


Abstract
Esophageal cancer is associated with a dismal prognosis. The armamentarium of approved drugs is focused on chemotherapy with modest therapeutic benefit. Recently, checkpoint inhibitory monoclonal antibody Pembrolizumab was approved. In order to identify new targets and modalities for the treatment of esophagus squamous cell carcinoma (ESCC) we searched the literature for circRNAs involved in the pathogenesis of ESCC. We identified two down-regulated and 17 up-regulated circRNAs as well as a synthetic circRNA with efficacy in preclinical in vivo systems. Down-regulated circRNAs sponge microRNAs directed against tumor suppressor genes. Up-regulated circRNAs sponge microRNAs directed against mRNAs, which encode proteins with pro-tumoral functions. We discuss issues such as reconstitution of down-regulated circRNAs and inhibition of up-regulated circRNAs with short interfering RNA (siRNA)- related entities. Also, we address druggability issues of the identified targets.
Esophageal carcinoma is the seventh most common cancer worldwide with 570,000 cases annually (1). Two major histological sub-types are esophageal squamous cell carcinoma (ESCC) and esophageal adenocarcinoma (EAC). ESCCs arise in squamous cells that line the esophagus, whereas EACs derive from glandular cells of the esophagus (2). The ratio of incidence between ESCC and EAC depends on the geographic location with ESCC emerging as the most frequent subtype worldwide (3). Therapies such as endoscopic resection, surgery, chemo- and radio-therapy and treatment with immune-checkpoint inhibitory monoclonal antibodies (mAbs) only confer minimal survival advantages (4-7). Among the agents under development are epidermal growth factor receptor (EGFR), human epidermal growth factor receptor 2 (HER2), vascular endothelial growth factor receptor (VEGFR), hepatocyte growth factor (HGF)-tyrosine protein kinase c-MET and fibroblast growth factor receptor 2 (FGFR2) signaling inhibitors in the format of mAbs or small molecule tyrosine kinase inhibitors (4) as well as small molecule inhibitors of mechanistic target of rapamycin (mTOR), poly(ADP)ribose-polymerase 1 (PARP1) and cyclin-dependent kinases (CDKs) (8-10), most of them affecting rat sarcoma (RAS), rapidly accelerated firbrosarcoma (RAF), extracellular signal-regulated kinase (ERK), mitogen-activated protein kinase(MAPK), phosphoinosite-3-kinase (PI3K), serine-threonine kinase AKT and mTOR pathways.
Despite these ongoing efforts, identification of new targets and treatment modalities for ESCC is an issue of high priority. In this review, we focus on circRNAs with documented efficacy in preclinical efficacy in in vivo models for treatment of ESCC.
Circular RNA
Several types of non-coding RNAs such as t-RNA, r-RNA, siRNA, lncRNA and circRNA have been described (11). CircRNAs are covalently closed circles of RNA and are products of backsplicing events in which one exon is spliced to a preceding exon (12). A unique junction is generated which allows their specific detection by molecular probes (12). At least 30000 circRNAs have been identified by deep sequencing (12). circRNAs are expressed in all cells and tissues and are deregulated in cancer (13). Only a few of them are expressed at significant abundance (13). They are transcribed by RNA polymerase II (RNA pol II) and have no 5’-caps and polyA extensions at the 3’-end (14). In cancer they can be expressed in tumor cells, in cells of the tumor-microenvironment, immune-related cells such as T- and natural killer (NK) cells, fibroblasts and endothelial cells thus regulating tumor growth, angiogenesis and anti-tumoral immune responses (15) (Figure 1). Also, they can be incorporated into exosomes and transferred into tumor cells and cells of the tumor microenvironment (16) (Figure 1). Most circRNAs are located in the cytoplasm, and a minority is located in the nucleus. CircRNAs can function as sponges and decoys for microRNAs (miRs), as binders of RNA transcripts, mediators of protein scaffolding (17) and recently it was found that they can represent short open reading frames encoding small proteins (Figure 1) (17). circRNAs located in the nucleus can associate with RNA pol II and modulate transcription (Figure 1) (17). In this review we focus on their role as sponges for miRs. Depending on the type of miRs they are interacting with, and their decreased or increased status of deregulation they can exert an oncogenic or tumor-suppressive effect (Figure 2) (16, 17). Due to their stability and presence in blood and saliva, circRNAs are potential biomarkers for cancer (18).

Roles of circRNAs in cancer. circRNAs can be found intracellulary in the nucleus and the cytoplasm and extracellulary in exosomes. In the nucleus, they can regulate splicing and transcription, in the cytoplasm they are involved in RNA binding, sponging of miRs, protein scaffolding, synthesis of new proteins, proliferation, apoptosis, EMT, remodeling of the ECM and drug resistance. circRNAs can be localized in exosomes in blood and saliva as biomarkers for detection and progression of cancer. CircRNAs can be transferred to cells of the TME such es endothelial cells and NK cells and regulate angiogenesis and cytotoxicity. Figures were created via BioRender.com. CircRNAs are shown as coloured circles. EC: Endothelial cell; ECM: extracellular matrix; EMT: epithelial mesenchymal transition; miR: microRNA; NK: natural killer cell; TC: tumor cell.

