


Abstract
Background: Breast cancer is the leading female cancer in the UK. Recent work has shown that Bone Morphogenetic Proteins (BMPs) and their receptors may be involved in the progression of breast cancer. The aim of the current study is to identify the role of BMPR-IB, one of these receptors, in breast cancer. Materials and Methods: Expression of BMPR-IB was examined in a cohort of breast tissue samples. The transcript level was determined using quantitative real time-PCR and protein levels were assessed with immunohistochemical staining (IHC). Constructed ribozyme transgenes were used to knock-down BMPR-IB in MDA-MB-231 cells, and the effect this had on in vitro cell growth was examined. Results: Decreased staining of BMPR-IB was seen in most of the breast tumour samples examined compared to normal breast tissues. Q-PCR analysis revealed lower expression levels of BMPR-IB transcript in patient samples with a predicted poor prognosis (Nottingham prognosis index (NPI) >5.4) (8.2±17, p=0.03) compared to samples with a good prognosis (NPI <3.4) (469±244). Higher BMPR-IB expression was seen in samples from disease free patients compared to those with poorer prognosis; including patients with metastasis, local recurrence and breast cancer deaths, (p=0.026). Furthermore, down-regulation of BMPR-IB in MDA-MB-231cells led to a promotion of in vitro cell growth. Conclusion: This study shows that decreased expression of BMPR-IB correlates with poor prognosis in breast cancer patients and leads to increased cell proliferation of breast cancer cells in vitro. This suggests that BMPR-IB mediates an inhibitory effect on breast cancer cells.
Breast cancer is the leading type of cancer in Western countries, and only follows lung cancer with regards to death rates (1). The majority of deaths resulting from breast cancer are due to the metastasis of the tumour cells at different sites of the body. Breast cancer most frequently metastasizes to the bone. Although this process is poorly understood, several protein families have been implicated in the progression and development of breast cancer (2).
One of these families is known as the transforming growth factor beta (TGF-β) superfamily which is involved in regulating processes including angiogenesis, differentiation and cell proliferation (3). TGF-β mainly acts as a potent growth inhibitor in normal breast epithelial and breast cancer cells. Breast cancer cells that attain resistance to TGF-β induced growth inhibition are involved in malignant progression and metastasis (4).
Bone Morphogenetic Proteins (BMPs), belong to the TGF-β superfamily. BMPs were first described by Urist et al. in 1965 (5) and now include over 20 proteins. They are pleiotropic growth-factors that play a role in embryonic development and regulate a wide range of important processes including proliferation, apoptosis, differentiation and invasion in many different cell types (6). BMPs have been implicated in the pathogenesis of a number of tumour types including prostate, colorectal, osteosarcomas, myelomas and breast cancer (7). However, unlike the well established role of TGF-β in breast cancer, the relationship of BMPs to breast cancer remains unclear, and sometimes controversial. It was recently shown that BMP signalling enhances the invasion and bone metastasis of MDA-MB-231 breast cancer cells via the Smad-dependant pathway (8).
Aberrant expression of BMPs has been indicated in breast cancer. BMP-4 and 7 have both been shown to be overexpressed in breast cancer cells (9), with BMP-7 expression being inversely correlated with breast cancer cell line invasiveness (10). BMP-2 has diverse effects on breast cancer cells depending on the cell lines examined and culture conditions. It was demonstrated that BMP-2 can inhibit the proliferation of breast cancer cells (11). Meanwhile, it was also shown to inhibit apoptosis thereby promoting survival of MCF-7 breast cancer cells (12).
Sequences of primers used in this study.
