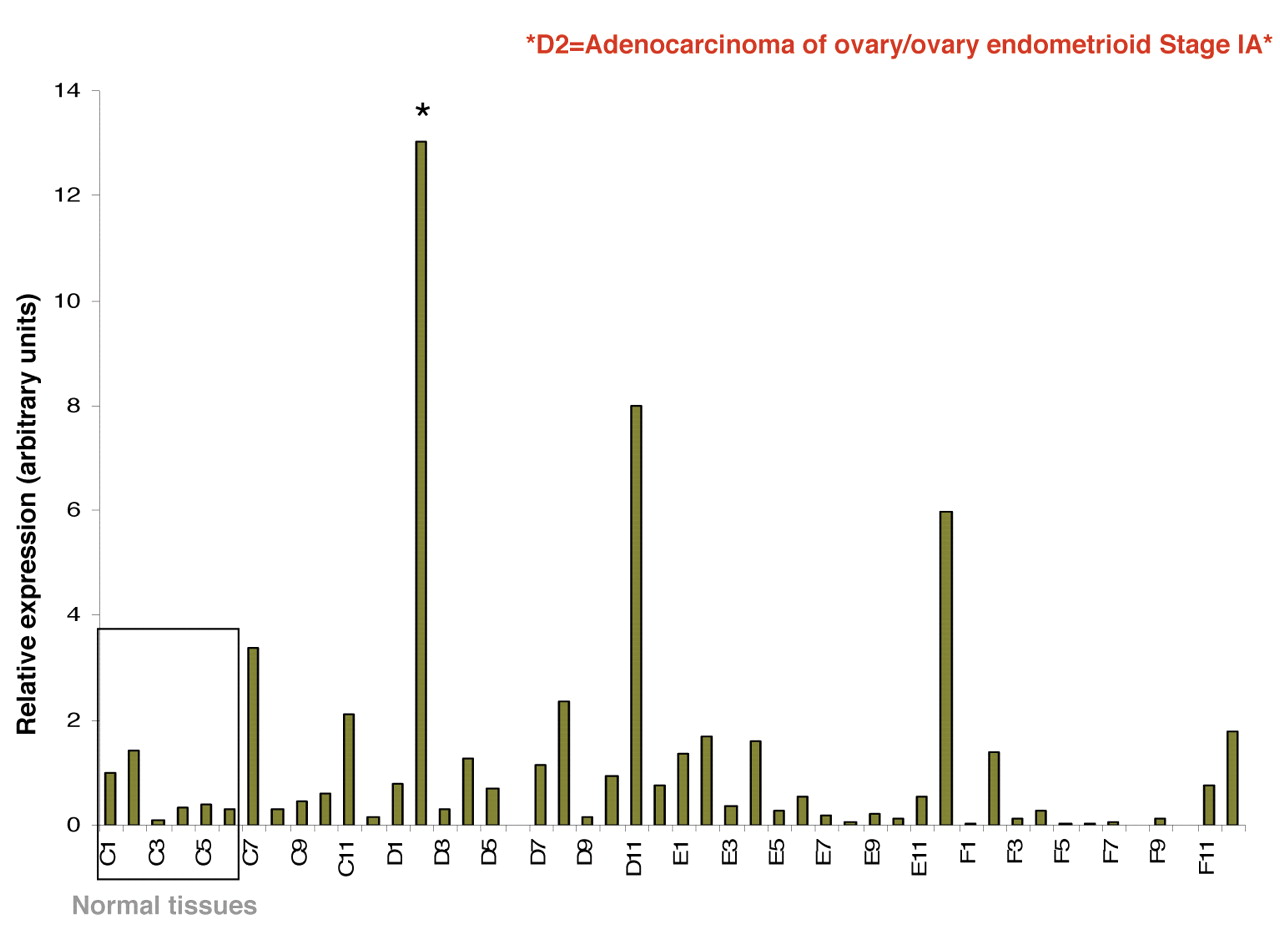
Expression of 20-HETE-producing CYP4A and CYP4F enzymes at mRNA level is up-regulated in human tumor tissue samples. We examined the presence of mRNAs encoding 4 human CYP450 isoforms that mediate 20-HETE production: CYP4A11, CYP4A22, CYP4F2 and CYP4F3 in eight different human cancer tissue samples (breast, colon, kidney, liver, lung, ovary, prostate and thyroid). As detected by real-time PCR (Figure 1 panels A, B, C, D), certain cancer tissues such as thyroid, breast, colon and ovary demonstrated striking overexpression of CYP4A and CYP4F genes at the mRNA level when compared with matched normal tissue. Expression of the CYP4F2 gene was further studied in panels representing different stages of ovarian cancer. A representative of two experiments is reported demonstrating up-regulated CYP4F2 expression (Figure 2).
