


Abstract
Background/Aim: We previously described four different vascular patterns (reticular, diffuse, fasciculate, and trabecular) in renal cell carcinoma (RCC) suggesting an early and heterogeneous acquisition of perivascular cells most probably due to a particular PDGF pathway gene expression profile. The aim of the study was to study PDGF pathway gene expression profiles, separately for each vascular pattern. Materials and Methods: TaqMan assay for the PDGF pathway was performed on twelve cases of ccRCC previously evaluated by histopathology, immunohistochemistry, and RNAscope. Gene expression profile was correlated with grade, invasion, vascular patterns, and VEGF. Results: PIK3C3 and SLC9A3 genes were overexpressed in all vascular patterns, but they were significantly correlated with high VEGF mRNA in the reticular and diffuse pattern. STAT1, JAK2, SHC2, SRF and CHUK (IKK) were exclusively overexpressed in cases with diffuse vascular pattern. SLC9A3, CHUK and STAT3 were overexpressed in G2 tumors. Conclusion: Three ccRCC subgroups were defined: 1) PIK3C3 (VSP34)/SLC9A3 which may be proper for anti PIK3C3 inhibitors; 2) VEGFhigh subgroup where association of anti VEGF may be a benefit and 3) JAK2/STAT1 subgroup, potentially being eligible for anti JAK/STAT therapy associated with IKK inhibitors.
Renal cell carcinoma (RCC) represents about 2.2% of total diagnosed cancers worldwide with an estimated number of newly diagnosed patients of 400,000 approximately (1, 2). Fifteen percent of newly diagnosed cancers are diagnosed in the primary metastatic RCC (mRCC) stage of disease but more than 30% of cases which had no metastases at the time of diagnosis become metastatic in a variable period of time of 4 to 8 years from the primary diagnosis despite application of novel and targeted therapies (3). Renal cell carcinoma clear cell type (ccRCC) arises from the proximal tubules of the kidney parenchyma and is the most common subtype of RCC, representing 80% of RCCs (4). ccRCC is a complex disease with a silent development and an unexpected behavior having a highly heterogeneous response to therapy (5). Normal kidney and malignant tissue arising from it have a special and unique microenvironment (6) and this, may be influence the resistance to therapy which appear early during the treatment (7) despite of modern and targeted therapies developed in the last years (8, 9).
Inside renal carcinoma microenvironment heterogeneity, two main components seem to be responsible for the rapid development of therapy resistance: tumor blood vessels and immunogenic profile (10, 11). Vascular and immunogenic compartments of ccRCC are the most “targeted” compartments in ccRCC, antiangiogenic therapies and immune-checkpoints inhibitors being, currently, the gold standard for the treatment of renal cancer.
The heterogeneity of vascular compartment of ccRCC tumor microenvironment is easy to observe also by studying the morphology and immunophenotype of ccRCC tumor vessels. Previously, our team described 4 vascular patterns based on their morphology and cellular components (endothelial cells versus perivascular cells) (12). An unusual tendency to a heterogeneous acquisition of perivascular cells (as time and morphology) was highlighted for tumor blood vessels from ccRCC but also a different arrangement of these vessels inside the tumor. Perivascular cells heterogeneity distribution (assessed by a double immunostaining of tumor blood vessels with CD34/Smooth Muscle Actin antibodies) was the hallmark of the 4 groups of tumor blood vessels from ccRCC defined by our team and was strongly correlated with tumor invasion (12).
Later, in a paper published by Ruis Sauri and colleagues (13) the authors confirmed the presence of the 4 vascular patterns and also reported that these patterns are strongly correlated with ISUP tumor grade (13).
Because of the discrepancies between the high number and unusual morphology of blood vessels from ccRCC (despite of a low endothelial cells proliferation rate) and highly maturated state of these tumor blood vessels, we considered that the study of PDGF-B/PDGFRβ axis is mandatory in ccRCC and correlated with the inhibitory isoform of vascular endothelial growth factor (VEGF), VEGF 165b. In a paper published in 2016 our group reported a significant correlation between VEGF165b and PDGF-B/PDGFRβ ratio (14), this being correlated with vascular patterns previously described. This correlation, exclusively found in the reticular pattern, supports the fact that concomitant inhibition of endothelial cells by VEGF165b and rapid perivascular cell recruitment driven by the PDGF-B/PDGFRβ axis, may define several types of heterogeneous tumor vasculature in ccRCC.
It is well known that any morphology and immunophenotype heterogeneity is supported by a specific gene expression profile (15). Gene expression profiling is a feasible tool to define molecular classes inside various malignancies and based on these stratify prognostic and therapeutic subgroups which may better define the heterogeneous response to therapy, prognosis and survival. The last papers in the field of ccRCC were focused in investigating molecular subgroups of ccRCC (15, 16).
Our previous findings on ccRCC vascular heterogeneity required validation by more specific methods. Tumor blood vessel maturation is given by acquisition of vascular smooth muscle cells driven by PDGF-B/PDGFRβ axis. We aimed to study gene expression profile of the PDGF pathway by using TaqMan™ Array Human PDGF Pathway and correlate the findings with vascular patterns previously described in ccRCC. Further, we aimed to identify potential subgroups of ccRCC able to give a better overview of ccRCC carcinomas with possible different prognostic and therapeutic impact.
Materials and Methods
Case selection. We previously selected 50 retrospective cases of formalin fixed paraffin embedded (FFPE) kidney cancer randomly selected from the archive of Pathology Department of Clinical County Hospital Timişoara, Romania. Biopsies were harvested between 2005 and 2015, fixed in 10% buffered formalin and paraffin embedded following routine protocol. Three independent experienced histopathologists certified microscopic diagnosis of renal cancer, clear cell type. On these cases we previously performed immunohistochemistry, RNAscope and in situ hybridisation focusing on the study of VEGF, PDGF and CD34/SMAct co-expression and we defined four vascular patterns in ccRCC (12). Twelve cases of ccRCC (selected based on vascular patterns previously described by our team (12) were prepared for TaqMan Assay technique. The TaqMan® Array Human PDGF Pathway 96-well plate contains 92 assays to PDGF Pathway associated genes and 4 assays to endogenous control genes. All the experiments were performed in triplicate.
