


Abstract
Background/Aim: Methionine addiction is a general and fundamental hallmark of cancer cells, termed the Hoffman effect. Previously Vanhamme and Szpirer showed that methionine addiction could be induced by transfection of the activated HRAS1 gene to a normal cell line. In the present study, we investigated the role of the c-MYC oncogene in methionine addiction of cancer, by comparison of c-Myc expression and malignancy of methionine-addicted osteosarcoma cells and rare methionine-independent revertants, derived from the methionine-addicted cells. Materials and Methods: Methionine-independent revertant 143B osteosarcoma cells (143B-R) were derived from methionine-addicted parental 143B osteosarcoma cells (143B-P), by continuous culture in medium depleted of methionine by recombinant methioninase. To compare in vitro malignancy of methionine-addicted parental cells and methionine-independent revertant cells, the following experiments were performed: for 143B-P and 143B-R cells, cell proliferation capacity was measured with a cell-counting assay, and colony-formation capacity was determined on plastic and in soft agar, all in methionine-containing Dulbecco’s Modified Eagle’s Medium (DMEM). Tumor growth was measured in orthotopic xenograft nude-mouse models, to compare in vivo malignancy of 143B-P and 143B-R cells. c-MYC expression was examined with western immunoblotting and compared in 143B-P and 143B-R cells. Results: 143B-R cells had reduced cell proliferation capacity, compared to 143B-P cells, in methionine-containing medium (p=0.003). 143B-R cells had reduced colony formation capacity on plastic (p=0.003) and in soft agar, compared to 143B-P cells in methionine-containing medium. 143B-R cells had reduced tumor growth in orthotopic xenograft nude-mouse models, compared to 143B-P cells, (p=0.002). These results demonstrate that 143B-R methionine-independent revertant cells lost malignancy. Expression of c-MYC was reduced in 143B-R methionine-independent revertant osteosarcoma cells, compared to 143B-P cells, (p=0.0007). Conclusion: The present study demonstrated that c-MYC expression is linked to malignancy and methionine addiction of cancer cells. The present study on c-MYC, and the previous study on HRAS1, suggest that oncogenes may play a role in methionine addiction, which is a hallmark of all cancers, as well as in malignancy.
- Oncogene
- c-MYC
- methionine addiction
- Hoffman effect
- osteosarcoma
- malignancy
- methionine-independent revertant
- methionine metabolism
- reprograming
Methionine addiction is a general and fundamental hallmark of cancer cells, which was discovered by one of us (RMH) almost a half-century ago (1-6). Methionine addiction, which includes folate addiction (3), is termed the Hoffman effect (2). It appears all cancer types are addicted to methionine (7-12). Methionine addiction is due to overuse of methionine for transmethylation reactions (7, 9-13).
Almost 40 years ago, Vanhamme and Szpirer showed that the transfected mutant HRAS1 oncogene could induce methionine addiction in a non-malignant cell line (14). Since then, the role of other oncogenes in methionine addiction has not been investigated. It has been previously reported that methionine-independent revertant cells, which were selected from methionine-addicted parental cancer cells by continuous culture in low-methionine medium, have reduced malignancy (8, 10-12, 15-17), in part due to changes in methylation of histone-H3 lysine marks (7, 9-12).
We subsequently reported that reversion of methionine-addicted osteosarcoma cells to methionine-independence reduced malignancy, along with reduced epithelial-to-mesenchymal transition (12).
In the present report, by comparing methionine-addicted osteosarcoma cells and methionine-independent revertant cells derived from them, we showed that malignancy of osteosarcoma cells is also linked to methionine addiction via reduction of c-MYC expression. The present results further support the concept that malignancy of cancer cells is linked to methionine addition, via oncogenes.
Materials and Methods
Cell culture. The 143B human osteosarcoma cell line was purchased from the American Type Culture Collection (ATCC) (Manassas, VA, USA), and cultured in Dulbecco’s Modified Eagle Medium (DMEM) (Corning Inc., Corning, NY, USA), with one IU/ml penicillin/streptomycin and 10% fetal bovine serum (FBS).
Animals. Female athymic nu/nu nude mice (4-6 weeks old) (AntiCancer, Inc., San Diego, CA, USA) were used in the present study. All mice were bred and maintained as previously reported (12), under Assurance Number A3873-1, with an Institutional Animal Care and Use Committee (IACUC) protocol, following the National Institutes of Health (NIH) Guide for the Care and Use of Animals. For all surgical procedures, a cocktail of anesthetics and analgesics (ketamine: 20 mg/kg, xylazine: 15.2 mg/kg, acepromazine maleate: 0.48 mg/kg) was used, in order to minimize animal distress.