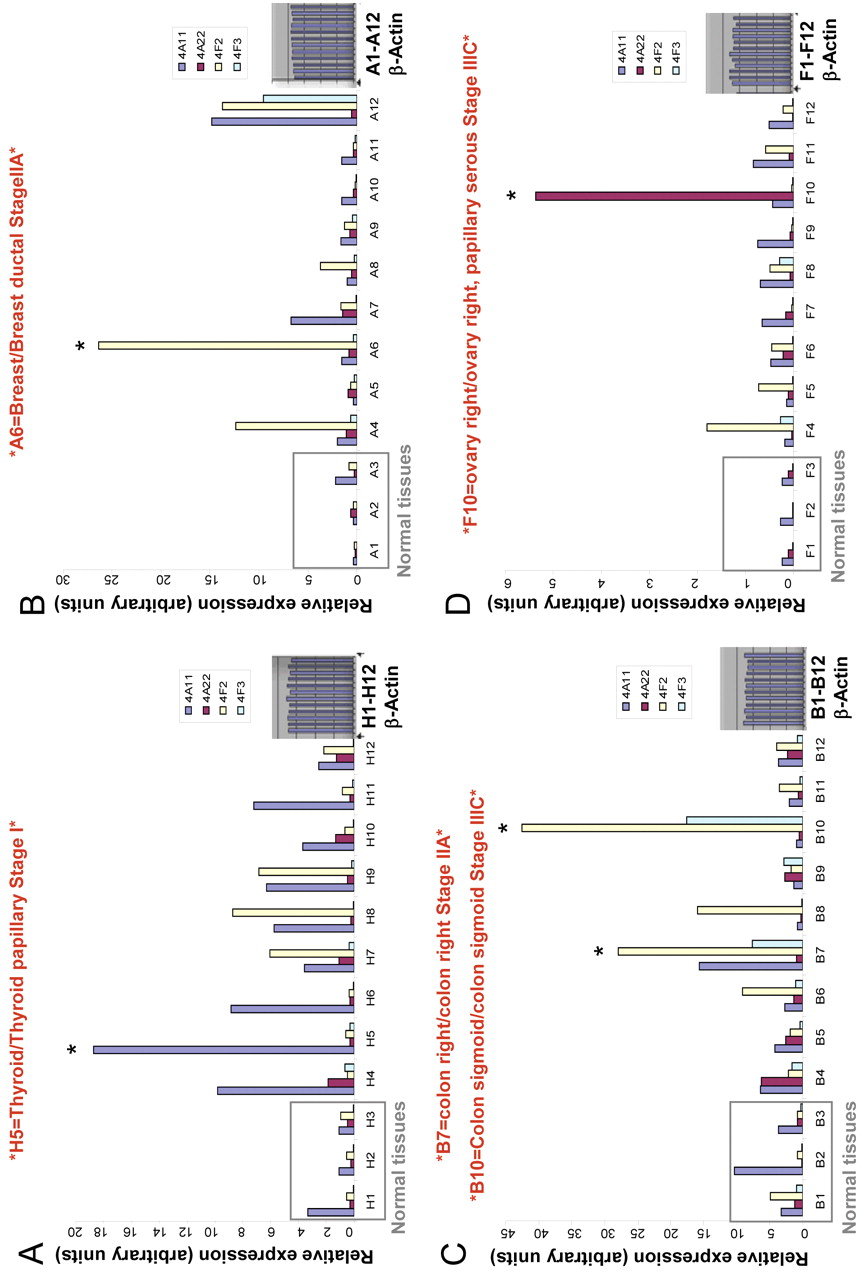
Expression of CYP4A and CYP4F genes at mRNA level is elevated in human cancer tissue samples. TissueScan cancer survey panels containing pre-normalized cDNA from multiple cancer tissues. (A) Thyroid; (B) breast; (C) colon; (D) ovary, were used to run real-time PCR reaction to analyze the expression of CYP4A and CYP4F genes at the mRNA level. The tissue samples used for each type of TissueScan panel, obtained from independent patients, cover various disease stages, and matched normal control tissues are included for comparison between normal and disease samples. The amount of PCR product is correlated with the amount of original RNA transcript and data the expressed in arbitrary units. *indicates tumor samples demonstrating striking up-regulation in CYP4A/F enzymes.

