


Abstract
Background/Aim: Sarcomas are considered a heterogeneous disease with incomplete understanding of its molecular basis. In the present study, to further understand general molecular changes in sarcoma, patient-derived xenograft (PDX) mouse models of the three most common soft-tissue sarcomas: myxofibrosarcoma, undifferentiated pleomorphic sarcoma (UPS) and liposarcoma were established and the methylation status of histone H3 lysine marks was studied. Materials and Methods: Immunoblotting and immunohistochemical staining were used to quantify the extent of methylation of histone H3K4me3 and histone H3K9me3. Results: In all 3 sarcoma types in PDX models, histone H3K4me3 and H3K9me3 were found highly over-methylated compared to normal muscle tissue. Conclusion: Histone H3 lysine over-methylation may be a general basis of malignancy of the major sarcoma types.
Sarcomas are a heterogeneous group of cancers and the molecular basis of their malignancy is poorly understood (1–3). Histone methylation at lysine residues can control large numbers of genes, and is therefore a promising area to study the basis of malignancy (4). Recently, there has been intense interest in the epigenetic control of cancer by specific histone methylation modifications, including histone H3K4me3 and H3K9me3, which are thought to be global gene regulators (5–7). H3K4me3 binds to gene-activating proteins, including transcription factors. H3K9me3 is involved with repressing gene expression (4). Histone lysine marks appear to be over-methylated in some cancers (5, 6).
There have been a number of publications, in recent years, attempting to correlate the extent of methylation of histone H3 lysine with patient outcomes in cancers of the liver, cervix and colon, with inconclusive results (8-10).
In the present report, we show that the most common types of soft tissue sarcoma (STS), myxofibrosarcoma, undifferentiated pleomorphic sarcoma (UPS) and liposarcoma, growing in PDX mouse models, all have over-methylated histone H3K4me3 and H3K9me3 lysine marks, in contrast to normal muscle in the host nude mouse. Histone H3 lysine over-methylation is discussed as a potential general alteration in apparently heterogeneous types of sarcoma.
Materials and Methods
Mice. Athymic female nu/nu nude mice (AntiCancer, Inc., San Diego, CA, USA), 4–6 week old, average weight approximately 20 g, were used to establish PDX mouse models of three types of STS: myxofibrosarcoma, undifferentiated pleomorphic sarcoma (UPS), liposarcoma. Each sarcoma tissue was implanted in 5 mice. All mice were maintained under standard conditions of 12 h light/dark cycles and fed autoclaved water and a laboratory rodent diet. All studies were performed with a protocol approved by an AntiCancer, Inc. Institutional Animal Care and Use Committee (IACUC) following the principles and procedures provided in the National Institutes of Health (NIH) Guide for the Care and Use of Animals under Assurance Number A3873-1. In order to minimize any suffering of the animals, they were anesthetized by subcutaneous injection of anesthesia and analgesics, which consist of 20 mg/kg ketamine, 15.2 mg/kg xylazine, and 0.48 mg/kg acepromazine maleate, for all surgical experiments. In order ensure the depth of the anesthesia, animal response during surgery was monitored. The animals were sacrificed by CO2 inhalation humanely when they met the following humane endpoint criteria: severe tumor burden (more than 20 mm in diameter), prostration, significant body weight loss, difficulty breathing, rotational motion and body temperature drop. Tumor volume and body weight were measured once a week with the formula as follows: tumor volume (mm3)=length (mm) × width (mm) × width (mm) × 1/2 (11–14).
Patient-derived xenografts (PDX)
Myxofibrosarcoma. A resected tumor tissue specimen resected from a 64-year-old woman with right anterior-thigh myxofibrosarcoma was previously obtained from surgery at the UCLA Medical Center, with written informed consent by the patient and Institutional Review Board approval (IRB#10-001857), and transported immediately to the laboratory at AntiCancer, Inc. The tumor specimen was cut into 5-mm3 fragments, with 4 fragments implanted subcutaneously per mouse as previously reported (11). The time interval between tumor implantation and subsequent tumor harvest was approximately 3 months.
Undifferentiated pleomorphic sarcoma (UPS). A resected tumor tissue specimen resected from 65-year-old man with a right-high UPS was previously obtained at surgery at the UCLA Medical Center, with written informed patient consent and Institutional Review Board approval (IRB#10-001857), and brought to AntiCancer, Inc. The tumor specimen was cut into 5-mm3 fragments, with 4 fragments implanted per mouse as previously described (12, 13). The time interval between tumor implantation and subsequent tumor harvest was approximately 2 months.
