


Abstract
Carcinogenic polycylic aromatic hydrocarbons can alter immune responses. Changes in immune response gene expression profiles in multiple human mammary cell strains exposed to benzo(α)pyrene (BP) (4 μM) in vitro, in the presence or absence of chlorophyllin (5 μM), were observed using Affymetrix gene arrays. Expressions of five immune response genes were altered ~3.0-fold by BP exposure and 24 genes by BP in the presence chlorophyllin. In silico pathway analysis revealed altered immune response genes form interactive gene networks with many cellular processes, suggesting their role in a complex multigenic response to toxins. Additionally, it was suggestive of the possible immunomodulatory potential of chlorophyllin apart from various other well-documented mechanisms of action. Gene expression matrices revealed consistent alteration patterns involving IL1B, SECTM1 and CXCL14 on exposure to BP, and IL1RN, CD86, IF144 and GIP2 in the presence of chlorophyllin and BP, suggesting some of these genes might constitute putative immune response biomarkers of PAH exposure. This study has therefore identified a battery of potential immune response biomarkers of PAH exposure, amidst several genes, for future validation studies.
Abbreviations: BP, benzo(a)pyrene; NHMEC, normal human mammary epithelial cell; PAH, polycyclic aromatic hydrocarbon.
Polycyclic aromatic hydrocarbons (PAHs) are formed mainly from the incomplete combustion of organic materials (1). They are a class of ubiquitous environmental pollutants, many of which are carcinogenic (2). To exert their carcinogenic effects they require metabolic activation to reactive electrophiles that can damage DNA (3). Because cytochrome P450 enzymes are involved, the redox cycling activity that accompanies metabolic activation of PAHs generates oxygen free radicals that can result in additional DNA damage (4, 5).
In addition to carcinogenicity, PAHs have been implicated in toxicity to many processes in mammalian systems, including cell-mediated and humoral immunity (6). The PAHs, including benzo(α)pyrene (BP), can have broad ranging effects in mammalian systems, for example, they have been shown to act synergistically on systemic immune responses (7, 8), be potent immunosupressants and immunotoxicants (9), and induce pro-inflammatory cytokines (10). They have also been shown to modulate expression of immune response genes in human biological systems (11).
Development of effective cancer intervention agents is an important goal in human cancer prevention where exposures to ubiquitous carcinogens, such as BP, are difficult to control. Chlorophyllin, a water soluble metalloporphyrin that is known to have anti-mutagenic and anti-carcinogenic properties, can mitigate the DNA-damaging effects of BP in part by preventing or reducing PAH-mediated P450 induction (12). Due to its potential to activate a variety of gene pathways, through common or related receptor molecules, chlorophyllin was evaluated for its ability to modulate BP-dependent immune response gene expression. To do this, a primary normal human mammary epithelial cell (NHMEC) model system, comprised of a panel of cell strains derived from different donors, was used (13-15).
A major objective of this study was to identify immune response biomarkers of carcinogen exposure and their modulation in the presence of chlorophyllin. Microarray observations from the same dataset that focused on carcinogen metabolism and oncogenes have been previously published (16); here the focus is on genes involved in immune responses and their interactions with multiple other cellular processes.
Materials and Methods
Cell culture and treatments. Normal breast epithelial cell strains (M98035, M99005, M98015, M98025, M99025, M99016) were developed from normal human breast tissue discarded at reduction mammoplasty and maintained as described in detail elsewhere (13-15). These cell strains were a smaller subset of the cell strains used by Keshava et al. (17). Briefly, breast tissues obtained from healthy women undergoing reduction mammoplasty (Cooperative Human Tissue Network sponsored by the National Cancer Institute and National Disease Research Interchange) were processed to obtain NHMECs that were then subjected to one of three protocols at passage 6. Treatments were as follows: T1, vehicle control (acetone: ethanol, 2:3 at 1:1000 dilution) for 24 h; T2, BP (4 μM) alone for 24 h; T3, pre-treatment with chlorophyllin (5 μM) followed by co-treatment with BP (4 μM) and chlorophyllin (5 μM) together for 24 h. Each treatment was carried out in duplicate.
Microarray analysis.