Expression of CYP4F2 gene is elevated in ovarian cancer tissue samples. TissueScan cancer survey panel containing pre-normalized cDNA from ovarian cancer tissues, was used to run real-time PCR reaction to analyze the expression of CYP4F2 gene at the mRNA level. The tissue samples used for ovary TissueScan panel were obtained from independent patients, covering various disease stages, and matched normal control tissues are included for comparison between normal and disease samples. The amount of PCR product is correlated with the amount of original RNA transcript and data are expressed in arbitrary units. A representative of two independent experiments is shown. *indicates tumor samples demonstrating striking up-regulation in CYPF2 enzyme.
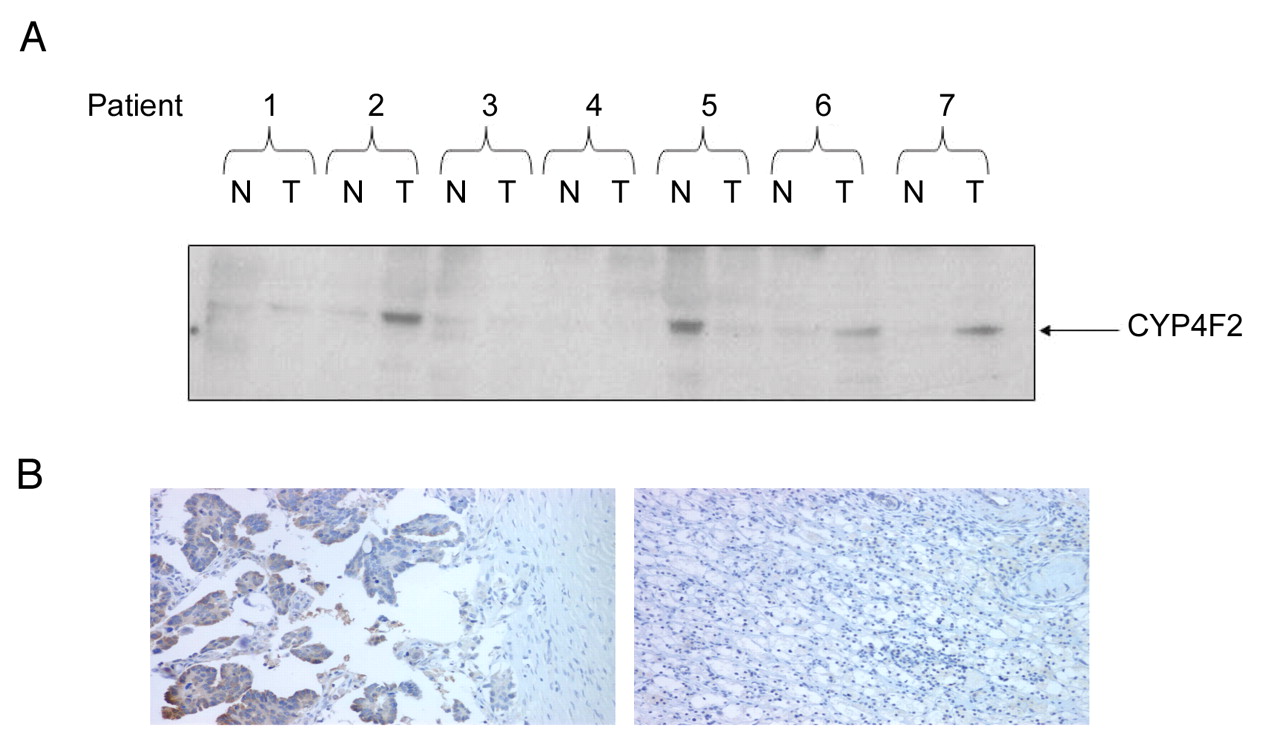
Expression of CYP4F2 enzyme at protein the level is up-regulated in ovarian cancer tissue samples. (A) Western blot analyses of OncoPair INSTA-Blot PVDF membrane containing; 14 lanes of alternating ovary tumor (T) and matched normal adjacent (NA) tissue protein lysates from 7 patient donors using isoform specific CYP4F2 antibody. (B) Immunohistochemical staining for CYP4F2 (brown precipitate) in ovarian cancer (left panel) and matched normal tissue (right panel). Cell nuclei and cytoplasm were counterstained with hematoxylin and eosin.