Tissue samples used for RNA extraction were collected by automated image guided coring with Tissue Micro Array technology. Microscopic slides were scanned by using Panoramic Desk Slide Scanner (3Dhistech, Budapest, Hungary) and areas of interest were marked by using automated software of TMA Grand Master (3Dhistech) overlapped to correspondent donor paraffin blocks loaded. Briefly, all 12 donor blocks and correspondent 12 PCR tubes were uploaded into automated TMA web-based digital platform. A 2.0 mm diameter core punch instrument was selected and used to pick up tissue fragments. The specimens are viewed and annotated (marked) using 2.0 mm diameter tool, multiple times using specific color to distinguish tissue areas. Four cores of 2 mm diameter each were collected into 0.2ml PCR tubes from each FFPE block followed by RNA extraction using a semi-automated method described below.
RNA extraction by semi-automated magnetic beads-based technology and cDNA synthesis. Dewaxing of FFPE tissue cores was performed by filling PCR tubes containing material with xylene and incubating them for 60 minutes at room temperature for cores dewaxing. Xylene was discarded and RNA extraction was started by using King Fisher Pure RNA Tissue Kit (Thermo Scientific, Waltham, MA, USA). Reagent preparation was done according to the protocol provided by the manufacturer and loaded into Kingfisher 96 Deep Well Plates based on the predesigned template included in the protocol. RNA extraction continued by loading the plates into King Fisher Duo Prime automated System (Thermo Scientific, Santa Clara, CA, USA) and by choosing Pure RNA Tissue Duo program as running protocol. The RNA total amount was checked using a Qubit™ 3.0 fluorometer (Thermo Scientific, Santa Clara, USA) by preparing the preliminary samples with the Qubit™ RNA HS Analysis Kit (code Q32852, Thermo Scientific). cDNA synthesis from the extracted RNA was performed by using the Applied Biosystem High-capacity RNA-to-cDNA kit™ (Fischer Scientific, Loughborough, UK) following the protocol provided by the manufacturer. Two micrograms of total RNA were used per 20-μl reaction. The TaqMan Array plates were prepared by loading 20 μl of cDNA sample prepared by using 10 μl of nuclease-free water and 10 μl of Master Mix (2x) on each well. The TaqMan Array Human PDGF plates were subjected of RT-PCR technique performed on the 7500 Fast Dx Real-Time PCR Instrument (Agilent, Santa Clara, CA, USA).
Immunohistochemistry. The cases for TaqMan Array were selected based on previous assessments of immunohistochemical stained slides with a double-stain method for endothelial cells (monoclonal mouse anti-human CD34, clone QBEnd 10, Novocastra, Newcastle upon Tyne, UK) and for perivascular cells (monoclonal mouse anti-human Smooth Muscle Actin antibody, clone 1A4, Novocastra). Each step of immunohistochemical procedure was done in an automated manner by using software of Bond Max Autostainer (Leica MicroSystems, Deer Park, IL, USA). Thirty minutes incubation time with primary antibodies was followed by the use of Bond Refine Detection System Dual Color (Leica MicroSystems). Based on the CD34/SMAct coexpression we selected cases according to our team classification reported by Ferician et al. (15) as we include all four types of vascular patterns previously described.
Data analysis. The microscopic assessment of CD34/SMAct was performed by three independent pathologists using AxioZoom Zeiss Microscope (Zeiss, Oberkochen, Germany). Interaction between CD34 positive endothelial cells and SMAct positive perivascular cells was the main criterion used to select cases for TaqMan Array. TaqMan assay results were evaluated by using Data Assist V.3.01 software (Thermo Fisher Scientific, Waltham, MA, USA). Heat Map and volcano plots were automatically generated from this analysis.
Results
We previously characterized all 12 cases of ccRCC included in the present study regarding tumor grade, vascular pattern, the presence of CD34/SMAct immunoexpression, and assessment of tumor blood vessels stabilization (a parameter which make ccRCC less responsive to antiangiogenic and/or antivascular therapy). Initial microscopic assessment of each ccRCC specimen included the evaluation of tumor grade. Eight cases were scored as having tumor grade 2 and 4 cases with tumor grade 3. We detected a high angiogenic process inside tumor mass (Figure 1) with a high density of tumor blood vessels being in all stages of angiogenic process (buds, cords, tube-like structures, perfused vessels). Despite an active angiogenic morphological appearance, even for angiogenic stages where SMAct-positive perivascular cells should be absent (buds and cords stages), we observed SMAct-positive perivascular cells with a heterogeneous distribution.

Intense tumor angiogenic process inside renal cell carcinoma clear cell type (ccRCC). Note tumor blood vessels heterogeneity inside the same microscopic field, containing vascular buds (sign of an intense endothelial cells activation), cords (suggesting an active endothelial cells migration and small vascular structures with lumen (as already functional tumor blood vessels). Yellow arrows indicate early presence of smooth muscle actin (SMAct) positive perivascular cells which usually appear only in stabilisation stage of tumor blood vessels. As herein shown, perivascular cells are partially attached to the outer part of the vessel, some of them having unusual extensions in between two newly formed blood vessels (yellow arrows, lower part of the image).
All cases showed stable tumor blood vessels highlighted by CD34/SMAct immunostaining in the whole area or just in spotted zones of tumor mass (Figure 2a). Most tumor blood vessels showed an oversized perivascular cell network highlighted with smooth muscle actin (SMA) immunostaining (Figure 2b). Also, the arrangement of SMA-positive perivascular cells was not normal (closely attached to the outer circumference of the vessel wall), being partially detached (Figure 2b) or being distributed as bridge-like structures in between small tumor blood vessels loops (Figure 2b).