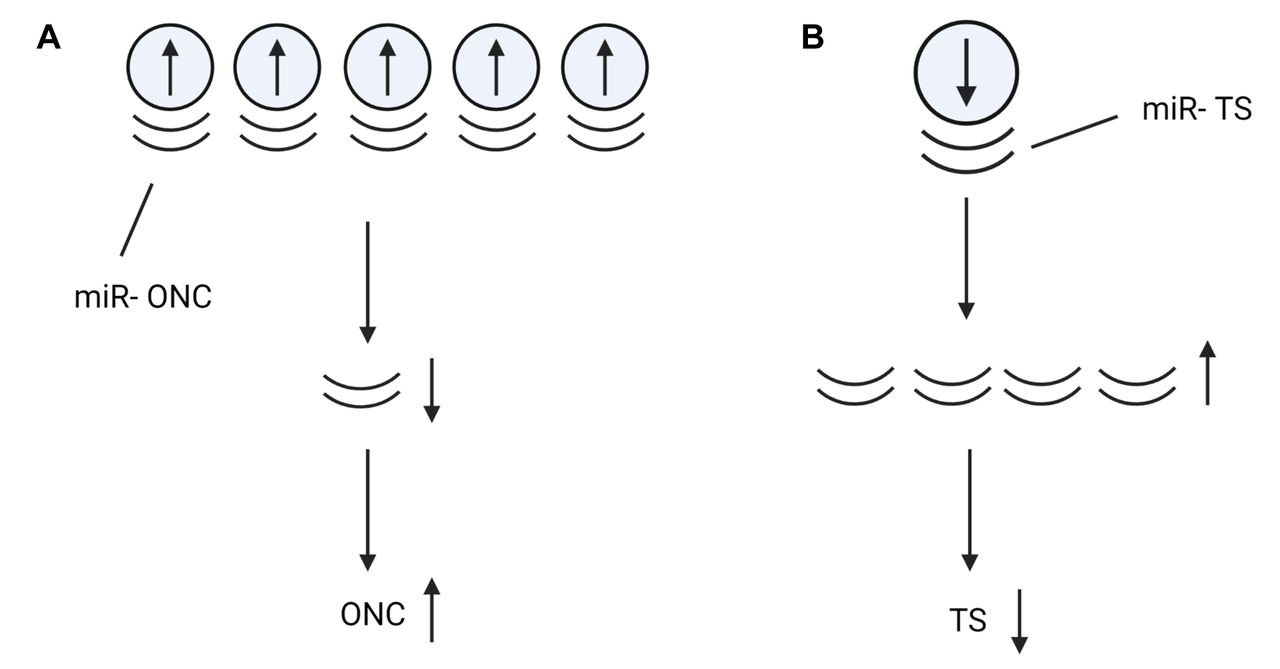
circRNAs can modulate the activity of oncogenes and tumor suppressor genes. A) Up-regulated circRNAs can sponge miRs targeting oncogenic mRNAs and thus up-regulate oncogenes. B) Down-regulated miRs targeting mRNAs for tumor suppressors lead to increased miRs targeting tumors suppressors and decrease of the corresponding proteins. TS: Tumor suppressive; ONC: oncogenic. Figures were created with Biorender.com
circRNAs were first identified as parasitic viroids in plants (19) and later in additional eukaryotic cells by electron microscopy (20). It has also been shown that circRNAs have an impact on brain function in mice (21).
Functional roles of circRNA in ESCC and their emerging role as cancer biomarkers are described in several reviews (22-25). In this review we focus on circRNAs with efficacy in ESCC-related preclinical in vivo systems. We also describe a synthetic circRNA which mediates an anti-tumoral effect. Taken together, circRNAs can affect hallmarks of cancer such as proliferation, apoptosis, epithelial mesenchymal transition (EMT), invasion, migration, metastasis, angiogenesis, anti-tumoral immune response and drug resistance.
Down-regulated circRNAs
Down-regulation of circRNAs sponging miRs can result in excess of corresponding miRs which degrade tumor suppressors.
circ-FOXO3 targets phosphatase and tensin homolog on chromosome 10. Circ-FOXO3 (Figure 3) is encoded by the human FOXO3 gene (26, 27). It was found to be poorly expressed in ESCC cell lines and tissues and inhibited growth, migration and invasion of ESCC cells in vitro and in vivo (28). circ-FOXO3 acted as a sponge for miR-23a which targets phosphatase and tensin homolog on chromosome 10 (PTEN) (28), a gatekeeper of the phosphoinosite-3-kinase (PI-3K) pathway (29).