BMPs act via their specific receptors (BMPRs), of which there are two types: type I and II serine/threonine kinase receptors. There are seven type-I TGF-β receptors, of which 6 are capable of binding BMPs: Activin receptor-like kinases (ALK) 1-6. Of the 5 type-II receptors, only BMP receptor type II (BMPR-II), Activin A receptor type-IIB (ACVRIIB), and ACVRIIA bind BMPs (13). Once the BMP ligands bind to their corresponding receptors they are capable of inducing either the Smad-dependant or Smad-independent pathway. The former is activated when BMPs bind a preformed heteromeric receptor complex (PFC) (14). This leads to the activation of the type-I receptor by the type-II receptor, allowing the type-I receptor to phosphorylate the receptor-mediated Smads (R-Smads) at their C-terminal. R-Smads 1, 5 and 8 are activated by BMP receptors, whereas Smad 2/3 are activated by TGF-β receptors. Smads 6 and 7 are known as inhibitory Smads and compete with the R-Smads for receptor binding as a way of regulating BMP signalling. The activated R-Smads then complex with Smad 4, which translocates into the nucleus and induces BMP responsive gene expression (15). The p38 MAPK pathway (Smad-independent) is activated when BMPs firstly bind to the type-II receptor and the type-I receptor is then recruited (14).
Germline mutations in BMPR-IA have been indicated in the development of juvenile polyposis (16). In reference to cancer, very little has been reported on the role of BMP receptors. Frequent loss of expression of the 3 main BMP receptors (BMPR-IA, BMPR-IB and BMPR-II) demonstrated that these receptors may be protagonists in the progression of prostate cancer (17, 18). Expression of the three BMPRs has been reported in MDA-MB-231 and MCF-7 breast cancer cells (12). BMPR-IA expression was reported to prevent the metastasis of breast cancer cells in vivo (8), whereas knocking down BMPR-II expression had a growth inhibitory effect (19). BMPR-IB, which is the focus of this paper, has been previously reported to represent a major hallmark in breast cancer progression and has been linked to poor prognosis (20).
The role of growth and differentiation Factor 9a (GDF9a) and BMP-15 (GDF9b) and the clinical significance of BMP-1-7 in breast cancer have been previously reported (21, 22). Due to the obvious role of BMPs in breast cancer found in such reports, this study focuses on the role of the type-I receptor (BMPR-IB) used by GDF9b for its downstream signalling, in the progression of breast cancer.
Materials and Methods
Breast tissue samples. Primary breast cancer tissue (n=124) and non-affected normal breast tissue (n=32) from the same patients was acquired immediately following surgery and placed in a deep freezer until use. The presence of tumour cells in the acquired tissues was confirmed by a consultant pathologist staining the frozen sections with H&E.
Cell culture. Breast cancer cell line MDA-MB-231 was purchased from the European Collection of Animal Cell Cultures (ECACC, Salisbury, UK). The cells were maintained in D-MEM F12 media supplemented with 10% FBS, antibiotics (penicillin and streptomycin), and L-glutamine. Cells were kept at 37°C in a humidified incubator with 5% CO2.
Tissue processing, RNA extraction and cDNA generation. Total RNA was isolated from sections of each frozen tissue sample by homogenisation using a hand held homogeniser. RNA from MDA-MB-231 cells grown in a monolayer at 85-90% confluency, were lysed. RNA isolation was carried out following the protocol of ABgene total RNA Isolation Reagent kit (ABgene, Epsom, England, UK). The concentration of the extracted RNA was quantified using a UV spectrophotometer and 1 μg was then converted into cDNA by reverse transcription using the iScript™ cDNA Synthesis Kit (Bio-Rad, Hammel Hemstead, UK).
Knockdown of BMPR-IB in MDA-MB-231cells. BMPR-IB expression was knocked-down in MDA-MB-231cells using antihuman BMPR-IB hammerhead ribozyme transgenes, which were designed based on the secondary structure of BMPR-IB using Zuker's RNA mFold program (23). The specific ribozymes were cloned into mammalian expression plasmid vector GFP (Invitrogen Ltd., Paisley, UK). Control empty plasmid vectors and the ribozyme transgenes were then transfected into MDA-MB-231 cells and underwent selection with G418 (Melford Laboratories LTD., Chelsworth, Ipswich, UK) for approximately 3 weeks before further use.