Recombinant methioninase production. Recombinant methioninase (rMETase) was produced by a modified previously-reported procedure (18).
Selection for methionine-independent revertant osteosarcoma cells. Methionine-independent revertant 143B osteosarcoma cells (143B-R) were selected as previously reported (12). Briefly, methionine-addicted parental 143B osteosarcoma cells (143B-P) were cultured with increasing concentrations of rMETase (0.75-1.6 U/ml) for 3 weeks, and they were then cultured in normal medium without rMETase for 3 weeks, followed by passage. This procedure was repeated 4 times.
Cell-proliferation assay. 143B-P and 143B-R cells (5.0×103 cells/well) were cultured in 6-well plates, in DMEM and incubated for the following durations: 24 h; 48 h; 72 h; 96 h at 37°C. After each incubation period, cells were detached by trypsin EDTA (Corning Inc.), and the number of cells was counted with a hemocytometer (Hausser Scientific Co., Horsham, PA, USA). Experiments were performed twice in triplicate.
Clonogenic assay on plastic. 143B-P and 143B-R cells (5.0×103 cells/well) were cultured in 6-well plates, in DMEM and incubated for one week at 37°C. After the incubation period, cells were stained with 0.5% crystal violet, and the area of the colonies was measured, as previously described (19). Experiments were performed twice in triplicate.
Clonogenic assay in soft agar. 143B-P and 143B-R cells (5.0×103 cells/well) were cultured in 6-well plates in DMEM, in 0.3% agar, over a layer of 0.5% agar underlay, for 2 weeks at 37°C. After the incubation period, cells were stained with 0.05% crystal violet, and the number of colonies was counted, as previously described (19). Experiments were performed twice in triplicate.
Osteosarcoma orthotopic xenograft mouse model. 143B-P and 143B-R cells [2.5×105 cells/12.5 μl PBS and 12.5 μl Matrigel (Corning Inc.)] were injected into the left proximal part of the tibia bone, of ten mice each. The procedure was performed as previously described (12). Tumor size was measured once a week with calipers, with the following formula: tumor volume = length (mm) × width (mm) × width (mm) × 1/2. All mice were sacrificed four weeks after cell injection.
Western immunoblotting. Protein extraction of 143B-P and 143B-R cells and Western immunoblotting for total proteins was performed as follows: Protein extraction was performed using RIPA Lysis and Extraction Buffer (Thermo Fisher Scientific, Waltham, MA, USA) with 1% Halt Protease Inhibitor Cocktail (Thermo Fisher Scientific). Protein samples were loaded onto 10% SDS-PAGE gels for electrophoresis, and then transferred to 0.45 μm polyvinylidene difluoride (PVDF) membranes (GE Healthcare, Chicago, IL, USA). The membranes were blocked with Bullet Blocking One for Western Blotting (Nakalai Tesque, Inc., Kyoto, Japan). Anti-c-MYC antibody (1:2,000, #10828-1-AP, Proteintech, Rosemont, IL, USA) and anti-GAPDH antibody (1:50,000, 60004-1-Ig, Proteintech) were used. GAPDH was used as an internal loading control. Horseradish-peroxidase-conjugated anti-mouse IgG (1:5,000, #SA00001-1, Proteintech) for GAPDH and horseradish-peroxidase-conjugated anti-rabbit IgG (1:5,000, #SA00001-2, Proteintech) for cMYC were used as the secondary antibodies. The signals were detected with the UVP ChemStudio (Analytik Jena, Upland, CA, USA) with Clarity Western ECL Substrate (Bio-Rad Laboratories, Hercules, CA, USA). The experiments were performed in quadruplicate.
Statistical analysis. Welch’s t-test was performed to statistically compare the means of two groups, including 143B-P and 143B-R, with Microsoft Excel for Mac ver. 16.16.27 (Microsoft, Redmond, WA, USA). Quantification of protein expression was performed with ImageJ ver. 1.53a (National Institutes of Health, Bethesda, MD, USA). Bar graphs show the mean and error bars show standard deviation of the mean. A probability value ≤0.05 was defined as statistically significant.
Results
Methionine-independent revertant osteosarcoma cells had reduced cell proliferation capacity in methionine-containing medium. 143B-R cells showed significantly reduced cell-proliferation capacity, compared to 143B-P cells (p=0.003) (Figure 1).