circRNAs sponging miRs targeting tumor suppressors and transcription factors with efficacy in preclinical in vivo models of oesophageal squamous cell carcinoma. Down- or up-ward arrows: Down- or up-regulated. A) circRNAs targeting tumor suppressors. circ FOXO3 and circ CNTAP3 are down-regulated. Sponged miRs and targeted proteins are indicated. B) circRNAs sponging microRNAs targeting mRNAs for transcription factors circ 6948, circ LRP6 and CiRS-7 are up-regulated and corresponding microRNAs and targets are shown. HMGA2: High mobility group A2; HOXB13: homeobox protein B13; MYC: transcription factor MYC; PTEN: tensin homolog on chromosome 10. Figures were created with Biorender.com
circ-CNTNAP3 targets RNA-binding motif protein 25. circ-CNTNAP3 (Figure 3) derives from exons 5 to 9 of the CNTNAP3 gene and was identified by screening for circRNA expression in ESCC (30). It was poorly expressed in ESCC cell lines and tissues (30). Knock-down promotes cell proliferation in wild-type p53 Eca-109 ESCC cells, but not in p53 mutant ESCC cells such as KYSE-450 and TE-1 cells (30). In vivo, over-expression of circ-CNTNAP3 suppressed growth of Eca-109 cells by acting as a sponge for miR-153-5p which targets p53 (30). miR-153-5p reversed the TS role of circ-CNTNAP3 in p53 wild-type ESCCs (30). RNA-binding motif protein 25 (RBM25) was found to be a direct transcriptional target of p53 (30, 31). A positive feedback expression loop was identified based on over-expression of p53 and increased expression of circ-CNTNAP3 (30). Low expression of circ-CNTNAP3 indicated poor prognosis in p53 wild-type ESCC patients (30). p53 acts as a TS and its loss is a driver of metastasis (32, 33). Gene transfer of p53 into ESCC tumors has been pursued as a modality for treatment of ESCC (34).
Up-regulated circ-RNAs. Up-regulation of circRNAs can lead to down-regulation of miRs which inhibit proteins with oncogenic functions such as transcription factors, mediators of cellular signaling, components of the extracellular matrix and further protumoral functionalities.
CircRNAs Sponging miRs Directed Against Transcription Factors
Circ-0006948 targets high mobility group A2. Circ-0006948 (Figure 3) originates from exons 2, 3 and 4 of the fibronectin type III domain containing 3B (FND3B) gene (35). Circ-0006948 was shown to be up-regulated in ESCC and was related to poor survival (35). It promoted proliferation, invasion, migration and epithelial mesenchymal transition in TE-1 and KYSE30 cells (35). In vivo, circ-0006948 mediated tumor growth of TE-1 cells after subcutaneous injection into nude mice. It functioned as a sponge for miR-490-3p and mediated up-regulation of its target high mobility group A2 (HMGA2) (35). The latter acts as a transcription factor by binding to AT-rich regions of DNA and functions as a critical regulator in cancer development (36). HMGA2 is over-expressed in several types of cancer and its over-expression can induce tumorigenesis (37-39). HMGA2 promotes cell-cycle entry, inhibits apoptosis, promotes EMT, supports cancer stem cell (CSC) phenotype and chemo-resistance and activates MAPK/ERK, transforming growth factor β (TGFβ)/SMAD), PI3K/AKT/mTOR, nuclear factor ĸB (NF-ĸB) and signal transducer and activator of transcription 3 (STAT3) pathways (37-39). Several small molecule inhibitors interfering with HMGA2/DNA interaction have been identified (40). HMGA2 over-expression plays a critical role in the progression of ESCC (41, 42).
circ-LRP6 targets the transcription factor MYC. circ-LRP6 (Figure 3) derives from the low-density lipoprotein receptor-related protein 6 (LRP6) gene (43, 44). It was found to be up-regulated in ESCC tissues and was mainly located in the cytoplasm (44). Knockdown of circ-LRP6 inhibited proliferation and invasion of TE-1 and EC109 ESCC cells in vitro and decreased TG of EC109 ESCC cells in vivo (44). Circ-LRP6 acted as a sponge for miR-182, which targets transcription factor MYC (44). MYC dimerizes with MYC-associated factor X (MAX) to exert its oncogenic functions such as proliferation, invasion, metastasis and angiogenesis (45-48). Inhibition of MYC transcription, disruption of MYC/MAX dimers, inhibition of post-translational regulation and proteolytic degradation of MYC are drug discovery approaches in preclinical and clinical stages of development (49, 50).
CiR-S7 targets homeobox protein B31. Up-regulation of CiR-S7 (Figure 3) and down-regulation of miR-7 have been shown to predict poor prognosis of ESCC patients (51). Over-expression of CiR-S7 promotes proliferation and invasion of Eca109 and KYSE150 ESCC cells by sponging miR-7 (51). Homeobox protein B31 (HOXB13) has been identified as a target of miR-7 (51). In vivo, CiR-S7 increased lung metastasis of Eca109 cells after tail vein injection (51). HOXB13 activates NFĸB signaling by phosphorylation of NFĸB/p65 (51). Forced expression of HOXB13 confers oncogenic hallmarks to esophageal keratinocytes (52), promotes cancer cell growth and predicts poor survival in ESCC patients (53, 54). HOX genes are master regulatory transcription factors during embryogenesis and HOXB13 is commonly altered in human cancers (55).
circRNAs Sponging miRs Directed Against Signaling Components
circ-0000654 targets interleukin 6. High circ-0000654 (Figure 4) is correlated with higher T stage and lymph node metastasis in ESCC patients (56). circ-0000654 enhanced proliferation, migration and invasion of KYSE450 ESCCs in vitro (56). In vivo, circ-0000654 promoted TG after subcutaneous implantation and lung metastasis after tail vein injection in nude mice (56). The effects are mediated by sponging of miR-149-5p which targets interleukin 6 (IL6) (56). IL6 is a tumor promoting cytokine, which binds to IL6 receptor (IL6R) and activates intracellular signals such as STAT3 phosphorylation via glycoprotein 130 (gp130). STAT3 is transferred to the nucleus and activates genes such as cyclin B1 and B-cell lymphoma extra-large (Bcl-xL), MYC, induced myeloid leukemia cell differentiation protein (MCL1) and vascular endothelial growth factor (VEGF) (57, 58). IL6 signaling is aberrantly hyperactivated in many types of cancer (59). IL6 is linked to the progression of ESCC and its inhibition is a potential strategy for treatment of this disease (60).