Target preparation: The generation and hybridization of labeled target on to Hu-Gene 133A arrays and quality control used are described in detail elsewhere (18). Briefly, total RNA reverse-transcribed to cDNA (SuperScript Double-Stranded cDNA Synthesis kit, Invitrogen Life Technologies, Carlsbad, CA, USA, and T7-oligo(dT) promoter primer kit, Affymetrix, Santa Clara, CA, USA) and subsequently in vitro transcribed to cRNA (GeneChip IVT Labeling Kit, Affymetrix) was hybridized on to Hu-Gene 133A arrays (Affymetrix) following fragmentation to 35-200 nucleotide sequences. The hybridized arrays were washed and stained on the GeneChip Fluidics Station 400 (Affymetrix) and subsequently scanned on the HP GeneArray 2500 Scanner (Hewlett Packard, Palo Alto, CA, USA).
Data analysis: “Absolute analysis” to compute absolute signal intensities of the control and treated groups followed by ‘comparison analysis’ to compute fold change (FC) of the treated groups over the control vehicle (T1 versus T2, and T1 versus T3) were carried out using Microarray Suite 5.0. Absolute analysis involves analysis of each of the individual arrays to determine the presence or absence of a particular transcript (Detection call), measure their relative abundance (Signal) and also provide a ‘Detection p-value’ evaluated against a user-defined threshold to determine the Detection call. With Affymetrix arrays, where all probes are arrayed as probe pairs, signals are calculated for each probe set based on One-Step Tukey's Biweight Estimate so as to give the weighted mean. During comparison analysis the expression patterns of the control vehicle-treated cells were set as the baseline and those of the treated groups were compared against them.
Two algorithms were used to define changes in expression patterns (Microarray Suite 5.0, Affymetrix). A change algorithm uses Wilcoxon's Signed Rank test to define a change (increase, decrease or no change) in expression trends and also assign a p-value to the same. A second algorithm then provides a quantitative measure of the magnitude of change in terms of signal log ratio (SLR). This again is calculated by taking the mean of log ratios of probe pair intensities across both the arrays being compared and is done using one-step Tukey's Biweight method (19). The log scale employed is to base 2. Each cell strain with genes possessing a present call (P) in both duplicates and altered by an SLR of at least 1.5 (FC ~3.0) in both the replicates in at least one of the 6 NHMECs were considered altered. Graphical representation of all the altered genes was generated using Cluster and Tree View software (20).
Cross-talks between genes altered by the respective treatments (T1 versus T2 and T1 versus T3) were computed using Pathway Studio Central software (Ariadne Genomics, Rockville, MD, USA). Pathway Studio relies on a proprietary ResNet database powered by Medscan technology to query nearly 500,000+ functional relationships distilled from the entire PubMed and 43 full text journals to automatically extract information from scientific literature, and works with leading public and commercial databases of signaling and biochemical pathways, including KEGG, BIND, GO, STKE Connection Maps and Prolexys HyNet™ to construct cross-talks between the input genes/protein (21). Initially, all possible interactions of the altered genes with all other processes and proteins in the cell were deciphered. Later, only the ‘shortest path’ interactions were computed. Finally, only direct interactions, if any, between the altered genes were deciphered.
Altered genes were also analyzed in terms of having a statistically significant association with well-known and characterized GenMAPP, PharmGKB, KEGG and BioCarta pathways using ArrayXPath software (22).
Validation of microarray gene expression. Confirmation of gene expression on microarray was carried out for a subset of genes altered by SLR ≥1.5 through real-time polymerase chain reaction (RT-PCR). Details of RT-PCR can be found in John et al. (18). In brief, total RNA reverse-transcribed to single stranded cDNA (Advantage RT PCR kit; BD Biosciences, Palo Alto, CA, USA) was used as a template in a reaction mixture (25 μl) with SYBR Green Universal PCR Master Mix (Applied Biosystems, Foster City, CA, USA) and 50 pmol each of the forward and reverse primers (designed using Primer Express v1.5, PE Applied Biosystems) for validation of the expression of a subset of immune response genes altered on Hu-Gene 133A arrays. Each sample was assayed in duplicate and their expression was normalized to that of GAPDH using the 2-ΔΔCT method (23).