Liposarcoma. A tumor specimen was resected from a 67-year-old man with upper-abdominal soft-tissue liposarcoma, at the UCLA Medical Center. The tumor was previously brought to AntiCancer, Inc., with written informed patient consent and Institutional Review Board approval (IRB#10-001857). The tumor specimen was cut into 5-mm3 fragments, with 4 fragments implanted subcutaneously per mouse as described above (14). The time interval between tumor implantation and subsequent tumor harvest was approximately 4 months.
Immunoblotting. The sarcoma PDX mouse models were sacrificed and tumor and femoral muscle tissues were resected. Then tissues were frozen immediately after resection in liquid nitrogen. To prepare for immunoblotting, the tumors were homogenized and the histones were extracted using a Epiquik Total Histone Extraction Kit (Epigentek, Farmingdale, NY, USA). Immunoblotting for histones was performed with the following procedure: 12% SDS-PAGE gel-electrophoresis and transfer to 0.2 μm polyvinylidene difluoride (PVDF) membranes were used. Blocking was performed using the Bullet Blocking One for Western Blotting Kit (Nakalai Tesque, Inc. Kyoto, Japan). Anti-H3K4me3 antibody (1:1,000, #9751, Cell Signaling Technology, Danvers, MA, USA); anti-H3K9me3 antibody (1:1,000, #13969, Cell Signaling Technology); and anti-H3 antibody (1:1,500, 17168-1-AP, Proteintech, Rosemont, IL, USA) were used. Total histone H3 was used as a loading control. Horseradish-peroxidase-conjugated anti-rabbit IgG (1:20,000, SA00001-2, Proteintech) was used as a second antibody. Immunoreactivity was visualized using Clarity Western ECL Substrate (Bio-Rad Laboratories, Hercules, CA, USA). The signals were detected with a UVP ChemStudio (Analytik Jena, Upland, CA, USA) (6, 7). Three repeated immunoblots were made for each sarcoma PDX. One homogenate was made per tumor and then analyzed on three separate gels.
Hematoxylin and eosin (H&E) staining. After resection of the sarcoma and femoral muscle tissues from PDX mice, the tissues were fixed with 10% formalin overnight before embedding in paraffin. The tumor and muscle tissues were sectioned and specimens were deparaffinized with xylene and rehydrated in a series of ethanol. Staining with H&E was performed in accordance with standard protocol.
Immunohistochemical (IHC) staining. Tissues were prepared and sectioned as described above. Autoclaving was then performed in order to retrieve antigen at 120°C for 5 min with a citrate-buffer solution (pH 6.0). Next, the specimens were immersed in absolute methanol containing 0.3% hydrogen peroxide solution in order to block endogenous peroxidase activity at room temperature for 30 minutes. Then, the specimens were blocked using 10% bovine serum albumin at room temperature for 15 min and incubated with H3K4me3 antibody (1:1,000) at 4˚C overnight. IHC reactions were detected with HistoFine (Nichirei, Tokyo, Japan) and Dako Omnis (Agilent, Santa Clara, CA, USA). Finally, the specimens were counterstained with hematoxylin (6, 7).
Results
Histone H3K4me3 and H3K9me3 in the sarcoma PDX are over-methylated compared to normal muscle. We evaluated the methylation status of histone marks H3K4me3 and H3K9me3 from the myxofibrosarcoma, UPS and liposarcoma grown in their corresponding PDX models. The tumor size at the end of the experiment was approximately 1,700 mm3. Histone H3K4me3 and H3K9me3 were over-methylated in all three sarcomas, compared to muscle tissue from the host mice, as seen by immunoblotting (Figure 1). Over-methylation of H3K4me3 in all three sarcoma models was also demonstrated by immunohistochemistry (Figure 2).