circRNAs sponging miRs affecting mRNAs for components of signaling pathways and the extracellular matrix with efficacy in preclinical in vivo models of esophageal squamous cell carcinoma. Down- or up-ward arrows: Down- or up-regulated. CDC25A: Cell-division cycle 25A; COL1A1: collagen1A1; FNDC3B: fibronectin type III domain containing protein 3B; IL6: interleukin 6; MET: transmembrane tyrosine kinase MET; RAB10: ras-related protein 10; RHOC: Ras homology member, member C; WNT3: proto-oncogene protein WNT3. Figures were created with Biorender.com.
circ-0072088 targets vascular endothelial growth factor A. circ-0072088 (Figure 4) is increased in ESCC tissues and cell lines (61). It derives from exons 13 to 17 of the ZFR gene, which encodes a zinc finger transcription factor (62). It promoted proliferation, migration and invasion in TE-13 and ECA109 ESCCs in vitro (61). In vivo, circ-0072088 promoted TG in nude mice after subcutaneous implantation (61). This leads to up-regulation of VEGF by sponging of miR-377 (61). Bevacizumab, a mAb directed against VEGF-A and Ramicurumab, a mAb directed against VEGFR2, are under clinical investigation in ESCC (63, 64), as well as several small molecule inhibitors of the VEGFR tyrosine kinases such as sunitinib, sorafenib, apatinib and anlotinib (65).
circ-LPAR3 targets transmembrane receptor kinase MET. circ-LPAR3 (Figure 4), which derives from the lysophophosphatic acid receptor 3 gene, was increased in human ESCC tissues and cell lines (66). It promoted migration and invasion of KYSE-450 ESCC cells, but had no impact on proliferation (66). In vivo, in TE-13 ESCC xenografted cells, circ-LPAR3 functioned as a mediator of lung metastases after tail vein injection into nude mice (66). circ-LPAR3 mediated activation of MET by sponging miR-198 which targets MET. Together with its ligand HGF, MET is an essential driver of oncogenesis and metastasis in many types of tumors (67). MET can activate many oncogenic pathways such as PI3K/AKT/mTOR, MAPK/ERK1/2 and cell division control protein 42 homolog (CDC42)/ras-related C3 botulinum toxin substrate1 (RAC1) (68). Furthermore, MET can be activated by gene amplification, mutations, gene fusion or exon 14 skipping (68, 69). MET is over-expressed preferentially in EAC, however, MET expression in ESCC has to be studied in more detail (70). Therefore, mAbs directed against HGF (Rilotuzumab, AMG102) and MET (Obinutuzumab) are in clinical development in patients with EAC or tumors of the gastroesophageal junction (71). AMG 377, a highly selective small molecule, which inhibits HGF/MET signaling, is in clinical trials in patients with several types of cancer (72).
circ-100367 targets proto-oncogene WNT3. circ-100367 (Figure 4) was found to be highly expressed in radio-resistant ESCC cell lines such as KYSE-150R and its silencing inhibited proliferation, invasion and migration of this cell line (73). Silencing of circ-100367 reduced the growth of KYSE-150R cells under radiation in nude mice (73). circ-100367 interacted with miR-217, which targets proto-oncogene protein WNT3 (73). The latter promotes stabilization of β-catenin to regulate radio-sensitivity (73, 74). WNTs are secreted lipid modified glycoproteins with 16 members in humans (75). They interact with the frizzled family of receptors and co-receptors such as lipoprotein-related protein 5 and 6 (LRP5,6) and their signaling activity is highly context-dependent (76).
circ-AKT3 targets ras homolog family, member C and transcription factor STAT3. circ-AKT3 (Figure 4) derives from the AKT3 gene and was identified in gastric cancer cells with cis-Pt resistance (77). Circ-AKT3 was found to be over-expressed in ESCC and induced proliferation, migration and invasion of TE-1 ESCCs (78). circ-AKT3 knockdown suppressed TG of TE-1 ESCCs in nude mice (78). miR-17-5p was sponged by circ-AKT3 resulting in expression of ras homolog family, member C (RHOC) and STAT3 (78). RHOC is a member of the protumoral ras homolog family and acts as a cytoskeleton organizer (79, 80). RHOC is related to lymph node metastasis and survival in ESCC patients (81). The role of transcription factor STAT3 in tumor formation, metastasis and drug resistance has been discussed previously in this review (57, 58). Recently, STAT3 has been identified as a mediator of immune suppression in tumors (82).