Results
The expression of genes belonging to various functional categories was altered by BP and subsequently modulated by chlorophyllin. A list of all functional categories of genes involved predominantly in functions other than immune responses and altered by SLR ≥1.5 in at least one of the 6 NHMEC lines by the respective treatment (T1 versus T2, and T1 versus T3) can be found on the Internet (24) (Tables A, B, C and D). Here, a list of immune response genes altered by SLR ≥1.5 across 6 NHMEC lines is shown (Table I). A more extensive version of this Table can be found on the website (25). SECTM1, which has a role in hematopoiesis as well as immune responses, is presented here (Table I) [as well as in Tables A, B, C and D (24)].
List of immune response genes altered by SLR ≥1.5 in at least 1 of the 6 NHMEC strains on comparison of respective treated groups vs. control vehicle.
A total of only 5 immune response genes were altered by BP of which 4 were up-regulated and only 1 was down-regulated by SLR ≥1.5 in at least one of the 6 NHMEC lines. Twenty-four genes were altered by the combined interaction of BP and chlorophyllin coming together in treatment T3. Of these, 15 were up-regulated and 9 were down-regulated by SLR ≥1.5. The expression of IL1B and SECTM1 were consistently up-regulated by BP in most individuals, with M99016 exhibiting the greatest expression with SLR ≥1.5. Both genes also exhibited the greatest expression in M99016 on comparison of T1 vs. T3 and were therefore represented in the list of genes up-regulated by T1 vs. T2 as well as T1 vs. T3. IL8 was up-regulated by SLR ≥1.5 in two cell strains (M98035 and M99016) but down-regulated by SLR ≥1.5 in M99005 and, therefore, represented in the list of genes up and down regulated by T1 vs. T3.
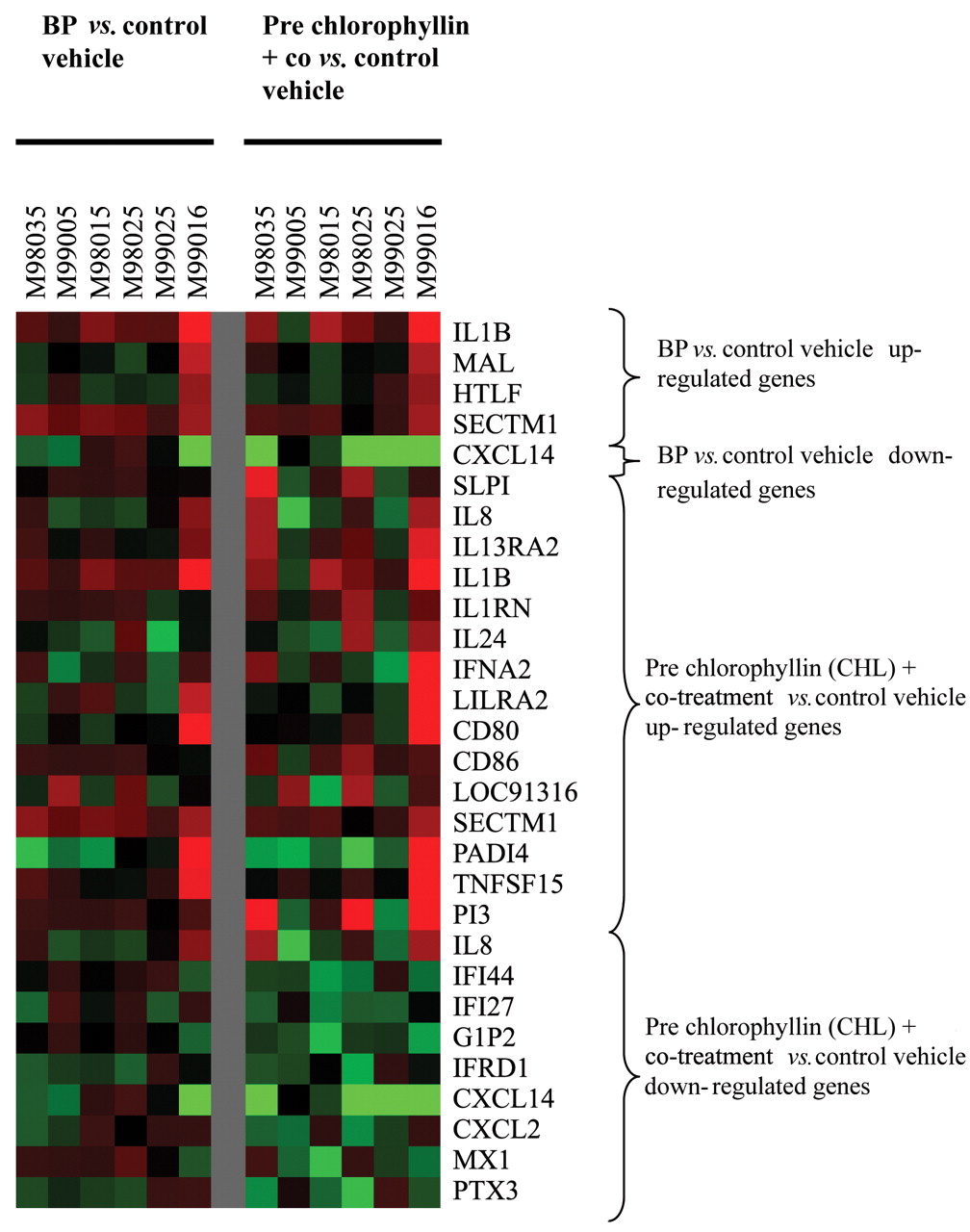
Chemokine ligand 14 (CXCL14) was another gene differentially regulated in different cell strains and, therefore, represented in common in the list of genes down-regulated by T1 vs. T2 as well as T1 vs. T3. A graphical representation of expression patterns of all genes represented in Table I generated by using Cluster and Tree View software is presented in Figure 1.
Results of RT-PCR confirmation of expression levels for a subset of 7 immune response genes altered by SLR ≥1.5 on gene-chip (GC) are presented in Table II. Though there was a good correlation for some genes, the remaining genes correlated moderately to poorly.