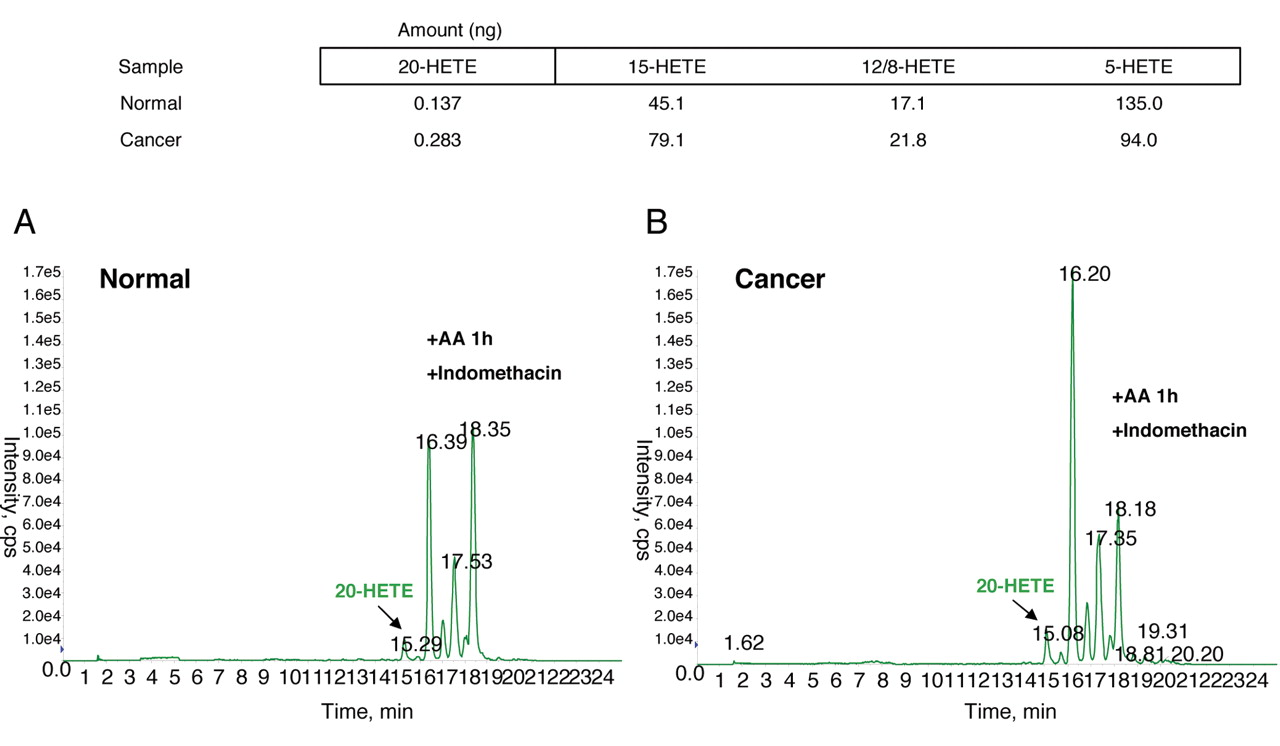
Increased metabolism of AA by CYP4A/F enzymes in ovarian cancer tissue samples. 1 mg/ml of (A) normal ovary and (B) human ovary cancer tissue homogenates was incubated with exogenous AA (40 μM), 1 mM NADPH and 2 μM indomethacin for 1 h at 37°C in the presence of 100% O2. AA metabolites were further extracted and subjected to LC–MS/MS, as described in Materials and Methods to detect levels of 20-HETE.
Expression of the CYP4F2 enzyme at the protein level is up-regulated in ovarian cancer. Since CYP4F2 is the major enzyme involved in the production of 20-HETE in man (14) and antibodies are available for this specific isoform, we further investigated whether the expression of CYP4F2 is also elevated at the protein level in ovarian cancer. Western blot analysis of ovarian cancer patients showed an increase in the CYP4F2 gene at the protein level; where three out of seven patients demonstrated an increase when compared to normal adjacent tissue (Figure 3A). CYP4F2 overexpression at the protein level was further confirmed by immunohistochemistry analysis of papillary adenocarcinoma of the ovary compared to normal ovaries (Figure 3B).
Increased metabolism of AA by CYP4A/F enzymes is observed in ovarian cancer. We further examined whether ovarian cancer homogenates produce higher levels of 20-HETE when incubated with AA as compared to normal tissue. Liquid chromatography/mass spectroscopy (LC–MS/MS) analysis revealed that ovary tumor samples showed increased synthesis of 20-HETE relative to what was observed for normal tissue samples (Figure 4).