Q-PCR. Expression levels of BMPR-IB were quantified in both the MDA-MB-231 cells and the breast tissue samples using real-time quantitative PCR and the icycler IQ system (Bio-Rad, Hammel Hemstead, UK). The reaction consisted of Hot-Start-Q-master mix (ABgene, Epsom, England, UK), 10 pmol of specific forward primer, 1 pmol reverse primer that has a Z sequence attached to it (5′-ACTGAACCTGACCGTACA-3′) which is complementary to the universal Z probe (TCS Biologicals Ltd., Oxford, UK), and the Z probe (refer to Table I for further details). BMPR-IB expression was quantified at the same time as β-actin (cDNA standard) in all samples under the following conditions: 94°C for 10 minutes, 50 cycles of 94°C for 30 seconds, 56°C for 30 seconds and 72°C for 1 minute, and then a final extension at 72°C for 10 minutes. Levels of BMPR-IB transcript were normalised according to the corresponding β-actin level, which was used for statistical analysis.
Summary of clinical and pathological data of tissue cohort used in this study.
Western Blotting. Protein isolated from the MDA-MB-231 cells was quantified using the DC protein assay kit (Bio-Rad, Hammel Hemstead, UK) and an ELX-800 spectrophotometer (BioTek Instruments, Inc., Winooski, VT, USA). Equal amounts of each protein sample were separated using SDS-PAGE, and then blotted onto nitrocellulose sheets. Goat polyclonal BMPR-IB antibody (SC-5679) was used to probe the blot, followed by a secondary peroxidise conjugated antibody. Protein bands were visualised using the supersignal West Dura system (Pierce Biotechnology, Inc., Rockford, IL, USA).