circ-UBAP2 targets ras-related protein RAB10. circ-UBAP2 (Figure 4) derives from ubiquitin-associated protein 2 and is up-regulated in ESCC tissues (83). Knockdown of circ-UBAP2 with lentivirus small hairpin RNA (shRNA) mediated inhibition of proliferation, invasion and migration of ESCC cells in vitro (83). Vice versa, circ-UBAP2 induced proliferation, migration and invasion of ESCCs (83). In vivo, down-regulation of circ-UBAP2 suppressed TG of ESCCs in nude mice (83). Circ-UBAP2 sponged miR-422a, which targets ras-related protein RAB10 (83). circ-UBAP2 promoted tumorigenesis in osteosarcoma, lung cancer, triple-negative breast cancer and cervical cancer (84-87). RAB10, a member of the superfamily of small GTPases, can regulate intracellular vesicle trafficking and its over-expression promotes TG of hepatocellular carcinoma (HCC) and correlates with poor prognosis (88).
Circ-0004771 targets cell-division cycle 25A. Circ-0004771 (Figure 4) was found to be up-regulated in ESCC tissues and its expression correlated with poor prognosis (89). Knockdown of circ-0004771 inhibited proliferation of ESCC cells in vitro and in vivo (89). It acted as a sponge for miR-339-5p, which targets cell-division cycle 25A (CDC25A) (89). CDC25A is a dual-specific phosphatase, which hydrolyses tyr- and ser-phosphates by dephosphorylation (90). CDC25A is over-expressed in many types of tumors and activates the cyclin-dependent kinase complex which promotes transition of G1/S and G2/M phase of the cell-cycle and plays a role in the DNA damage response (91, 92). In ESCC, targeting of CDC25A mediates radio-sensitivity (93).
CircRNAs Sponging miRs Directed Against Components of The Extracellular Matrix
Circ-0004370 targets collagen 1A1. Expression of circ-0004370 (Figure 4) was found to be increased in ESCC tissues in comparison to normal controls (94). In OE19 and EC109 ESCCs, knockdown of circ-0004370 suppressed proliferation, invasion and migration in vitro and in vivo after subcutaneous implantation into nude mice (94). Circ-0004370 sponged mir-1310-3p, which targets collagen 1A1 (Col1A1) (94). Collagen, an abundant component of the ECM, plays an important role in EMT and TG in gastric cancer (95). COL1A1 knockdown suppresses metastasis of breast cancer (96). COL1A1 was found to be up-regulated in ESCC compared to normal controls (97) and its expression is associated with poor prognosis (98).
Circ-0005948 targets fibronectin type III domain-containing protein 3B. circ-0005948 (Figure 4) was up-regulated in ESCC tissues and cells (99). Down-regulation of expression of circ-0005948 repressed growth, migration and invasion and EMT in vitro and TG in vivo of ESCCs (99). circ-0005948 sponged miR-4262, which targets fibronectin type III domain-containing protein 3B (FNDC3B) (99). FNDC3B, a protein of the extracellular matrix (ECM), also known as factor for adipocyte differentiation 104, is a member of the fibronectin family and contains nine fibronectin III domains and one transmembrane domain (100). It can be amplified in HCC, act as an oncogenic driver and promote EMT, PI3K/AKT, retinoblastoma (Rb) and TGFβ signaling (100). Over-expression of FNDC3B predicts poor prognosis in lung adeno-carcinoma (101).
Circ RNAs Sponging miRs Targeting Additional Protein Categories
Circ-0003340 targets protein for Xklp2. circ-0003340 (Figure 5) was shown to be over-expressed in ESCC in comparison to adjacent normal tissues and non-transformed human esophagus squamous cells Het-1a (102). Knockdown of circ-0003340 induced apoptosis and inhibited proliferation in EC1 and EC9706 ESCCs in vitro and TG in vivo (102). circ-0003340 has a binding site for mir-562, which inhibits targeting protein for Xklp2 (TPX2) (102). TPX2 acts as a spindle assembly factor, mediates growth during M phase as well as cell invasion and metastasis (103, 104). TPX2 recruits and activates ser-thr kinase aurora A, which is a validated, druggable target for cancer therapy and corresponding inhibitors are evaluated in several clinical studies in cancer patients (105). TPX2 has been shown to promote growth of EC9706 xenografts in vivo (106). TPX2 may be useful as a potential biomarker for tumor progression, metastasis and prognosis in patients with ESCC (107).