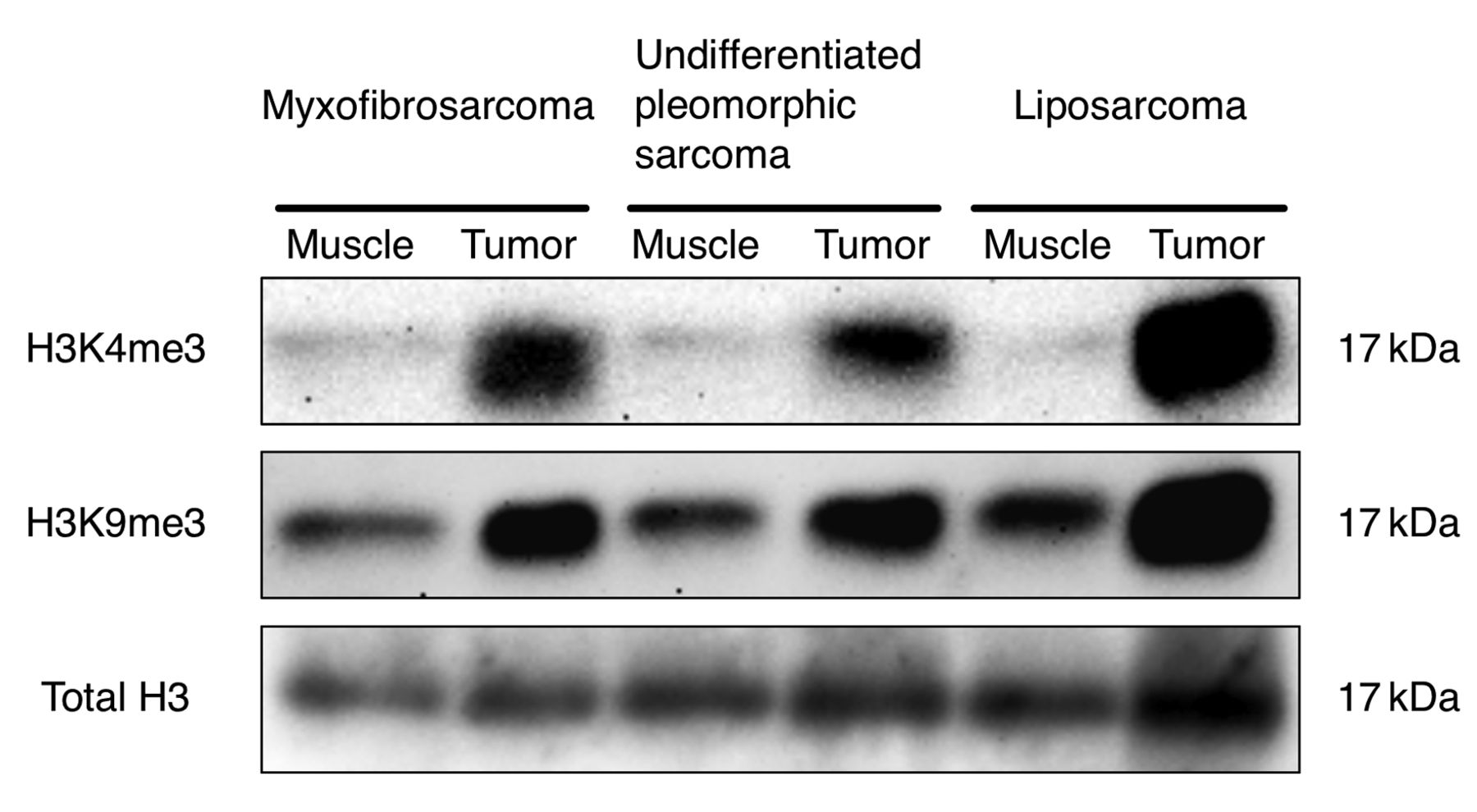
Immunoblotting of histone marks H3K4me3 and H3K9me3 from myxofibrosarcoma, undifferentiated pleomorphic sarcoma and liposarcoma and normal mouse muscle in patient-derived xenograft (PDX) models. Please see Materials and Methods for details.

(A) Representative hematoxylin and eosin (H&E) staining images of myxofibrosarcoma, undifferentiated pleomorphic sarcoma and liposarcoma grown in patient-derived xenograft (PDX) mouse models. (B) Immunohistochemical staining for the histone mark H3K4me3 in myxofibrosarcoma, undifferentiated pleomorphic sarcoma and liposarcoma grown in PDX mouse models, and normal mouse muscle. Magnification: 100×. Scale bar: 50 μm. Please see Materials and Methods for details.
Histology of each sarcoma type in the PDX model. H&E staining showed that all three types of sarcomas have each maintained the histological characteristics of their original sarcoma type after growth in PDX mouse models (Figure 2). The myxofibrosarcoma PDX tissue comprised high-density spindle-shaped cancer cells with myxoid stroma. The UPS PDX tissue comprised high-density pleomorphic cells. The liposarcoma PDX tissue comprised both well differentiated and dedifferentiated cells.