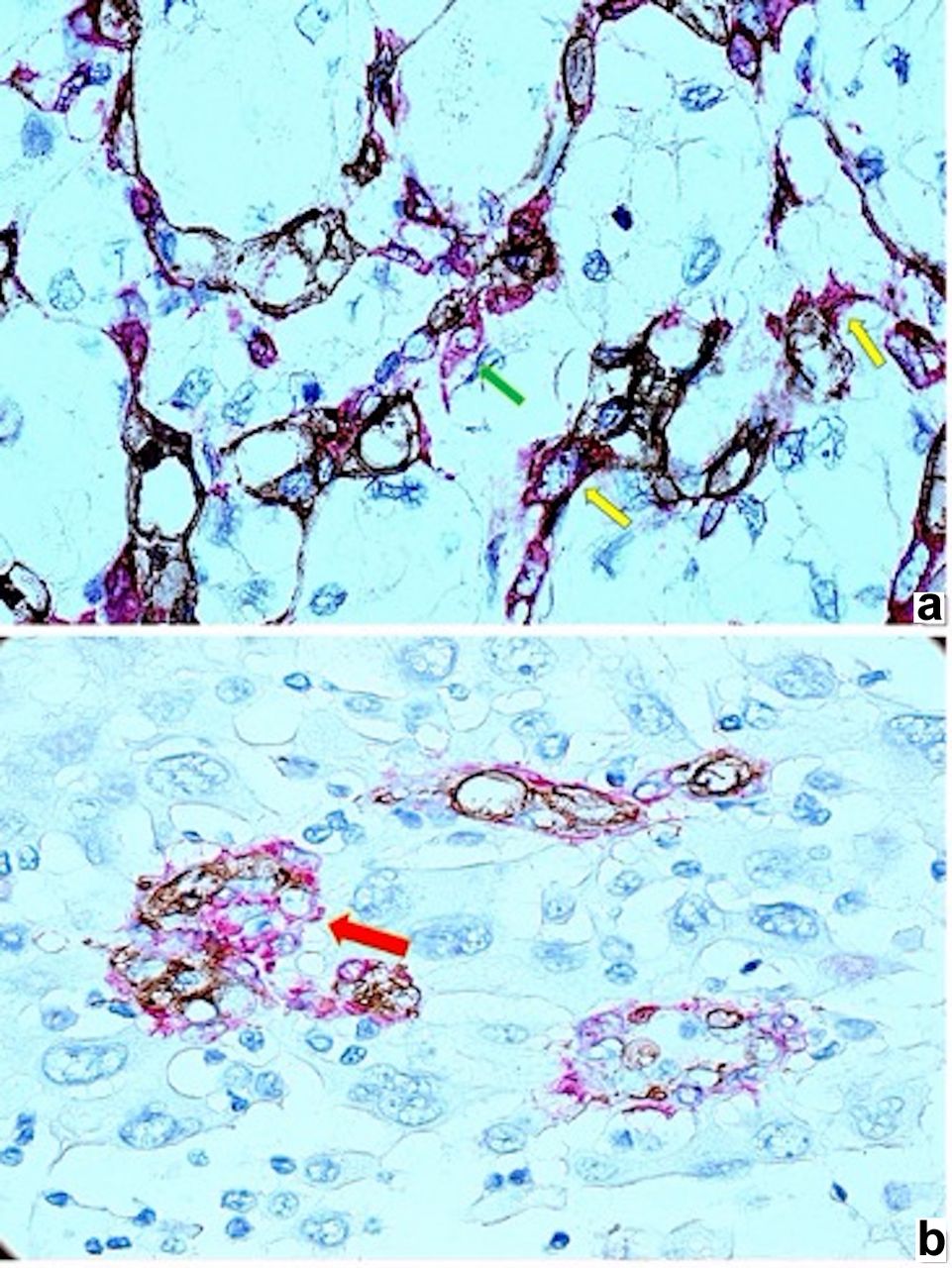
Early presence, particular morphology and distribution of smooth muscle actin (SMAct) positive perivascular cells related to renal cell carcinoma clear cell type (ccRCC) tumor blood vessels. Incomplete attachment of perivascular cells on tumor blood vessels [(a), green arrow, stained in red)] and bridge-like arrangement of smooth muscle actin (SMAct) positive perivascular cells (stained in red) in between tumor vessels [(a), yellow arrows)]. Other tumor areas have glomeruloid like structures [(b), red arrow)] with smooth muscle actin (SMAct) positive perivascular cells surrounding CD34 positive structures without evident lumen (most probably newly formed blood vessels in early stages of angiogenesis).
Based on these preliminary microscopic observations together with a high immunohistochemical expression of PDGF-BB previously reported by our team (17) and given the fact that perivascular cells acquisition is driven by PDGF-B/PDGFRβ pathway, we performed TaqMan Array to highlight the peculiarities of PDGF pathway gene expression to identify potential gene overexpression related to unusual early presence, morphology and distribution of SMAct-positive perivascular cells.
Immunohistochemical evaluation of PDGF-B/PDGFRβ showed that all 12 cases of ccRCC were positive for PDGF-BB, by immunohistochemistry and RNAscope of which 91.6% were confirmed by RT-PCR.
PDGF pathway gene expression profile was evaluated related to our previous results regarding VEGF immunohistochemical and RNAscope expression and also with our previous classification which stratifies the types of tumor blood vessels from ccRCC in 4 categories: reticular, diffuse, fascicular, trabecular.
Based on TaqMan Array results analysis, a PDGF pathway gene expression profile HeatMap has been automated created by Data Assist analysis software. As it has been shown in Figure 3, HeatMap analysis revealed several overexpressed PDGF pathways genes. Genes with the highest overexpression were: PIK3C3 (VsP34), SLC9A3, STAT1, JAK2, SHC2, SRF and CHUK.

Heat Map of platelet derived growth factor (PDGF) pathway automatically generated based on TaqMan Assay analysis. Gene overexpression was highlighted in red, gene down-regulation in green and the lack of expression in black. As shown in the heatmap, overexpressed genes were: PIK3C3 (VsP34), SLC9A3, STAT1, JAK2, SHC2, SRF and CHUK.