Immunohistochemistry. Frozen specimens from the normal and breast tumour samples were cut to a thickness of 6 μm before being fixed and blocked with horse serum. This was followed by one hour incubation with or without (control) a BMPR-IB antibody and then a secondary biotinylated antibody. The secondary antibody, along with an avidin biotin complex was used to detect BMPR-IB expression in compliance with the Vecastain Universal Elite ABC kit protocol (Vector Laboratories, Peterborough, UK). Sections were then stained using standard conditions and finally counterstained with Gill's Haematoxylin.
Summary of Q-PCR results. BMPR-IB transcript levels were statistically analysed. Numbers in bold are those found to be statistically significant.
In vitro cell growth assay. Cells were seeded into a 96 well plate at a concentration of 2,500 cells per well, and incubated for up to 5 days. The cells were then stained with crystal violet and absorbance was measured at 540 nm with a spectrophotometer (Bio-tek ELx800).
Statistical analysis. Normally distributed data was analysed using a two sample t-test, while non-normally distributed using Mann-Whitney and Kruskal-Wallis tests were used as statistical analysis.
Results
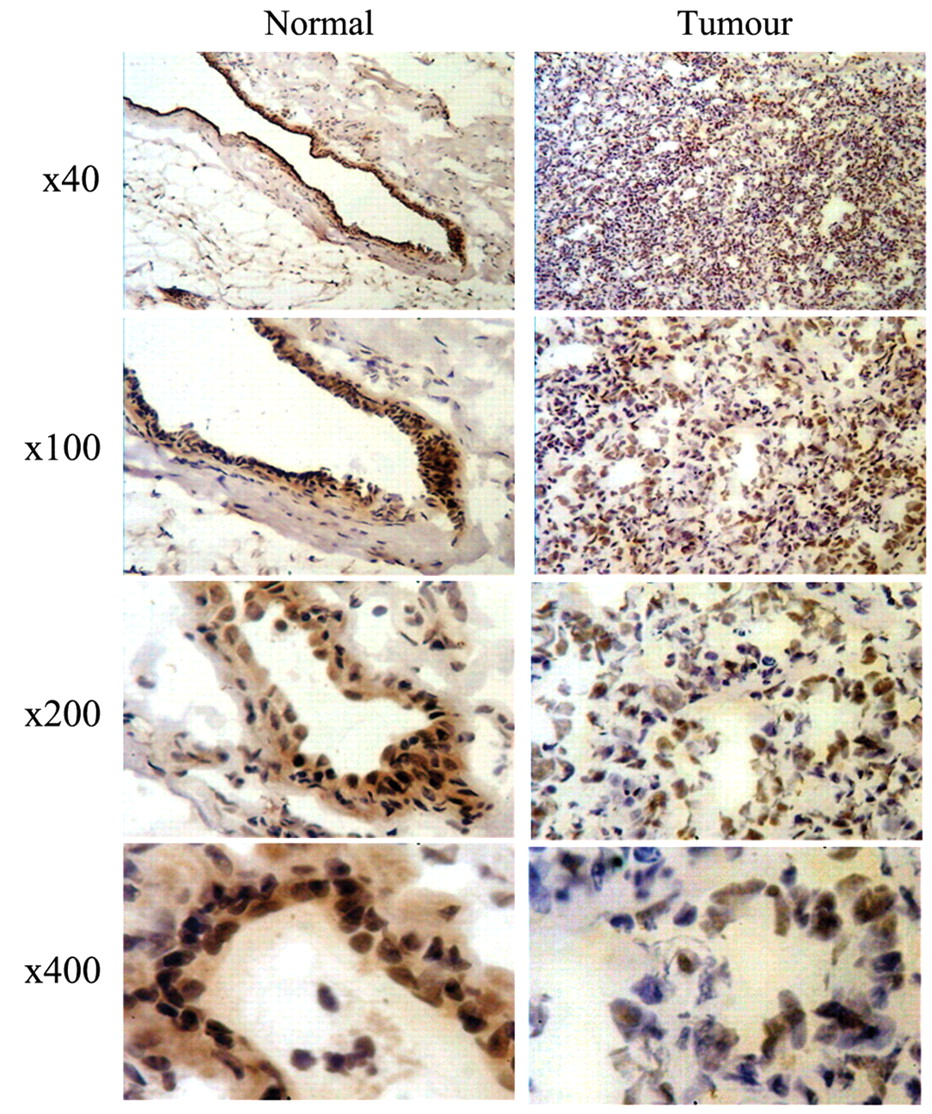
Expression of BMPR-IB in normal breast and breast cancer tissue. A cohort of normal and breast tumour samples were examined for expression of the BMP type I receptor, BMPR-IB. Table II shows the clinical and pathological data of the tissues used, while Figure 1 displays the characteristic results of immunohistochemistry. It shows that intensive BMPR-IB staining was confined to the luminal epithelial cells of normal breast tissue. Decreased expression of BMPR-IB was seen in most of the breast tumour samples examined compared to normal breast tissue.