Graphical representation of expression patterns of all immune response genes altered by treatment with BP or pre chlorophyllin plus co-treatment generated using Cluster and Tree View software.
When genes altered by T1 vs. T2 were queried for all possible interactions with all other molecules and processes within the cell using Pathway Studio Central, a total of 2,248 relationships/interactions involving 1,485 entities including proteins, small molecules and various functional classes of molecules, complexes and cellular processes were found to be involved. When only ‘shortest cross-talk’ interactions were considered for the above genes, only 60 relationships were found between 249 entities. However, there were no direct interactions between the 5 altered genes (Figure 2). More details about the various interactions among the genes altered by BP can be found on the Internet (25).
Genes altered by T1 vs. T3 were found to exhibit 5,782 interactions among and between 2,299 entities within the cell. Of these, 379 entities including the altered genes were found to be involved in 704 ‘shortest cross-talk’ interactions. However, there were only 51 direct interactions between the altered genes (Figure 3). More details about the various interactions among the genes altered by T1 vs. T3 can be found on the Internet (25).

No direct interactions were observed between the genes altered by BP alone when queried using Pathway Studio Central.

Fifty-one direct interactions were found between genes altered by pre chlorophyllin + co-treatment when queried using Pathway Studio Central. Genes with no direct interactions between themselves are shown as separate entities.
Using ArrayXPath software, 2 (IL1B and MAL) out of the 5 genes altered by T1 vs. T2 were found to have a statistically significant association (p<0.05) with 5 Biocarta pathways (Table III). A total of 45 Genmapp pathways, 9 PharmGKB pathways, 70 KEGG pathways and 346 Biocarta pathways were queried by Array X to obtain the above associations. A total of 2 (CD80 and CD86) out of 23 genes altered by T1 vs. T3 were involved in one GenMapp pathway and 6 genes (SLP1, IL8, IL1B, IL1RN, CD80 and CD86) were found to be involved in 13 Biocarta pathways (Table IV).
Correlation of FC on gene chip (GC) vs. RT-PCR for a selected set of immune response genes altered by SLR ≥1.5 (FC ~3.0).