Volcano plot analysis was correlated with ccRCC vascular patterns, VEGF immunohistochemical expression and tumor grade.
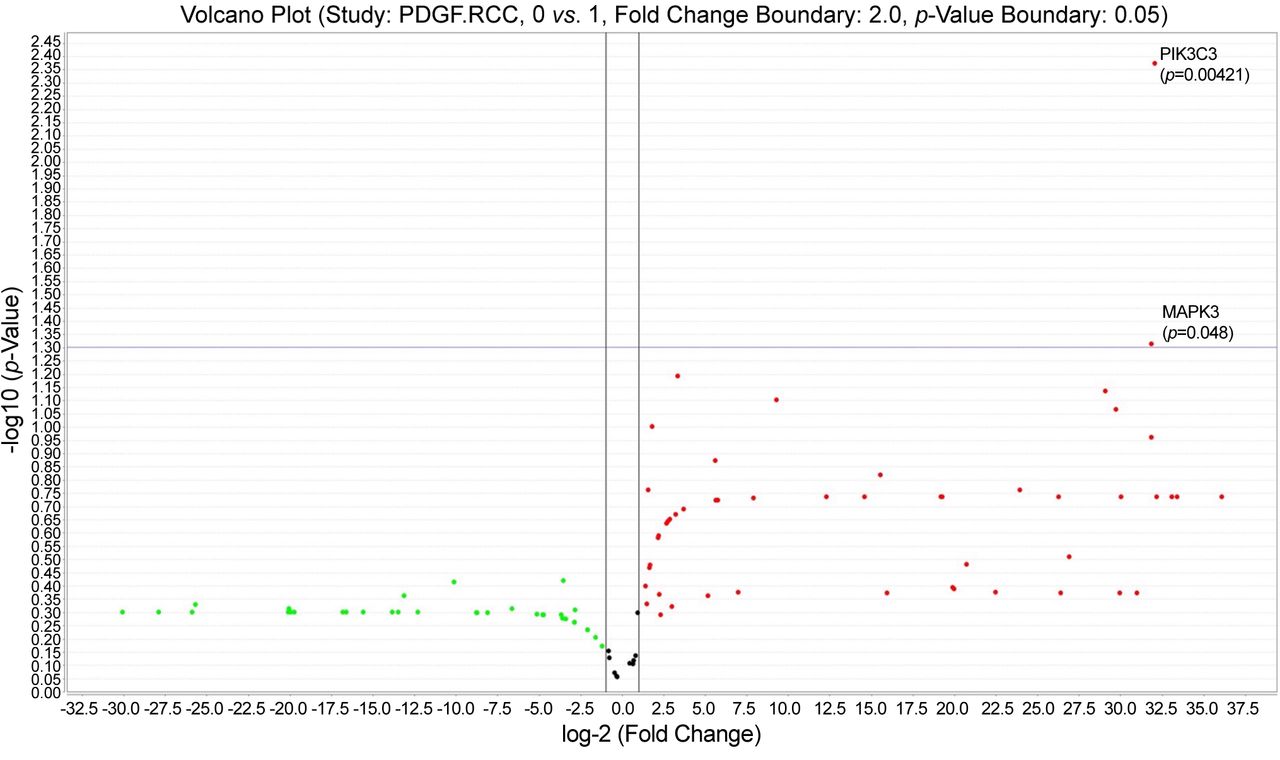
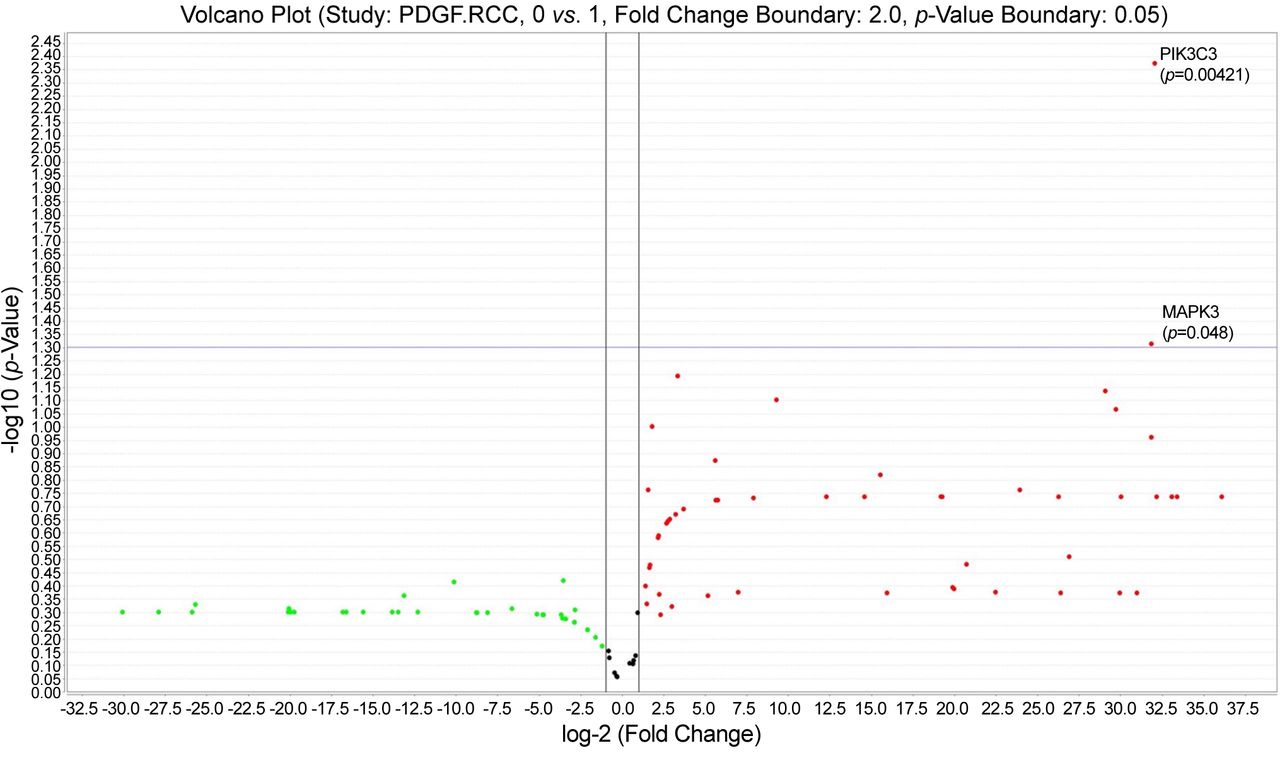
PIK3C3 (VsP34) and SLC9A3 were up-regulated (Figure 4) for both reticular and diffuse vascular patterns ccRCC cases which had a moderate and intense VEGF immunohistochemical expression (VEGF score 2 ore 3) certified by RNAscope in situ hybridization method (Figure 4, inset).