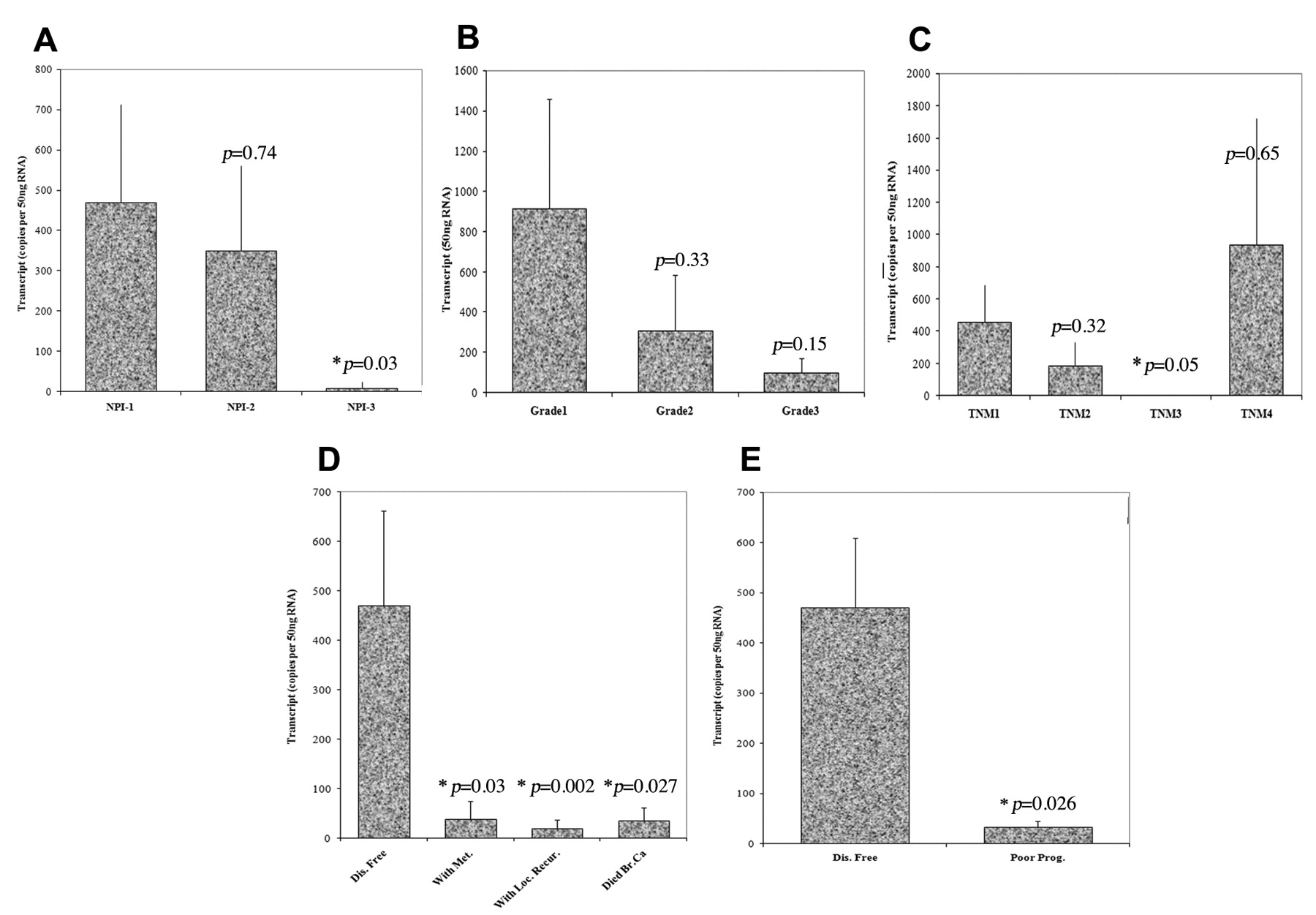
Prognostic value of BMPR-IB expression in breast cancer patients. In order to investigate the association between BMPR-IB expression status and breast cancer prognosis, BMPR-IB transcript levels were determined in the same cohort of breast tissue, using Q-PCR. These results are summarised in Table III, and those that have been deemed significant, are represented in graphical format in Figure 2. BMPR-IB expression was demonstrated to be inversely correlated with the Nottingham prognosis index (NPI) (Figure 2A). Lower expression levels of BMPR-IB were detected in patient samples with NPI greater than 5.4 (poor prognosis group) (8.2±17, p=0.03) compared to samples in the good prognosis group (469±244 copies).

IHC showing staining of BMPR-IB in normal breast tissue and breast tumour tissue at varying magnifications. Frozen tissue was stained with antibody against BMPR-IB. The level of expression of BMPR-IB was notably higher in normal breast compared to breast cancer tissue.

Q-PCR results showing transcript levels of BMPR-IB in breast cancer patient cohort. A. Nottingham prognosis index (NPI). NPI=0.2xT+N(1-3)+G(1-3). T: Max. tumour diameter, N: Number and level of nodes metastasised, G: Elston and Ellis grade. NPI-1(<3.4)=good prognosis, NPI-2(3.4-5.4)=moderate prognosis, NPI-3(>5.4)=poor prognosis. B. Tumour grade. C. Tumour nodal metastasis (TNM) stage. D. Disease free, with metastasis, with local recurrence, and died from breast cancer. E. Disease-free versus poor prognosis.