Discussion
Immune surveillance plays a vital role in protecting the body not just against various pathogens but also against various toxins, chemicals, drugs and foreign particles. It plays a role in preventing the development of cancers and also the elimination of tumor cells (26). Breast epithelial cells have been shown to participate in immune surveillance (27). Other epithelial model systems have also highlighted the importance of the immune system in cancer prevention by showing that impairment of this system as by exposure to chemical carcinogens can contribute to tumor promotion (28). In the processes of manifesting their carcinogenic effects, many carcinogens are known to behave as immunosupressants. BP and other PAHs have previously been shown to be immunotoxicants leading to T-cell suppression (29). BP has also been shown to act as an immunosuppressant leading to suppression of B cell lymphopoiesis (30). Other studies have shown impairment of humoral and cell mediated immune responses in the progeny of BP exposed mice (31).
Pathways in which immune response (IR) altered by SLR ≥1.5 on Hu-Gene133A arrays on comparison of BP treated cells vs. vehicle control are involved. A total of 5 IR genes were altered by SLR ≥1.5, of which 2 (40%) (IL1B, MAL) were found to possess a statistically significant association (p≤0.05) with the above pathways. For genes involved in more than one pathway, a q-value (q≤0.05) accounts for the multiple-comparison.
Pathways in which immune response (IR) altered by SLR ≥1.5 on Hu-Gene133A arrays on comparison of chlorophyllin treated cells (pre-chlorophyllin + co-treated) vs. control vehicle are involved. A total of 23 IR genes were altered by SLR ≥1.5 of which 6 (26%) (SLP1, IL8, IL1B, IL1RN, CD80 and CD86) were found to possess a statistically significant association (p≤0.05) with the above pathways. For genes involved in more than one pathway, a q value (q≤0.05) accounts for the multiple-comparison.
In addition, BP has also been shown to be capable of transforming and immortalizing cells in vitro (15). While modulating BP metabolism along with elimination of some toxic metabolites could form a primary strategy to reduce its adverse effects, stimulation of immune responses could augment defense systems that help prevent early accumulation of mutated and transformed cells (26, 32). In some instances the immune system has also been shown to have a role in the inhibition of BP toxicity via suppression of cytochrome P450-mediated CYP activation and subsequent mutagenesis (33).
Chlorophyllin has been shown to manifest its anti-carcinogenic properties by acting as a CYP inhibitor (12), an antioxidant (34), a stimulator of phase II enzymes (35), an inducer of apoptosis (36), a modulator of carcinogen transport (37) and also by acting as an ‘interceptor’ molecule sequestering and degrading carcinogens (38). Potential modulation of host immune responses may be a new mode of action broadening its capabilities. Some recent studies point to an immunomodulatory role of chlorophyllin although these reports appear to be conflicting (12, 39). The objectives of this study were, therefore, three-fold: to identify immune response biomarkers following carcinogen exposure in the absence and presence of modulators, examine interindividual variations in response patterns and thirdly to examine whether chlorophyllin is also capable of stimulating immune responses in addition to the above documented mechanisms of action.
In light of the above mechanisms of action of chlorophyllin, it is reasonable to assume that the maximum chemopreventive potential will be achieved when pre-dosed, thereby ‘priming’ cells for any potential carcinogenic insult, followed by a co-treatment facilitating chlorophyllin to function as a ‘desmutagen’ as well, reducing carcinogen bioavailability. In fact, earlier studies have shown a maximum reduction in genetic damage in terms of mitigation of CYP1 gene expression and BP-DNA adduct formation when chlorophyllin was dosed as a ‘pre + co-treatment’ dose (unpublished data).
All genes discussed here are listed in Table I. Interleukin 1 beta (IL1B), a proinflammatory cytokine with a proangiogenic role in vivo, was consistently up-regulated in all 6 cell strains exposed to BP. This may suggest that among the many changes occurring following exposure to BP are cell proliferation and angiogenesis. This parallels a study where exposure of rats to Fe2O3 or BP or in combination to study their role in lung cancer resulted in enhanced production of various inflammatory cytokines including IL1B (10).
Another study involving exposure of CL5 human lung adenocarcinoma cells to motorcycle exhaust particulates or BP also found enhanced expression of various inflammatory cytokines. However, interleukin1 alpha (IL1A), a cytokine which produces effects opposite to those of IL1B was found to be up-regulated (40) rather than IL1B. Being an angiogenic factor, IL1B has also been found to be amplified in various types of cancer, potentiating their invasive ability (41, 42). The other genes up-regulated by BP were MAL, HTLF and SECTM1, of which only SECTM1 was altered in a consistent fashion. SECTM1 (K12) is a protease inhibitor thought to have a role in hematopoiesis and also in immunogenic processes by acting as CD7 ligand (42). Though initially unknown, CD7 is now known to be involved in T- and NK cell activation and cytokine production. K12 by interacting with CD7 is therefore thought to play a role in regulation of thymocyte signaling and cytokine release (43). SECTM1 is also one of the 28 uncharacterized genes found to be a potential activators of the NF-κB pathway (44).