Volcano plot showing statistically significant up-regulated genes: SLC9A3 and PIK3C3, common for reticular and diffuse vascular patterns of ccRCC.
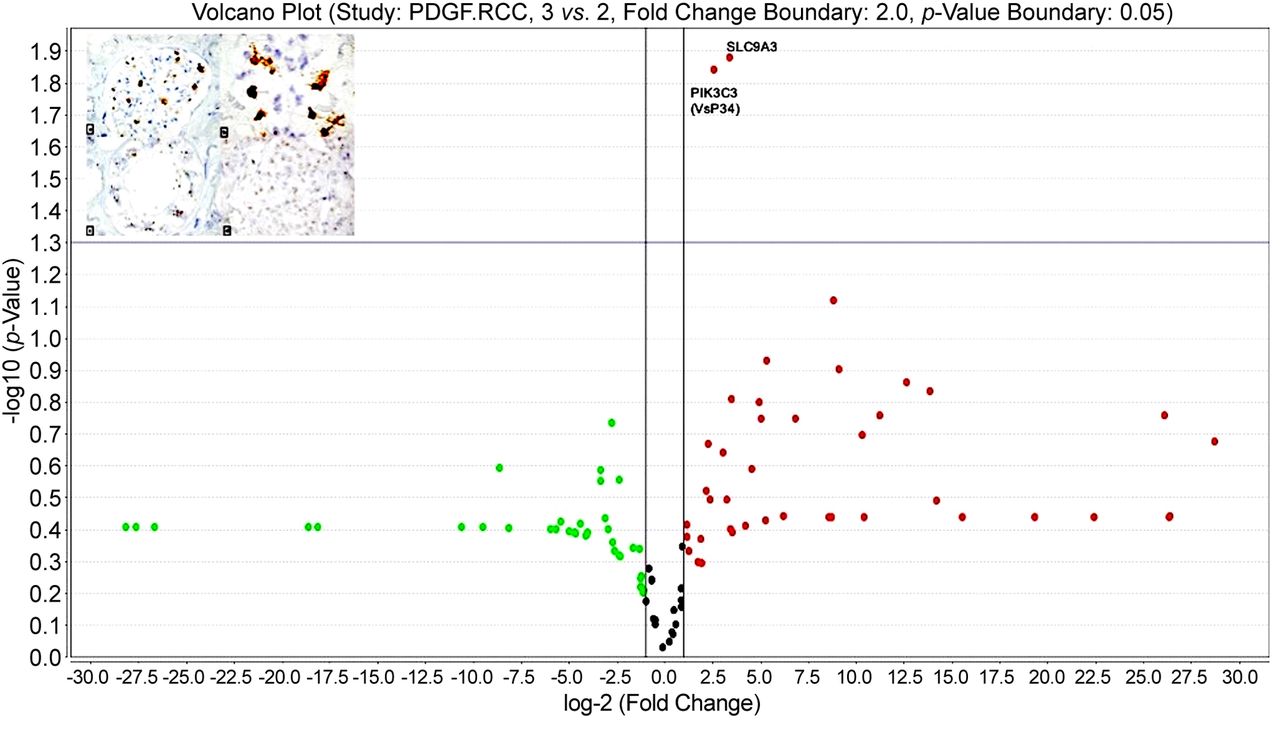
In the reticular vascular pattern ccRCC group, vascular invasion was found in 66.6% out of the total number of cases. For this group, PDGF pathway TaqMan analysis revealed a statistically significant MAP2K2 up-regulation (p=0.004). For the diffuse and trabecular pattern group we detected vascular invasion for 40% of cases. Analysis of PDGF pathway gene expression profile identified other two up-regulated genes specific for this group: PIK3C3 (p=0.004211) and MAPK3 (p=0.048) (Figure 5). MAPK3 was found to be up-regulated also in group with high expression of VEGF inhibitory isoform, VEGF 165b (p=0.0127).
Volcano plot showing gene up-regulation for diffuse vascular pattern cases with invasion.