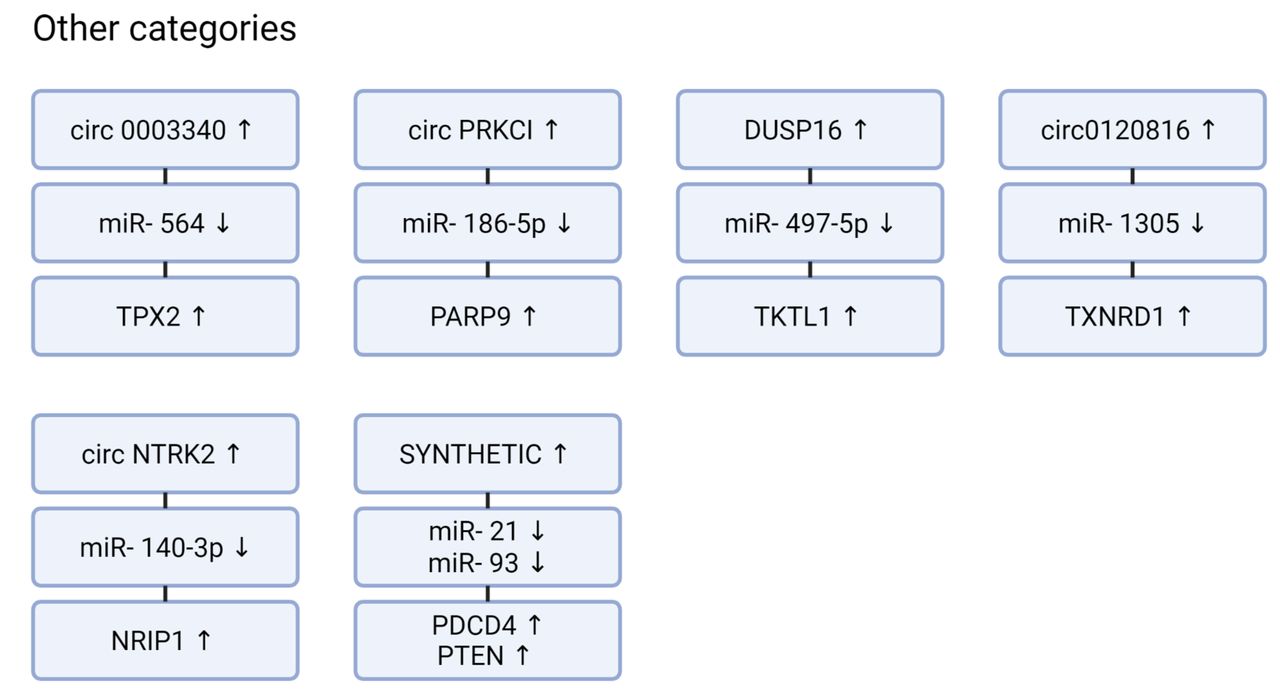
circRNAs sponging miRs affecting mRNAs of diverse other components with efficacy in preclinical in vivo models of esophageal squamous cell carcinoma. Down- or up-ward arrows: Down- or up-regulated. PARP9: Poly(ADP-ribose) polymerase, family member 9; PDCD4: programmed cell death protein 4; PTEN: tensin homolog on chromosome 10; NRIP1: nuclear receptor interacting protein 1; TKTL1: transketolase-like 1; TPX2: targeting protein for Xklp2; TXNRD1: thioredoxin 1. Figures were created with Biorender.com.
Circ-PRKCI targets poly(ADP-ribose) family member 9. circ-PRKCI (Figure 5) and poly(ADP-ribose) family member 9 (PARP9) were up-regulated and miR-186polymerase-5p was down-regulated in ESCC tissues in comparison to adjacent normal tissues (108). circ-PRKCI derives from the protein kinase C iota type (108). Knockdown of circ-PRKCI decreased cell viability, colony formation and elevated radiosensitivity of ESCC in vitro and inhibited TG in vivo (108). Mechanistically, circ-PRKCI sponged miR-186-5p and up-regulated PARP9 (108, 109). PARP9 is over-expressed in breast cancer and promotes cell migration (110). In lymphoma and prostate cancer, PARP9 promotes metastasis and correlates with poor prognosis (111, 112). From a mechanistic point of view, the function of PARP9 in tumor progression is not yet resolved.
Circ-DUSP16 targets dual specificity phosphatase 16. circ-DUSP16 (Figure 5) derives from dual specificity phosphatase 16 and was found to be elevated in ESCC tissues and hypoxia stimulated ESCC cells (113). Knockdown of DUSP16 suppressed hypoxia-induced ESCC viability, colony formation, migration, invasion and glycolysis in vitro and inhibited TG of ESCC cells in vivo (113). DUSP16 positively regulated transketolase like-1 (TKTL1) by sponging miR-497 (113). TKTL1 is a thiamine diphosphate (vitamin1)-dependent enzyme, which catalyses several key reactions in the non-oxidative branch of the pentose phosphate pathway; It has been identified as a marker of the anaerobic glucose metabolism (Warburg effect) and is involved in the generation of NADPH and ribose-5-phosphate (114, 115). It is also involved in cell proliferation, lipid synthesis, ATP generation, inhibition of apoptosis, angiogenesis and lactate-based inhibition of the immune system by T- and NK cells and therapy resistance (114, 115). TKTL1 promotes proliferation and metastasis of ESCC (116, 117). The druggability of TKTL1 and its prospect for cancer treatment are still under evaluation.
circ-12086 targets thioredoxin 1. circ-12086 (Figure 5) was shown to be up-regulated in ESCC tissues, enhanced cell proliferation, cell viability and cell adhesion and inhibited apoptosis in KYSE450 and KYSE510 ESCCs in vitro (118). In vivo, its inhibition led to TG inhibition and attenuation of lung metastasis after subcutaneous implantation in KYSE450 cells (118). circ-12086 sponged miR-1305, which targets thioredoxin 1 (TXNRD1) (118). The latter is a cytosolic selenocysteine containing enzyme, which is over-expressed in many types of cancer (119). It is involved in adaption of cancer cells to stress (120, 121). Irreversible TXNRD1 inhibitors have been identified, which inhibit growth of xenografts and syngeneic mouse tumors without overt cytotoxicity (122).