20-HETE signaling through activation of the small-GTPase Ras. To determine whether 20-HETE can activate small GTPase Ras, HRPTEC cells were treated with 20 μM WIT003 (20-HETE agonist) and incubated with Ras binding domain (RBD) of Raf1, to specifically isolate the active GTP-bound form of Ras. Western blot detection with anti-Ras antibody demonstrated that WIT003 increased the level of GTP-bound Ras after 5 min of treatment relative to untreated cells, which was similar to EGF effect. Ras remained activated for at least 20 min after WIT003 stimulation (Figure 5).
Discussion
Recent development and characterization of selective and potent inhibitors of the CYP4A/F enzymes and antagonists of 20-HETE signaling (9, 10, 13), as well as the development of sensitive LC–MS/MS assays for the measurement of 20-HETE levels in biological samples (15), have suggested that 20-HETE plays a role in tumor growth and progression, at least in animal models. However, very little work has been done to determine whether expression of the enzymes that synthesize 20-HETE, as well as levels of 20-HETE are elevated in human cancer tumor samples. The present study is the first to demonstrate that the expression of CYP4A/F enzymes is elevated in human cancer tissue samples. We found that the expression of message for CYP4A/F enzymes are markedly elevated in four (thyroid, breast, colon and ovarian) of the eight types of cancer samples, relative to normal tissue controls. Furthermore, western blotting, immunohistochemistry and LC–MS/MS analysis showed that the expression of the CYP4F2 protein level and the production of 20-HETE was increased in human ovarian cancer samples.
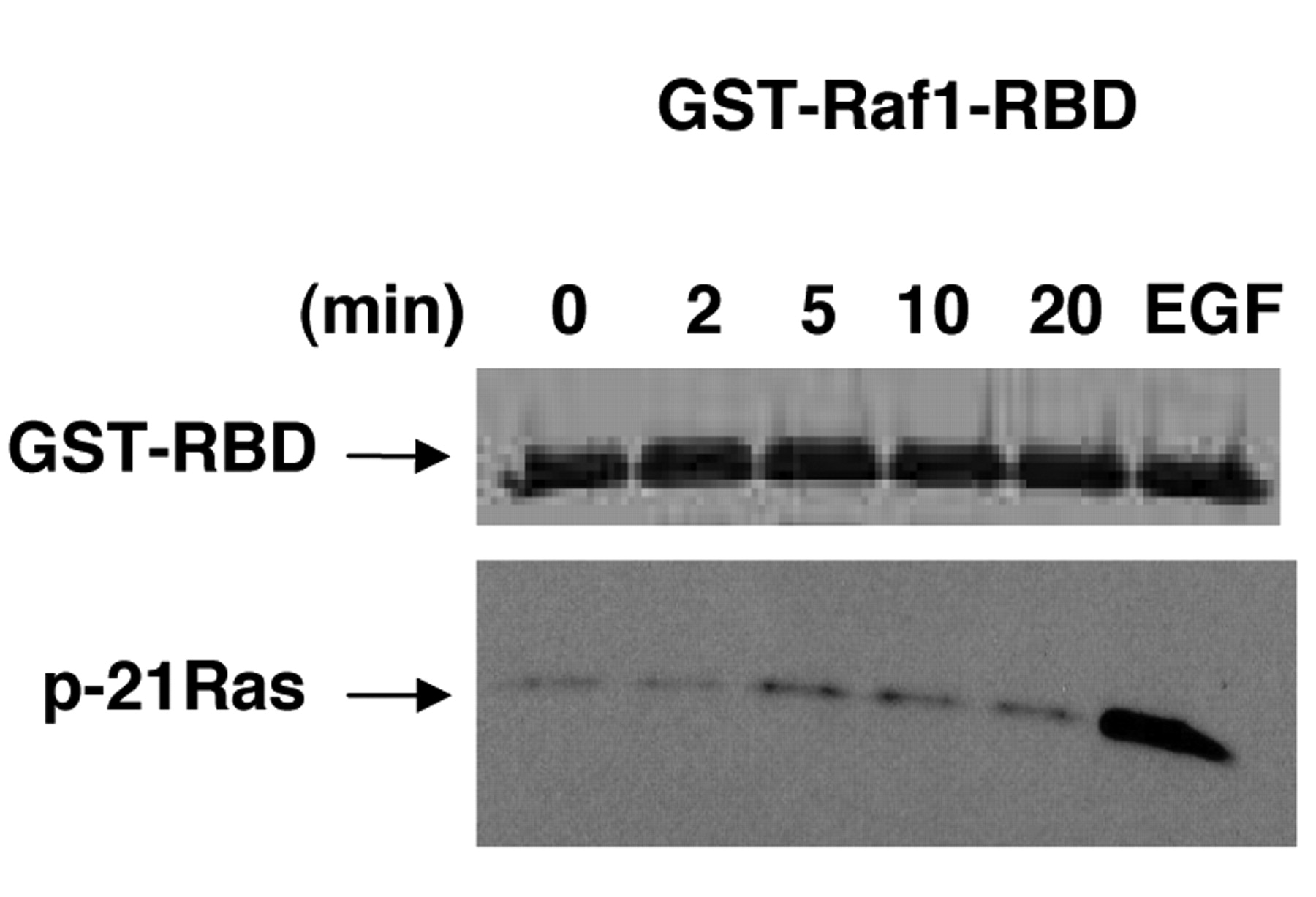
20-HETE signaling in HRPTEC cells through activation of the small-GTPase Ras. HRPTEC cells were serum-starved overnight before the addition of 20 μM WIT003 for the times indicated (min). GTP-bound active Ras was isolated from the stimulated lysates by utilizing an Active GTPase pull-Down and a Detection Kit as described in Materials and Methods, followed by western blot analysis with the p-21Ras antibody (lower panel). The GST fusion protein (~42 kDa, upper panel), was detected by staining the membrane with Ponceau S to demonstrate equal loading. Untreated cells and EGF-treated cells were used as negative and positive controls respectively, and a representative of three separate experiments is shown.