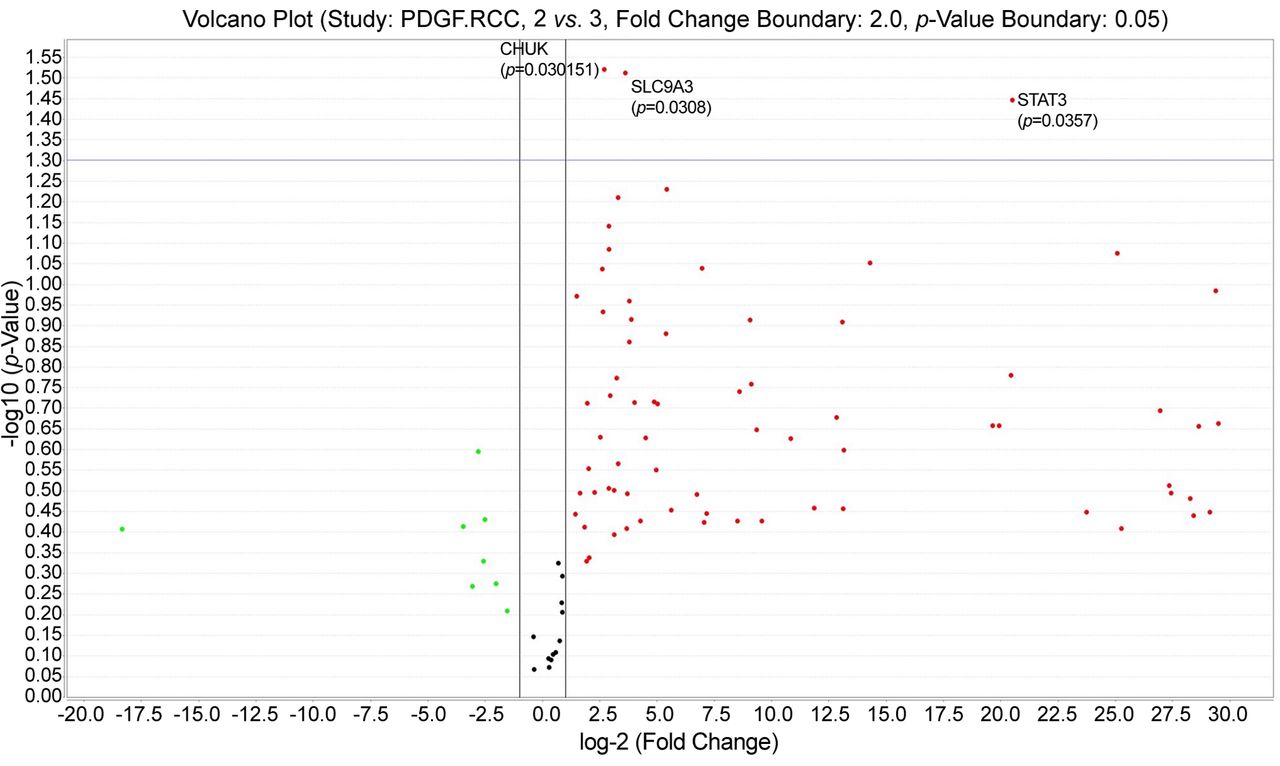
PDGF pathway TaqMan assay was also assessed related to tumour grade. CHUK (p=0.030151), STAT3 (p=0.0357) and SLC9A3 (p=0.0308) genes were statistically significant up-regulated for ccRCC cases having tumor grade 2 (see volcano plot from Figure 6).

Platelet derived growth factor (PDGF) pathway gene expression profile assessment related to tumor grade. Grade 2 was characterized by significant up-regulation for 3 genes compared to other tumor grades.
Five new different genes have been found to be down-regulated in ccRCC cases with reticular pattern (Figure 7) and up-regulated for diffuse pattern exclusively. These genes are: STAT1, JAK2, SHC2, SRF, CHUK.

STAT1, JAK2, SHC2, SRF, CHUK down-regulation for the diffuse and reticular pattern.
Based on gene analysis of the PDGF pathway in ccRCC correlated with VEGF expression and tumor blood vessels types, we defined three subgroups of renal carcinomas which may have three different therapeutic options. The first is the VEGFhigh subgroup showing PIK3C3 (VPS34) and SLC9A3 up-regulation. This group includes both reticular and diffuse vascular patterns and might be eligible to a combined therapeutic option of anti-VEGF agents and PIK3C3 inhibitors. The second group may be defined as VEGFlow/- showing a similar PIK3C3 (VPS34), SLC9A3 up-regulation. For this group, anti-PDGF-PDGFR may be suitable. The third subgroup that we called JAK/STAT subgroup is characterized by overexpression of the 5 genes, suitable for anti JAK/STAT therapy associated with PIK3C3 and IKK inhibitors (Figure 8).

Proposed renal cell carcinoma clear cell type (ccRCC) subgroups based on Platelet Derived Growth Factor (PDGF) pathway assessment correlated to ccRCC tumour vascular pattern.
Discussion
Renal cancer remains one of the most unpredictable malignancies related to its intratumor heterogeneity, invasiveness, metastatic behaviour, response to conventional and targeted therapies (17, 18) as well as to high vascularisation and heterogeneity of tumour blood vessels (12, 19). VHL gene mutation has been reported for several years as one of the main inductors of ccRCC initiation and progression, while other gene expression profile pathways (as angiogenic or lymphangiogenic pathways gene expression profile) being somehow neglected for several years.
Most of ccRCC are characterized by an increased density of intratumor blood vessels at the time of diagnosis. Intratumor blood vessels have a highly heterogeneous morphology previously described and classified by our team in four vascular patterns based on their microscopic morphology, presence and arrangement of perivascular cells (12). Three out of four vascular patterns described before (12) included tumor blood vessels showing a high grade of maturation certified by the presence of SMA-positive perivascular cells. Following our paper related to vascular pattern heterogeneity (12), other reports continued to be published being increasingly focused on ccRCC tumor blood vessels heterogeneity and its potential impact on the development of resistance to targeted therapies (13, 20-22).
Relatively recent data suggested that there is a strong mutual communication in between tumor cells and intratumor blood vessels. The relationship between the tumor microenvironment and ccRCC tumor cell heterogeneity is increasingly reported in the literature both on experimental models, and by gene expression analysis of human tissues derived from patients with renal cancer (23, 24). Intratumor and peritumour microenvironment is highly heterogeneous not only by its cellular components, such as tumor fibroblasts (25) or macrophages (26), but also by its vascularisation (12). Yuan and colleagues, reported in 2016 that highly perfused ccRCC tumor areas, have increased cellularity and increased microvessel density compared to low-perfused areas. This suggests a direct correlation between vascularity and tumor cells in ccRCC (27). Following this report, one of the first to suggest the interrelation between tumor cell behavior and renal tumor vascularization, several studies have confirmed that tumor cell aggressiveness and their response to therapy are closely related to tumor vascularization (27-30). As evidence of the essential involvement of blood vessels in the behavior of tumor cells, most targeted therapies used in the treatment of ccRCC, target tyrosine kinases and their receptors (31, 32). Under these targeted therapies recurrence and metastasis rates decreased significantly but resistance to therapy still developed by incompletely elucidated mechanisms (33, 34).