Conflicting reports exist regarding the role of CXCL14 (BRAK), the only member down-regulated by BP. While it was found to play a role in tumor suppression inhibiting tumor growth and angiogenesis in some studies (45, 46) it was reported in another study to enhance proliferation, migration and invasion of myoepithelial cells and myofibroblasts thereby promoting breast tumorigenesis (47).
Fifteen genes were up-regulated by the combined effect of BP and chlorophyllin but only 4 (IL13RA2, 1L1B, CD86 and PAD14) were altered in a consistent manner across at least 5 cell strains though not by SLR ≥1.5 in all of them. Of these, IL13RA2, 1L1B and CD86 were up-regulated in 5 cell strains and down-regulated in one while PAD14 exhibited exactly the opposite trend. SECTM1, up-regulated in the presence of BP alone may reflect the effect of BP alone. IL13, an interleukin, has been shown to signal through the interleukin 13-alpha (2) receptor (IL13RA2) to induce TGF-β (1) production and fibrosis (48).
TGF-β has been documented to have diverse cellular roles such as regulation of neuronal function (49), cellular proliferation (50) and modulation of immune responses (51). The expression of IL1B up-regulated by BP was further up-regulated by chlorophyllin in some individuals. CD86 up-regulated in most of the cell strains has been found to have a role in the priming and activation of naive and memory T-cells, respectively (52) and also for enhanced activity of B-cells (53). PADI4 was down-regulated in all individuals on treatment with BP plus chlorophyllin except M99016. It is a gene which is known to encode an enzyme responsible for the conversion of arginine to citrulline and also have a role in granulocyte and macrophage development leading to inflammation. A study showed a significant increase in its expression in many tumor samples, especially in various adenocarcinoma (54).
Nine genes were down-regulated by BP plus chlorophyllin, IL1B and CXCL14 were common to the set of genes up-regulated by BP plus chlorophyllin and down-regulated by BP alone, respectively, indicating inter-individual variations in response profiles among different individuals. IL144, IF127, GIP2 and IFRD1 were altered in a consistent manner across most of the cell strains. IL144, an interferon alpha/beta inducible protein has been shown in a previous study to be elevated in the liver of chimpanzees infected with hepatitis C or hepatitis D, suggesting it to be one of the mediators involved in the antiviral action (55). In another study, IL144 was also shown to play a role with various other genes (interleukins, GADD45A, TNFRSF12A) in candoxin (a neurotoxin from Bungarus candidus venom) induced glial inflammation, DNA-damage and degeneration (56). Interferon alpha inducible protein 127 (IFI27) has been suggested to be a novel biomarker of epithelial proliferation and cancer (57). GIP2 (INSIG1) consistently down-regulated in all the cell strains has previously been suggested to function intracellularly as a ubiquitin homologue and a cytokine that induces production of IFN-gamma to augment NK/lymphokine-activated killer cell proliferation and function, possibly playing a vital role in antiviral response (58). IFRD1 is a gene known to code for a protein which has recently been shown to be involved in muscle differentiation and has a possible role in signal transduction (59).
Several genes altered in this study showed statistically significant associations with the modulation of various other well known pathways (Tables III and IV). Notable among these is an association with the NF-κB pathway. NF-κB has recently emerged as an important player in carcinogenesis, having a role in the modulation of cellular proliferation, apoptosis, cell signaling, angiogenesis and differentiation. Its down-regulation has been implicated in the suppression of cellular and induction of apoptosis. Studies have also revealed NF-κB to be a chemopreventive target for several classes of chemopreventive agents (60, 61). Chlorophyllin, in particular, has been shown to mitigate the activation of NF-κB and also suppress its binding to a cognate recognition motif (62, 63). This points to a possibility of some degree of commonality in the mechanism of immunomodulation with those of other chemopreventive agents.