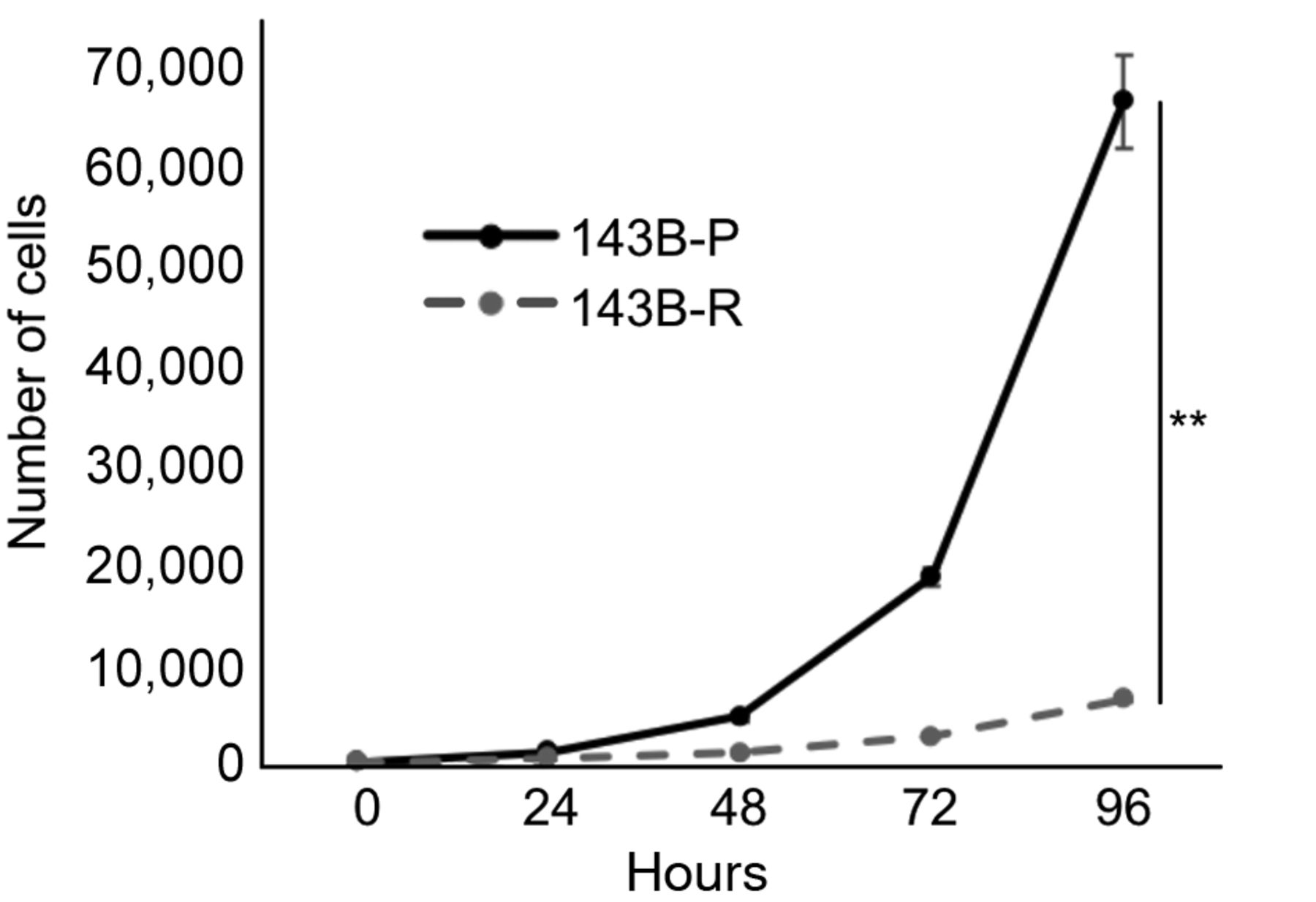
Time-course of proliferation for 143B-P and 143B-R cells in methionine-containing medium. 143B-P: Methionine-addicted parental 143B osteosarcoma cells, 143B-R: methionine-independent 143B osteosarcoma cells. **p<0.01.
Methionine-independent revertant osteosarcoma cells had reduced colony formation capacity in methionine-containing medium. 143B-R cells showed significantly reduced colony formation capacity on plastic (p=0.003) (Figure 2) and in soft agar (Figure 3), compared to 143B-P cells.

Clonogenic assay on plastic for 143B-P and 143B-R cells. (A) Representative photographs of colonies after one-week growth. Scale bar: 10 mm. (B) Bar graphs corresponding to photographs in (A). 143B-P: Methionine-addicted parental osteosarcoma cells, 143B-R: methionine-independent osteosarcoma cells. **p<0.01.