Despite the above, there are few studies that refer to the morphology of blood vessels, as well as the involvement of perivascular cells in the early maturation of ccRCC tumor vessels. Ferician and colleagues, classified intratumoral blood vessels into 4 subtypes depending on the presence and architecture of perivascular cells (12). The same team showed that VEGF165b has a major contribution in the phosphorylation of PDGF-beta. This combination most likely cause inhibitory effects on endothelial proliferation and migration, which partly explains the early maturation of blood vessels with the 4 different patterns described above.
Morphological and immunohistochemical ccRCC tumor blood vessels heterogeneity certified by previous mentioned studies suggested a possible PDGF pathway gene expression profile heterogeneity which was the aim of the present study.
TaqMan analysis revealed two genes overexpression (PIK3C3 (VPS34) and SLC9A3) common to ccRCC diffuse and reticular vascular patterns for cases with moderate and high VEGF expression.
PIK3C3 also called VPS34, is involved in intracellular membrane trafficking and autophagy, and is a target for a SAR405 compound approved in 2014 together with Everolimus, the combination giving synergistic effects as an antiproliferative of renal cancer tumor cells (35). In 2020 Jones and colleagues described three VPS34 inhibitors with proved effects on ccRCC tumor cell autophagy (36).
Although there are numerous studies on autophagy of tumor cells, studies related to autophagy in smooth muscle cells and endothelial cells of tumour blood vessels normalization are limited (37).
Currently, there are studies proving that overexpression of the PIK3C3 gene (VPs34) stimulates proliferation of perivascular smooth muscle cells in the experimental model by differentiating their SUMO1-dependent, as a morphological substrate responsible for pulmonary arterial hypertension (38). These data, correlated with the microscopic observations of an early maturation of the newly recruited tumor blood vessels in ccRCC as well as the gene overexpression of PIK3C3 (VPs34), reported in our study, support the hypothesis that this gene is not only involved in the autophagy mechanism of tumor cells but also acts on perivascular smooth muscle cells autophagy, practically performing an early remodeling of tumor blood vessels.
This may partially explain the early maturation of ccRCC tumor blood vessels, most of these tumours already having tumor blood vessels covered by perivascular cells at the time of diagnosis (20). Early maturation of tumor vessels results in a partial response or a development over time of partial or total resistance to antiangiogenic and/or antivascular therapies actually used in ccRCC.
If for perivascular smooth muscle cells the role of PIK3C3 (VPs34) is partially explained and accepted in benign lesions and less in malignant ones, the involvement of PIK3C3 (VPs34) in endothelial cell activation seems to be closely correlated with VEGF overexpression. The performance of this function is related to the interaction with the caveolin system in the endothelial cell membrane (39).
The role of PIK3C3 (VPs34) in tumor angiogenesis is indirectly suggested at this time by its interaction with the VEGF pathway, but also with hypoxia in the tumor microenvironment mediated by the HIF1α pathway (40). A proposed mechanism for the involvement of PIK3C3 (VPs34) in tumor angiogenesis is that of interaction with tyrosine kinase receptors, forming currently important targets in ccRCC therapy (41).
PIK3C3 (VPs34) is involved in the cellular internalization of VEGFR2 (41). This aspect, correlated with the maturation of blood vessels, may partly explain the low rate of endothelial cell proliferation in tumor vessels despite VEGF overexpression in tumor cells described by our team in previous studies (42).
SLC9A3, the second overexpressed gene in our study, in cases with diffuse and reticular vascular network and increased VEGF is known and studied as an ion transporter in the renal parenchyma (43). Its action on vascular smooth muscle cells is scarcely studied in the literature. The experimental model of pulmonary hypertension in rats demonstrated that overexpression of the NHE family of which SLC9A3 is a part causes proliferation and migration of vascular smooth muscle cells that are responsible for pulmonary hypertension (44, 45). The role of this gene in tumor angiogenesis is unknown. Its overexpression in our study may be related either to a study published by Karumanchi et al. (46) where the authors reported its overexpression in renal cell carcinoma cells dependent on the von Hippel-Lindau gene mutation (VHL) so common in renal cell carcinoma, or an action of hyperplasia of vascular smooth muscle cells that causes early maturation of tumor vessels in the ccRCC.
For the diffuse pattern of ccRCC vascularization, we identified the overexpression of 5 genes, most of them being part of the JAK/STAT pathway well known as having a major role in tumor angiogenesis. Activation of the JAK/STAT pathway causes proliferation of perivascular smooth muscle cells, with their hyperplasia, especially the JAK2/STAT3 axis (47) and JAK2/STAT1 (48) through a PDGF-dependent mechanism.
The JAK/STAT pathway is intensely studied in renal cell carcinoma progression and metastasis (49-51) as well as in ccRCC-associated inflammation (52) but the association between this pathway and tumor blood vessel types has not been performed to date. We found that JAK2 and STAT1 were up-regulated in our study related to ccRCC group having a diffuse pattern of tumor blood vessels which showed a high maturation grade by the presence of perivascular smooth muscle cells. Recently, Kong et al. (53) reported that the JAK2/STAT1 pathway is correlated with PD-L1 activation and expression mediated by HIF2α and IFNγ. HIF2α up-regulates genes involved in tumor blood vessel maturation (53). The JAK2/STAT1 pathway has been also reported to be involved in vascular remodeling by acting on vascular smooth muscle cells during different steps of atherosclerosis or following vascular injuries (54, 55). Bartoli et al. demonstrated that VEGF activates STAT1 in bovine aortic endothelial cells by facilitating the interaction between STAT1 and VEGFR2 (56). This may explain our finding of JAK2/STAT1 genes up-regulation in ccRCC cases with moderate and intense expression of VEGF from both diffuse and reticular pattern.