circ-NTRK2 targets nuclear receptor-interacting protein 2. circ-NTRK2 (Figure 5) derives from the neurotrophin receptor tyrosine kinase 2 gene, and was found to be up-regulated in ESCC patients and its expression correlated with poor prognosis (123). In vitro, knockdown of crc-NTRK2 inhibited proliferation, invasion and EMT and accelerated apoptosis in Eca-109 and KYSE-150 ESCCs (123). In vivo, silencing of NTRK2 led to inhibition of TG of KYSE 150 cells after subcutaneous implantation in nude mice (123). circ-NTRK2 sponged miR-140-3p leading to up-regulation of nuclear receptor-interacting protein 2 (NRIP2), also known as receptor interacting protein 140 (RIP140) (123). It interacts with the hormone-dependent activation function 2 domain (AF2) of nuclear receptors and modulates transcriptional activity of several transcription factors, including estrogen receptor (124). NRIP1 regulates various oncogenic signaling pathways and participates in progression of solid tumors (125). Suppression of NRIP1 inhibits growth of breast cancer cells in vitro and in vivo (126). NRIP1 is also involved in cachexia in mice (127).
circ 21-93 targets PTEN, programmed cell death protein 4 and transforming growth factor receptor β2. Using enzymatic ligation, circ 21-93, a circular multi-miR sponge was generated which is more stable than its linear counterpart (128). It contains two binding sites for miR-21 and three binding sites for miR-93 (128). circ 21-93 decreased SKGT4 and OE33 ESCC transfectants proliferation and migration and increased apoptosis in vitro (128). In vivo, circ 21-93 suppressed TG of xenografts after subcutaneous implantation into nude mice (128). Tumor suppressors PTEN, programmed cell death protein 4 (PDCD4) and transforming growth factor receptor β2 (TGFRβ2) have been identified as targets for miR-21 and -93 (129-132).
Technical Issues
Down-regulated circRNAs. circ-FOXO3 and Circ-CNTNAP3 are down-regulated in ESCC tissues and have been identified as sponges for miRs-23a and -513a-5p, which target PTEN and p53, respectively (28, 30). Therapeutic intervention can be based on reconstitution of the corresponding TS by small molecules or gene transfer. However, both approaches have their limitations with respect to specificity and delivery issues. Proof-of-concept (POC) experiments have shown that TG can be inhibited in vitro and in vivo by transfecting ESCC with the corresponding circRNAs. However, extrapolation of these preclinical findings to clinical studies would require to solve issues such as targeting ESCC cells and optimization of transfection efficacy. The same considerations hold true for transfer of synthetic circ-RNAs acting as sponges for miRs targeting TS mRNAs (128).
Up-regulated circRNAs. POC experiments as outlined in the previous sections have indicated that up-regulated circRNAs can exhibit an oncogenic function by sponging miRs, which inactivate mRNA encoding for tumor-promoting proteins in ESCC tumor cells. circRNAs are generated by a backsplicing event generating specific new junctional sequences; therefore, circRNA can be specifically targeted by short interfering RNA (siRNA) or short hair-pin RNA (shRNA) (133-135). siRNA is chemically synthesized double-stranded RNA, 19-23 base pairs in length with 2nts unpaired at the 5’-phosphorylated and 3’-unphosphorylated ends (136). shRNA are artificial RNA molecules with tight hairpin loops, which also silence target gene expression via RNA interference (137). siRNA and shRNA represent exogenously added RNAs, whereas miRs are synthesized intracellularly. All rely on the endoribonuclease DICER for processing (136). RNAi requires stringent complementarity pairing to the corresponding target, whereas in the case miRs more relaxed specificity of base-pairing has been observed (137). As of January 2020, ten oligonucleotide drugs have received regulatory approval. These drugs are focused on local delivery such as eye, spinal cord or liver (138, 139). For example, Onpattra (Patisiran), an agent approved for treatment of transerythrin-mediated amyloidosis that silences production of an abnormal form of transerythretin, represents a chemically modified anti-transerythrin oligo-nucleotide formulated in liposomes targeted to the liver (139).