Furthermore, there was a trend seen between BMPR-IB expression and tumour grade. BMPR-IB levels decreased with worsening tumour grade and tumour differentiation (Figure 2B). Higher levels were detected in the well differentiated tumours (915±542) compared with the poorly differentiated tumours (102±69, p=0.15). However, despite this trend, the data was not found to be statistically significant. A similar trend was seen in reference to the tumour node metastasis (TNM) staging (Figure 2C). Patient samples with a TNM stage of 3 (poor prognosis) (0.4±0.1) had decreased BMPR-IB transcript levels. BMPR-IB expression was also assessed according to the clinical outcome of the patients. Higher BMPR-IB levels were seen in tumour samples from patients who remained disease-free (470±192) compared to patients with poor prognosis (33.7±18, p=0.026 vs. disease free), including those with metastasis (38±37, p=0.03), local recurrence (19±18, p=0.002) and those who died from breast cancer (35.6±26, p=0.027).
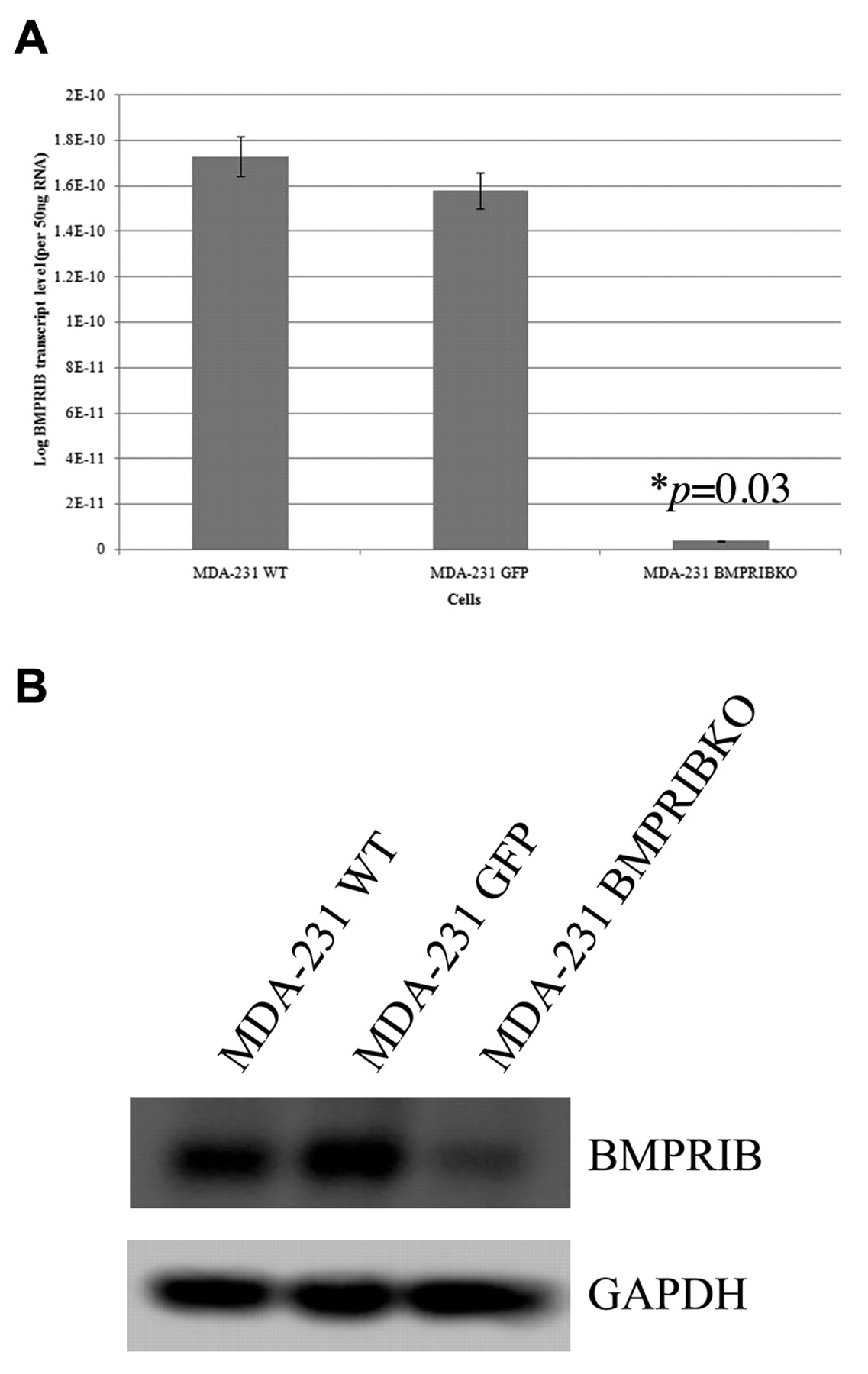
Knocking down BMPR-IB expression in MDA-MB-231 breast cancer cell line. In order to elucidate the role of BMPR-IB in vitro, hammerhead ribozymes were used to down-regulate BMPR-IB expression in the highly metastatic breast cancer cell line, MDA-MB-231. This transfected cell line is further referred to as MDA-MB-231 BMPRIBKO. Figures 3A and B demonstrate this down-regulation at both the mRNA and protein level as revealed by Q-PCR and western blotting. Reduced BMPR-IB expression was detected in the MDA-MB-231 BMPRIBKO (4.3×103±7.52×10-3) cells compared to both the wild-type (MDA-MB-231WT) and empty plasmid control (MDA-MB-231GFP) (2.14×106±2.96×102, p=0.03).

Down-regulation of BMPR-IB expression in MDA-231cells by ribozyme transgene. A. Western blot showing decrease at protein level. B. Q-PCR showing logarithmic amount of BMPR-IB transcript level.