The origin of smooth muscle perivascular cells obtained by ccRCC is currently unknown. Also, the molecular mechanisms involved in the early and rapid acquisition of perivascular smooth muscle cells around ccRCC tumor vessels are unknown. Normal kidney contains cells (in both parenchyma and stroma) with the ability to transdifferentiate in smooth muscle-like cells which might be recruited by ccRCC tumor vessels during their maturation. Mesangial cells (considered to be modified pericytes) (57) were recently classified by He et al. (58) as having two distinct phenotypes: a pericyte-like phenotype and a fibroblast-like phenotype. Also, mesangial cells express FoxD1 gene which is also expressed by a population of progenitor cells that give rise to renal stroma, pericytes, vascular smooth muscle cells and mesangial cells (59-61).
The JAK2/STAT1 pathway is activated in mesangial cells during kidney response to an injury as diabetic nephropathy (62) and renal fibrosis (63) but their involvement in kidney malignancies is not yet reported. JAK2/STAT1 pathway is controlled by PDGF (62). In the normal human kidney, a high PDGF expression was reported inside mesangial cells which also have PDGF receptors (63). Moreover, PDGF has an autocrine effect on mesangial cells inducing their proliferation (64). During embryonic development of renal glomerulus, PDGF secreted by endothelial progenitor cells attracts mesangial cells and stimulates their attachment to the outer side of the capillaries inducing glomerular capillaries loops development. Our microscopic assessment of smooth muscle cells attached to ccRCC tumor blood vessels linked to our molecular findings and also related to previous presented indirect evidence suggests mesangial cells as a potential source of tumor blood vessels perivascular cells by a mechanism mediated by PDGF/PDGFRs activation through JAK2/STAT1 pathway. In our study, all 12 cases were PDGF positive by RNAscope and immunohistochemistry and in more than 90% this expression was confirmed by RT PCR. Simon et al. demonstrated that PDGF has a mitogenic effect on human airways smooth muscle cells and this effect is mediated in part by JAK2/STAT1 pathway (48). In our study the diffuse pattern of tumor vasculature includes PDGF-positive ccRCC cases having intratumor blood vessels with a continuous layer of SMAct positive perivascular cells. For this group JAK2 and STAT1 up-regulation found by TaqMan analysis may explain, in part, the rapid maturation of tumor blood vessels most probably due to mitogenic effects of PDGF on perivascular smooth muscle cells by a mechanism involving JAK2 and STAT1. PDGF stimulated smooth muscle cells activates STAT1 alpha (p91) (65) and switch to a secretory phenotype for growth factors and IFN γ (65).
STAT1 activation decreased expression of proangiogenic molecules bFGF, MMP-2, and MMP-9 and subsequently may induce a decrease of endothelial cells activation (66, 67). STAT1 action on both endothelial and perivascular cells may explain the presence of already mature tumor blood vessels at the time of ccRCC diagnosis (67).
STAT1 increased in radioresistant human ccRCC samples and inhibition of STAT1 by fludarabine and siRNAs enhanced ccRCC cell radiosensitivity (67). Banes-Berceli et al. (68) reported that levels of JAK1 and JAK2 mRNA were higher in ccRCC cells compared to normal kidney tissue and this molecular mechanism may be responsible for inappropriate cell survival and chemoresistance. Sunitinib therapy influences JAK2/STAT1 pathway by magnifying IFNg-induced STAT1 activity and causing an increase of the activity of janus kinase 2 (JAK2) (69). The same authors linked STAT1 up-regulation by indoleamine 2,3-dioxygenase (IDO) activation.
IDO has been reported to be expressed in mature tumor associated blood vessels from early stage cervical cancer (70). Also, IDO is known to be one of the immune checkpoint regulators (71). Recently, Jonasch et al. (72) proposed combination of anti-angiogenics and checkpoint inhibitors for renal cell carcinoma as a novel therapeutic strategy based on promising results of preclinical studies showing the benefits of such association, some of them targeting perivascular cells detachments. Our finding about JAK2/STAT1 pathway up-regulation for ccRCC group with diffuse pattern of tumor blood vessels may sustain this novel therapeutic strategy.
Serum response factor (SRF) was also up-regulated in diffuse vascular pattern ccRCC group. SRF increases ccRCC tumor cells migration and invasion by stimulating epithelial to mesenchymal transition (73). SRF gene is well known to have a dual role on both endothelial cells and perivascular cells. SRF is responsible for actin cytoskeleton dynamics and is critical for EC filopodia formation, tip cell contractility and EC migration (74, 75). Also, SRF is involved in epithelial to pericyte transition reported in renal tubular epithelial cells (76) as well as in cancer cells (77). This SRF action on both endothelial and perivascular cells may explain in part the high microvessels density in ccRCC, but also the predominance of mature type tumor blood vessels.
Conclusion
We herein demonstrated PDGF pathway gene expression profile heterogeneity of ccRCC related to vascular tumor pattern, invasion status and tumor grade. By correlating PDGF pathway gene expression profile and ccRCC vascular patterns we defined 3 ccRCC distinct subgroups which may have a potential impact on choosing the best therapeutic strategy for each patient. Further studies are needed to elucidate ccRCC tumor blood vessels perivascular cell origin and also to assess molecular mechanisms responsible for the early and rapid maturation of ccRCC tumor blood vessels.
Acknowledgements
The Authors are grateful to Ciprian Onica and Raluca Amalia Ceausu for their great technical support and also to Pathology Department of County Hospital Timisoara for providing us FFPE blocks. All our gratitude to Victor Babes University of Medicine and Pharmacy Timisoara, Romania for its continuous financial support for covering publishing fees and for the logistics.
Footnotes
Authors’ Contributions
AMF, OCF and AN obtained ccRCC tumours sample, FFPE blocks and selected them for immunohistochemistry and molecular studies. AMF and OCF designed the study and wrote the paper. ADC, PLB and AB (former student of VADA program from Histology Department, currently PhD student at Uniklinikum Frankfurt) performed all steps of molecular analysis. AMC supervised the study by validating each preliminary and final results and also reviewed the final draft.
Conflicts of Interest
The Authors have no conflicts of interest to declare.
- Received March 18, 2022.
- Revision received April 18, 2022.
- Accepted May 11, 2022.
- Copyright © 2022, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}