However, from a broader perspective, delivery of siRNA into tumor cells is impeded by several factors such as intravascular degradation, renal clearance, recognition by the immune system, tumor tissue penetration, uptake into TC, endosomal release and off-target-effects (140, 141). Delivery of siRNA directly is not recommended due to its short half-life ranging from a couple of minutes to half-an hour (140) and its low targeting and transfection efficacy. Its negative charge due to the anionic backbone prevents it from permeating the plasma membrane and gives rise to toxicity (141). Therefore, chemical modification and development of delivery methods and corresponding vehicles are of paramount importance. Chemical modifications of the backbone, nucleobases, ribose and alternative chemistries, have improved pharmaco-kinetic and pharmaco-dynamic properties of siRNA (142). Conjugation of siRNA with polyethylene glycole and cholesterol have improved serum stability and prevented renal clearance (143, 144). Formulation of siRNA into nanoparticle-based structures has improved half-life of siRNA and its delivery to TC due to facilitated membrane fusion. Lipid nano-particles, composed of lipids play an important role in delivery of si-RNA (145), and prevention of immunogenicity and cytokine-release syndrome. An important step in the development of siRNA delivery is the development of dynamic polyconjugates (146). This technology includes a membrane active polymer, which is masked until it reaches the acid environment of the endosomes and induces burst of their membrane. Targeting of lipid nanoparticles can be achieved by conjugation to antibodies, ligands, aptamers and RGD peptides as shown for transmembrane antigens such as integrins, prostate-specific membrane antigen and folate receptor (147-149). A breakthrough for delivery of siRNA to the liver was the identification of the asialoglycoprotein complex on hepatocytes which is targeted by entities containing N-acetylgalactosamine (GalNAc) and allow targeting of siRNA conjugated with GalNAc to hepatocytes (150, 151). For ESCC, specific ligands, antibodies or transmembrane receptors have not yet been identified. In order to improve cellular uptake of siRNA complexation with viral proteins such as trans-activator of transcriptin (TAT) (152) or penetratin (drosophila) (153), have improved delivery of siRNA through the cell membrane as well as endosomal escape.
Targets
P53, PTEN, PDCD4 and TGFβR2 are TS frequently down-regulated in ESCC (129-132). As already outlined previously, their functional reconstitution by small molecules or gene therapy is problematic from a drug discovery point of view.
Regarding oncogenic circRNAs, delivery of siRNA into TC can inhibit their growth in vitro and in vivo. It is important to mention that in lung adenocarcinoma and gastric patient derived xenografts (PDX), corresponding cholesterol-conjugated siRNAs have mediated in vivo efficacy (154, 155). For ESCC, data derived from PDX-derived models are not yet available.
circRNAs can mediate an oncogenic effect by sponging miRs which inactivate oncogenic proteins. We have identified targets which can be grouped into transcription factors, signaling related entities, components of the ECM and targets, which do not match with these major categories. Inhibitors of the identified targets VEGF and MET, such as small molecule tyrosine kinase inhibitors or antagonizing antibody moieties, are in clinical trials in esophageal cancer patients as outlined in the previous text (63-65, 71, 72). TPX2 activates Aurora kinase A, which is clinically evaluated in several types of cancer (105). WNT3 signaling is context-dependent and deserves further validation in ESCC (73). CDC25A, is difficult to target and alerts toxicity issues due to ubiquitous expression of this enzyme (91, 92). It will be of interest to evaluate the identified TXNRD1 inhibitors in several in vivo models of ESCC as a further path for their clinical evaluation (122). A couple of potential targets such as the transcription factors HMGA2 (37-39), MYC (45-48), HOXB13 (53, 54), STAT3 (57, 58), the GTPases RHOC and RAB10 (79, 80, 88) and NRIP1 are associated with druggability problems (125). However, proteolysis targeting chimeras (PROTACs) are on the horizon. PROTACs mediate degradation of the corresponding targets by the proteasome after their ubiquitinylation (156-158). At least 15 targeted degraders will be in clinical studies until the end of 2021 with targets such as androgen receptor (AR), estrogen receptor (ER), Bcl-xL, the zinc finger transcription factors Helios, Ikaros, Aiolos, GTPase1, glutathion P-transferase 1 and members of the bromodomain family, such as BRD9 (159). Another interesting target identified by our search is TKTL1, which is dependent on vitamin B1 as a cofactor (114, 115). The druggability of this enzyme is unclear, since it has not been shown yet that vitamin B1 antagonists or other inhibitory compounds give rise to in vivo inhibition of ESCC-related xenografts. The identified components of the ECM, COL1A1 and FNDC3B, might be targeted with corresponding antibodies (94, 99). With respect to targeting ESCC, identification of corresponding transmembrane antigens is an important issue. Claudin 18.2 is expressed in some esophageal cancers and members of the FGFR family deserve further investigation (160, 161). Taken together, the field of circRNAs in ESCC is still in its infancy and its translational impact deserves further investigation.
Footnotes
Authors’ Contributions
The Authors contributed equally to all aspects of the article.
Conflicts of Interest
UB, TS, JN are and UHW was an employee of Roche.
- Received January 27, 2022.
- Revision received March 2, 2022.
- Accepted March 3, 2022.
- Copyright© 2022, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY-NC-ND) 4.0 international license (https://creativecommons.org/licenses/by-nc-nd/4.0).
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- Circular RNA
- Down-regulated circRNAs
- CircRNAs Sponging miRs Directed Against Transcription Factors
- circRNAs Sponging miRs Directed Against Signaling Components
- CircRNAs Sponging miRs Directed Against Components of The Extracellular Matrix
- Circ RNAs Sponging miRs Targeting Additional Protein Categories
- Technical Issues
- Targets
- Footnotes
- References
- Figures & Data
- Info & Metrics
Related Articles
Cited By...
- Hepatocellular Carcinoma: Up-regulated Circular RNAs Which Mediate Efficacy in Preclinical In Vivo Models
- Up-regulated Circular RNAs in Colorectal Cancer: New Entities for Therapy and Tools for Identification of Therapeutic Targets
- Circ_ARHGAP32 acts as miR-665 sponge to upregulate FGF2 to promote ox-LDL induced vascular smooth muscle cells proliferation and migration