In the analysis of CYP4A/F enzyme expression in human cancers by real time-PCR, we detected some level of variability in the relative p450 content in the tumor samples; with some individuals showing striking overexpression in the CYP4A/F enzymes versus others that do not. We did not have enough tissue samples to establish a correlation between the stage of the disease (metastatic versus primary tumors) and the expression of CYP4A/F isoforms. It is not surprising that there will be variation in the expression of these isoforms among patients and even within regions of the tumors (16). The major point of the current study is that the expression of CYP4A and 4F enzymes is up-regulated in a wide variety of tumors collected from some patients.
We further, additionally, demonstrated that the mechanism of signaling of 20-HETE in regulation of cell growth is via activation of the small-GTPase Ras in HRPTEC cells. 20-HETE activation of small-GTPase Ras is consistent with previous findings by others that 20-HETE can act as a second messenger in the mitogenic actions of some growth factors and activate the ERK and PI3K-Akt cascades (17). To overcome the obstacle of the presence of mutated form of Ras in cancer cells, we have selected a normal human cell line, HRPTEC to study 20-HETE signaling by small-GTPase Ras. In the absence of authentication of the 20-HETE transmembrane receptors, the alternative that intracellular signaling molecules such as Ras serve as direct intracellular targets of 20-HETE molecules can't be excluded.
In summary, the results of previous studies have indicated that inhibitors of the synthesis of 20-HETE block the proliferation and growth of brain and renal tumors in animal models (6, 7). In the present study, we provide the first evidence indicating that the expression of CYP4A/F enzymes and the production of 20-HETE are elevated in human cancer tumor samples obtained from some patients. Future clinical studies using a larger number of tumor samples will be required to establish the frequency of up-regulation of CYP4A/F expression in different types of human cancers, but the present studie provides the first rationale for undertaking this type of analysis.
Acknowledgements
Grant support: Johnson and Johnson grant (Roman), Midwest Affiliate American Heart Association, Predoctoral Fellowship (Alexanian), Wisconsin Breast Cancer Showhouse Research Grant (Sorokin), NIH grants RO1DK41684-19 (Sorokin), R21DK088018 (Sorokin), RO1HL036279 (Roman), RO1DK052448 (P.I.-Raymond) and Fraternal Order of Eagles Research Grant (Sorokin).
- Received June 11, 2012.
- Accepted June 22, 2012.
- Copyright© 2012 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Deciphering the Role of Fatty Acid-Metabolizing CYP4F11 in Lung Cancer and Its Potential As a Drug Target
- Expression and Functional Characterization of Breast Cancer-Associated Cytochrome P450 4Z1 in Saccharomyces cerevisiae
- 20-HETE Signals Through G-Protein-Coupled Receptor GPR75 (Gq) to Affect Vascular Function and Trigger Hypertension