The in vitro effect of BMPR-IB knockdown in breast cancer cells. Following down-regulation of BMPR-IB, the cell proliferation rate of MDA-MB-231 was assayed in order to determine if there were any noticeable changes on the growth rate. Figure 4 is a representative graph showing the growth rate taken at day three. As can be seen, there was a significant promotion of the cell growth rate in the MDA-MB-231 BMPRIBKO (231.7% ±30.2%) cells compared to both MDA-MB-231WT (179.7% ±4.3%) and MDA-MB-231GFP (170.3% ±17.6%, p=<0.01 vs. both controls). This suggests that growth promotion is indeed due to the down-regulation of BMPR-IB expression.
Discussion
The role of TGF-β in breast cancer has undergone extensive study and has been shown mainly to have a growth inhibitory effect on breast cancer cells (4). For this reason, many breast cancers lose expression of TGF-β in order to become refractive to this growth inhibitory effect, as has been demonstrated with TGF-βRII (24, 25) and TGF-βRI (26). The role of BMPs in breast cancer is less clear. However, it has been recently shown that GDF9a and BMP-15 have an inhibitory effect on breast cancer cells (21) and several other BMPs (BMP-2, -6 and -7) have been shown to have a similar effect (11, 22, 27). In the case of BMPRs, several studies have shown conflicting results of their role in breast cancer.

Growth rate of MDA-MB-231 cells taken at day 3. Absorbance of crystal violet was measured at 540 nm and the percentage growth rate was calculated.
The current study has shown that loss of expression of BMPR-IB in breast cancer correlates with a high NPI score, a late pathological stage and poor prognosis in patients including those with metastasis, local recurrence and those who died of breast cancer. Also, BMPR-IB protein levels were shown to be reduced in breast tumour tissue compared to normal breast epithelium. Finally, when BMPR-IB expression was down-regulated in the highly metastatic breast cancer cell line MDA-MB-231, this led to a promotion of the cell growth rate of these cells in vitro. These results suggest that loss of expression of BMPR-IB may aid in the development and progression of breast cancer. As previously reported, several BMPs that use BMPR-IB as a downstream receptor (BMP-2, -6, -7 and -15) have been shown to have inhibitory effects on breast cancer cells, and hence down-regulation of BMPR-IB in these cells would abrogate this effect (7, 28).