Clonogenic assay in soft agar for 143B-P and 143B-R cells after two-weeks’ growth. Representative photographs (top) and photomicrographs (bottom). Scale bar in photographs: 10 mm. Scale bar in photomicrographs: 250 μm. 143B-P: Methionine-addicted parental 143B osteosarcoma cells, 143B-R: methionine-independent 143B osteosarcoma cells.
Methionine-independent revertant osteosarcoma cells had reduced tumor growth in orthotopic xenograft mouse models. 143B-R cells formed tumors in 9 out of 10 mice, and 143B-P cells formed tumors in 8 out of 10 mice (Figure 4A). However, the tumor size of 143B-R was 10-fold smaller than that of 143B-P (p=0.002) (Figure 4B and C).

Tumor growth capacity in orthotopic nude-mouse models. (A) Tumor tissues of orthotopic xenograft mouse models of 143B-P and 143B-R cells 4 weeks after cell injection. Dotted lines show the margin of the tumor tissue. (B) Tumor growth curve of 143B-P and 143B-R cells. (C) Mean tumor volume of 143B-P and 143B-R cells 4 weeks after cell injection. Scale bar: 20 mm. 143B-P: Methionine-addicted parental osteosarcoma cells, 143B-R: methionine-independent osteosarcoma cells. **p<0.01.
Expression of c-MYC was reduced in methionine-independent revertant osteosarcoma cells. Methionine-independent 143B-R revertant cells had 0.68-expression of c-MYC, compared to methionine-addicted 143B-P parent cells (p=0.0007) (Figure 5).
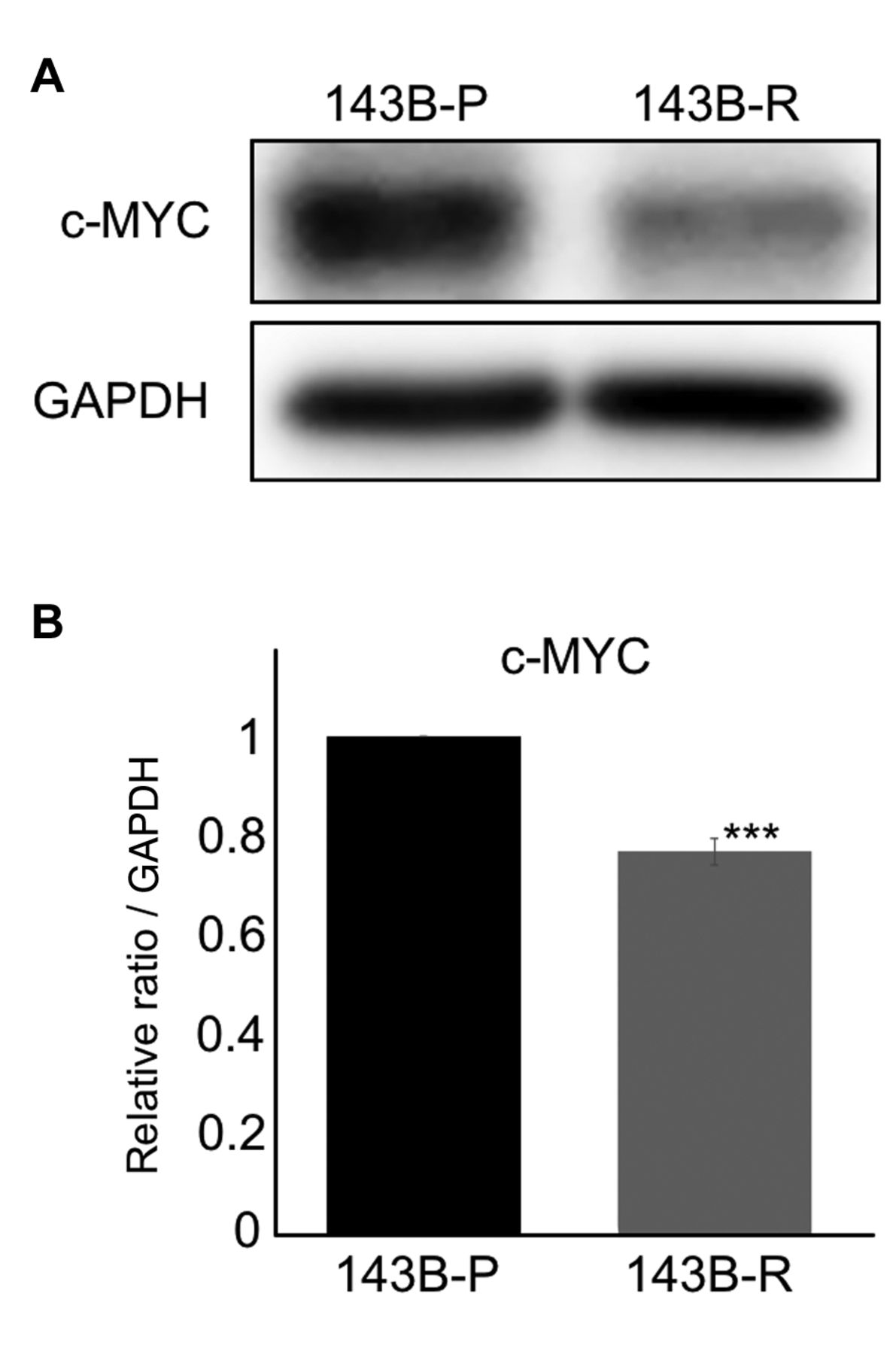
c-MYC expression. (A) Expression of c-MYC in 143B-P and 143B-R cells, with immunoblotting. (B) Bar graph quantifying c-MYC expression from the Western blot in (A) cells. 143B-P: Methionine-addicted parental osteosarcoma cells, 143B-R: methionine-independent osteosarcoma cells. ***p=0.0007.