Discussion
The present study showed that histone marks H3K4me3 and H3K9me3 were over-methylated in myxofibrosarcoma, UPS and liposarcoma, growing as PDX nude mice, in contrast to normal muscle from the host mice, which was only slightly methylated in histone H3K4me3 and H3K9me3 (Figure 1). This result is consistent with our previous report that histone H3K4me3 and H3K9me3 are over methylated in methionine-addicted cancer cell lines, but not in normal cells or in low-malignancy methionine-independent revertants derived from the methionine-addicted cell lines (6, 7). This is the first report, to our knowledge, showing that histone H3K4me3 and/or H3K9me3 are over-methylated in the 3 major types of sarcomas, suggesting a potential common mechanism of malignancy in apparently heterogeneous types of sarcoma.
Methionine addiction has been found in all properly-tested cancer cell types in vitro (15, 16) and in vivo (17). The present results of over-methylated histone H3 marks in the three sarcoma PDX types indicate they also may be methionine addicted. The present report and our previous reports (6, 7) suggest that over-methylation of histone H3K4me4 and H3K9me3 may be a general biomarker in cancer, since histone H3 lysine over-methylation is linked to methionine addiction, which is found in all major cancer types (15, 16).
In recent years, there have been a few published reports on histone H3 lysine marks methylation in soft tissue sarcoma. In one case, an isocitrate dehydrogenase (IHD) gene mutation altered gene expression of SOX9, COL2A1, CDKN1C, and ALPL, which was associated with an increase in both histone H3K4me3 and H3K9me3 methylation in chondrosarcoma cell lines and mesenchymal stem cells (18, 19). A de-differentiated liposarcoma was found to have higher levels of methylation of histone H3K9me3 compared to well-differentiated liposarcoma (20).
Histone H3K4me3 and H3K9me3 are thought to be global gene regulators and their over-methylation status may alter their function and change the expression of a large number of genes that result in malignant conversion of the cell. The over-methylation of H3K4me3, which is involved in gene activation, and H3K9me3, which is involved in gene repression, is a further indication of unbalanced transmethylation in cancer, a classical concept (21). Such unbalanced methylation of histones may have profound effects and importantly contribute to the malignant phenotype (6).
Future studies will correlate the malignant behavior, including survival or metastasis of the sarcomas, with over-methylation of histone H3 lysine marks, in patient-derived orthotopic xenograft (PDOX) models (22). Future studies will also use other methods to investigate epigenetic modifications of histones in cancer, such as ChIP (23). The over-methylation of H3K4me3 and H3K9me3 found in the 3 major types of sarcoma in the present study may be related to methionine addiction, a fundamental hallmark of cancer (24–28), termed the Hoffman effect (29–31). The over-methylation of these histone marks required excess methionine in the form of S-adenosylmethionine and may, at least in part, account for the excess methionine required by cancer cells and is a basis why methionine restriction with recombinant methioninase has been an effective therapy for sarcoma in all major types of sarcoma PDOX models (32–46).
Acknowledgements
The study was supported, in part, by the Robert M. Hoffman Foundation for Cancer Research, which had no role in the direction of the research. This paper is dedicated to the memory of A. R. Moossa, MD, Sun Lee, MD, Professor Li Jiaxi and Masaki Kitajima, MD.
Footnotes
Authors’ Contributions
YA, JY, YT and RMH were involved in study conception and design. YA, JY, KH, NM and RMH were involved in acquisition of data. YA, JY, YT, KH, NM, SI, MB, IE, KN and RMH analyzed and interpreted data. YA, YT and RMH wrote this manuscript. All Authors reviewed and approved the manuscript.
This article is freely accessible online.
Conflicts of Interest
The Authors declare that there are no potential conflicts of interest.
- Received July 22, 2021.
- Revision received August 21, 2021.
- Accepted September 7, 2021.
- Copyright© 2021, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Reduction of Tumor Biomarkers from very High to Normal and Extensive Metastatic Lesions to Undetectability in a Patient With Stage IV HER2-positive Breast Cancer Treated With Low-dose Trastuzumab Deruxtecan in Combination With Oral Recombinant Methioninase and a Low-methionine Diet
- Extensive Synergy Between Recombinant Methioninase and Eribulin Against Fibrosarcoma Cells But Not Normal Fibroblasts
- The Combination of Methioninase and Ethionine Exploits Methionine Addiction to Selectively Eradicate Osteosarcoma Cells and Not Normal Cells and Synergistically Down-regulates the Expression of C-MYC
- Deletion of MTAP Highly Sensitizes Osteosarcoma Cells to Methionine Restriction With Recombinant Methioninase
- Oral-recombinant Methioninase Converts an Osteosarcoma from Methotrexate-resistant to -sensitive in a Patient-derived Orthotopic-xenograft (PDOX) Mouse Model
- Extent and Instability of Trimethylation of Histone H3 Lysine Increases With Degree of Malignancy and Methionine Addiction