These results are consistent to those previously reported in prostate cancer, where loss of expression of BMPR-IA, BMPR-IB and BMPR-II appeared to be a common occurrence (17). Furthermore, low BMPR-II expression was correlated with poorly differentiated tumours, high tumour grade and poor prognosis (18). Both breast and prostate tumours most commonly metastasise to the bone where BMPs are highly concentrated and act to negatively regulate cell proliferation. Therefore, loss of BMPR expression would aid these cells in metastasising to the bone allowing them to thrive in the bone microenvironment (18).
In breast cancer however, previous reports appear to conflict with the results of the current paper. Loss of BMPR-II expression was shown to induce growth arrest by blocking the P38/MAPK pathway, hence promoting breast cancer cell survival (19). BMPR-IA down-regulation meanwhile was shown to inhibit the motility and osteolytic metastasis of breast cancer cells (8). Finally, Helms et al. demonstrated that high BMPR-IB expression correlated with pro-proliferation, poorer tumour grade, cytogenetic instability and poor prognosis in oestrogen-receptor positive breast cancer cells (20).
There are several potential explanations for these discrepancies. Firstly, the clinical data in this current study is supported by the in vitro work carried out, where down-regulation of BMPR-IB promoted proliferation of MDA-MB-231cells. In the case of Helms et al., no in vitro work was carried to confirm the results from the patient tissue cohort. Also, they mention that the pro-tumourigenic effect reported was only in oestrogen-receptor positive breast cancers. MDA-MB-231, and the majority of the tissue samples used in this current paper are oestrogen-receptor negative. It has been previously reported that oestrogen can suppress the function of BMPs by interfering with Smad transcriptional activity and by inhibiting BMP gene expression (including BMPR-IA and -IB) (27). The oestrogen-receptor status of breast cancer cells may therefore influence the effect that BMPs have on them.
Other reasons for conflicting results may be due to the contradictory effect of BMPs on breast cancer. Some claim BMPs have a pro-tumourigenic effect (8), whereas others suggest an inhibitory effect (17, 18). It is possible that like TGF-β, BMPs initially inhibit breast cancer cells, but promote tumourigenesis during later stages of breast cancer progression (29). Furthermore, the cellular effect of BMPs depends on many factors, such as cell type and the expression status of BMP antagonists and receptors, which can regulate activation of different pathways (30).
For all the aforementioned reasons, it is therefore plausible that BMPR-IB has varying effects on breast cancer, depending on several cellular factors, including oestrogen-receptor status. In the case of this current study however, it can be concluded that expression of BMPR-IB may be lost during the progression of breast cancer in order to abrogate the inhibitory effect of BMP signalling. BMPR-IB may therefore act as a tumour suppressive molecule in breast cancer and may be useful as a possible molecular marker in breast cancer patients. Further work has to be carried out in order to elucidate the mechanisms behind this tumour suppressive effect.
Acknowledgements
The authors would like to thank Cancer Research Wales, The CURE Fund, and The Fong Family Foundation for supporting their research.
Footnotes
- Received January 20, 2009.
- Revision received February 12, 2009.
- Accepted February 16, 2009.
- Copyright© 2009 International Institute of Anticaner Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- The Activation of MEK/ERK Signaling Pathway by Bone Morphogenetic Protein 4 to Increase Hepatocellular Carcinoma Cell Proliferation and Migration
- Transforming Acidic Coiled-coil-containing Protein 2 (TACC2) in Human Breast Cancer, Expression Pattern and Clinical/Prognostic Relevance
- The Transforming Growth Factor-{beta} Type III Receptor Mediates Distinct Subcellular Trafficking and Downstream Signaling of Activin-like Kinase (ALK)3 and ALK6 Receptors