Discussion
The present study showed that methionine-independent revertant osteosarcoma cells had reduced cell-proliferation capacity, colony-formation capacity, and tumor-growth capacity, compared to their methionine-addicted parental cells, indicating their reduction of malignancy, which was linked to reduced expression of the c-MYC oncogene in the revertant cells compared to the methionine-addicted parental cells. The results of the present study are consistent with our previous reports, in which it was shown that methionine-independent revertant cancer cells had reduced malignancy, evidenced by cell migration and invasion capacity in vitro, and reduced tumor growth and metastatic capacity in vivo, compared to their methionine-addicted parental cells, (8, 10-12, 15-17). The present and previous results strongly support the concept that malignancy of cancer cells is linked to methionine addiction.
In the present study, methionine-independent revertant osteosarcoma cells had reduced c-MYC expression. c-MYC is one of the most commonly activated oncogenes in human cancers, including osteosarcoma. c-MYC contributes to cell proliferation and growth, and c-MYC expression has been reported to be associated with worse prognosis in osteosarcoma (20-24). In the present study, cell proliferation and colony-formation were reduced in vitro and tumor size of the methionine-independent revertant osteosarcoma was significantly smaller than that of the methionine-addicted parental cells. These results indicate that the methionine-independent revertant osteosarcoma cells have reduced their malignancy, at least in part, by reduced expression of c-MYC.
The present results, which show that c-MYC expression is related to methionine addiction, extend the concept that oncogenes play a role in methionine addiction, first observed by Vanhamme and Szpirer, who showed that transfected HRAS1 induced methionine addiction in a non-malignant cell line (14).
Future studies will focus on further understanding the relationship of c-MYC expression and methionine addiction, and whether other oncogenes are involved in methionine addiction, which involves a reprograming of methionine metabolism (1-6, 10-14, 25-31). Metabolic addiction can also include oxygen addiction (32), as well as methionine and folate addiction (1-6) and is an important area of future investigation.
Acknowledgements
This article is dedicated to the memory of A. R. Moossa, MD, Sun Lee, MD, Professor Li Jiaxi, Masaki Kitajima, MD, Joseph R. Bertino, MD, Jack Geller, MD, and Shigeo Yagi, PhD.
Footnotes
Conflicts of Interest
The Authors declare that there are no conflicts of interest in relation to this study.
Authors’ Contributions
YA, YT and RMH were involved in study conception and design. YA, YK, NM and KO were involved in acquisition of data. YA, YT, YK, NM, KO, and RMH analyzed and interpreted data. YA, YT and RMH wrote the manuscript. All Authors reviewed and approved the manuscript.
Funding
This study was funded in part by the Robert M. Hoffman Foundation for Cancer Research.
- Received January 16, 2023.
- Revision received January 30, 2023.
- Accepted February 7, 2023.
- Copyright © 2023, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).
References
In this issue





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Reduction of Tumor Biomarkers from very High to Normal and Extensive Metastatic Lesions to Undetectability in a Patient With Stage IV HER2-positive Breast Cancer Treated With Low-dose Trastuzumab Deruxtecan in Combination With Oral Recombinant Methioninase and a Low-methionine Diet
- The Combination of Methioninase and Ethionine Exploits Methionine Addiction to Selectively Eradicate Osteosarcoma Cells and Not Normal Cells and Synergistically Down-regulates the Expression of C-MYC