Though some genes had good correlation of FC as measured on GC versus RT-PCR, the presence of only a moderate to poor correlation for some genes may be due to differences in the precise regions being probed by the two methods, greater sensitivity of RT-PCR over GC and also lack of consistency in expression patterns and levels among all the 6 cell strains. Though differences in the region of the same transcript being probed should typically be expected to yield similar expression patterns, possibility of polymorphisms or transcript variants for the genes in question could alter their expression profiles. Additionally, most probes imprinted on Affymetrix arrays have a 3′ bias, with each gene being represented by 11-20 probes. Given the fact that cRNA is fragmented prior to hybridization on to the arrays it may also be possible that for some genes amidst the overall signal, a greater part of the signal may be generated from certain particular probes whereas these may not be the exact regions being probed by RT-PCR leading to a poor correlation. Moreover, a low level of expression of a particular gene in only some individuals may cause GC to ‘miss’ it or at most exhibit its expression at an extremely low level, whereas the greater sensitivity of RT-PCR may be able to more realistically track its expression giving a higher FC compared to GC, culminating in a poor correlation. However, GC could be an effective technique to identify interesting candidate biomarkers for further validation.
Given that several genes were altered by the presence of BP and chlorophyllin together in treatment T3, it is probable that a synergistic interaction was triggered by the two coming together with the possibility of some of these changes being caused by chlorophyllin alone. These genes in turn are found to interact with each other and various other processes and entities in the cell thereby altering cell signaling. However, there seems to be considerable inter-individual differences in the pattern and extent of modulation, with most genes being differentially modulated in different individuals, with some individuals likely to benefit more from a favorable regulatory effect of chlorophyllin and others an adverse one. For example, M99016 seems to exhibit a slightly different response profile compared to the other cell strains. All genes up-regulated by treatment T2 or T3 in at least one of the other 5 NHMEC lines were all consistently up-regulated in M99016. Similarly, most genes down-regulated by T2 or T3 in at least one of the other 5 NHMEC lines were also almost consistently down-regulated in M99016. Similarly, M98035, M98015, M98025 and M99025 exhibited an almost consistent trend in down-regulation of most genes on treatment with T3 though not with other comparisons. In spite of these differences in individual responses, there appeared to be some genes consistently altered, may be to different extents, but in a predictable direction. These could become potential biomarkers upon further validation. Some of these genes may include IL1B, SECTM1 and CXCL14 for BP exposure and IL1RN, CD86, IF144 and GIP2 for intervention of BP mediated damage by chlorophyllin.
This was a pilot study intended to examine clusters of immune response genes altered on exposure to a prototypical chemical carcinogen, BP, in the presence or absence of the chemopreventive agent chlorophyllin, so as to generate a putative suite of immune response biomarkers. It is acknowledged that before meaningful interpretation of biomarkers of gene expression can be made in the occupational setting, the question of interindividual variation, as it pertains to temporal patterns of gene expression and tests of biological effects should be addressed. Since no data on BP-DNA adduct formation or any other phenotypic anchors was generated, it could be argued that the changes observed could represent metabolic effects to toxic intermediates of BP or other unknown donor factors. However, given the duration of these cells in culture (P6) ‘residual donor signatures’, if any, as a contributing factor to the observed changes could be weak. Additionally, though not shown in this study, these cells have been found to form adducts on BP exposure (unpublished data) thereby suggesting that at least some of these changes may be due to metabolism of the parent compound to toxic intermediates.
In summary, this study has revealed biomarkers of polycyclic aromatic hydrocarbon exposure for further consideration. In addition, it has shown potential modulation of some of the intrinsic immune responses by intervention agents such as chlorophyllin. However, differences in response profiles due to interindividual differences need to be taken into account when designing intervention strategies.
Acknowledgements
We would like to thank Dr. Suresh Jhanwar for critically reviewing this manuscript. This research is published with the approval of the Director of the West Virginia University Agriculture, Forestry and Consumer Sciences Experimental Station as Scientific Paper No. 3022.
Footnotes
-
Disclaimer: The findings and conclusions in this report are those of the Authors and do not necessarily represent the views of the National Institute for Occupational Safety and Health.
- Received September 30, 2008.
- Revision received December 3, 2008.
- Accepted December 10, 2008.
- Copyright© 2009 International Institute of Anticaner Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}