


Abstract
Background/Aim: Germline mutations in PTCH1 or SUFU in the sonic hedgehog (SHH) pathway cause Gorlin’s syndrome with increased risk of developing SHH-subgroup medulloblastoma. Gorlin’s syndrome precludes the use of radiotherapy (a standard component of treatment) due to the development of multiple basal cell carcinomas. Also, current SHH inhibitors are ineffective against SUFU-mutated medulloblastoma, as they inhibit upstream genes. In this study, we aimed to detect differences in the expression of genes and microRNAs between SUFU- and PTCH1-mutated SHH medulloblastomas which may hint at new treatment directions. Patients and Methods: We sequenced RNA and microRNA from tumors of two patients with germline Gorlin’s syndrome – one having PTCH1 mutation and one with SUFU mutation – followed by bioinformatics analysis to detect changes in genes and miRNAs expression in these two tumors. Expression changes were validated using qRT-PCR. Ingenuity pathway analysis was performed in search for targetable pathways. Results: Compared to the PTCH1 tumor, the SUFU tumor demonstrated lower expression of miR-301a-3p and miR-181c-5p, matrix metallopeptidase 11 (MMP11) and OTX2, higher expression of miR-7-5p and corresponding lower expression of its targeted gene, connexin 30 (GJB6). We propose mechanisms to explain the phenotypic differences between the two types of tumors, and understand why PTCH1 and SUFU tumors tend to relapse locally (rather than metastatically as in other medulloblastoma subgroups). Conclusion: Our results help towards finding new treatable molecular targets for these types of medulloblastomas.
Basal cell nevus syndrome, also known as Gorlin syndrome (GS) increases the risk of developing odontogenic jaw keratocysts, skeletal abnormalities (1, 2), basal cell carcinoma (BCC) of the skin [in areas that have undergone radiation (1)], and medulloblastomas (MBs) by the age of five years (2). The most common cause of GS is a heterozygous germline mutation in the Patched (PTCH1) gene (3), but germline mutations in the SUFU gene have also been found to cause GS (4). Up to 5% of individuals with GS develop childhood SHH-MB (4); therefore, children with germline mutations are recommended to undergo periodic MRI screening of the brain until the age of eight years (5).
Both PTCH1 and SUFU are vital players in the activation of the sonic hedgehog (SHH) pathway, which is one of the main trafficking networks that regulate events during embryonic development, and aberrations in its regulation may cause congenital disabilities and cancer. Activation of the SHH signaling pathway is mediated by the receptor Smoothened (SMO). When the SHH ligand is low or absent (“off-state”), SMO transports to the membrane, where its activity is inhibited by Patched (PTCH1). The downstream effectors are inhibited via SUFU, resulting in inhibition of target gene expression. When SHH binds to Patched (“on-state”), SMO levels increase, and SUFU is deactivated, leading to activation of gene expression, resulting in cell growth and the patterning of multicellular embryos (6).
One of the four subgroups of MB – the most common malignant brain tumor in children (7) – is the SHH subgroup, which is most frequent in infants (<3 years old) and young adults (>16 years old). Mutations in PTCH1 or SUFU are frequent in the tumors of infants with SHH-MB (8). Although most of these mutations are sporadic, SUFU and PTCH1 germline mutations can be detected in 2% of all patients with MB, exclusively in the SHH subgroup (9). The risk of developing MB has been suggested to be 20 times higher in germline SUFU mutations and at a younger age than germline PTCH1 mutations (4, 10). MBs with a germline SUFU mutation show poor prognosis with overall survival rate of 66% (10), which is much lower than the >90% overall survival rate reported for desmoplastic MB in young children (11). These children often demonstrate local relapse with progression-free survival of 42% at five years, and they will most likely need radiation for salvage therapy (10).
Standard MB therapy for children over three years old includes surgical resection, upfront craniospinal irradiation, chemotherapy, and high-dose chemotherapy with hematopoietic stem cell rescue in high-risk patients (12). Due to the enormous cognitive damage of radiation in infants, treatment is usually based on chemotherapy alone. However, some children will relapse or progress and would need subsequent radiation therapy. Children undiagnosed with GS will develop hundreds to thousands of BCCs in the irradiated areas. Therefore, it is vital to identify those children with germline PTCH1/SUFU mutations to avoid irradiation at all costs.
Recently, new SHH inhibitors have been developed for the treatment of SHH-MB (13). However, these are SMO inhibitors and, therefore, will only inhibit the upstream activation of the pathway – e.g., at the level of PTCH1 or SMO – and will not affect downstream mutations, such as SUFU (14). Also, the SMO inhibitors cause irreversible growth plate fusion in children and, therefore, clinical studies are employing these agents for skeletally mature children only (15). There is a desperate need for new therapies for young children with GS-SHH-MB that should avoid radiation therapy, and, in particular children with germline SUFU mutation, who will not respond to SMO inhibitors.
We aimed to find new potential molecules such as microRNAs (miRs), to serve as diagnostic biomarkers or as drug targets (16, 17). miRs are short noncoding RNAs, which play an essential role in gene translational regulation. Moreover, miRs can be used to define specific signatures for individual cancers and cancer stages (18, 19), including for MB subgroup classification (20).
In this study, we searched for targetable pathways in the tumors of two patients diagnosed with SHH-MB, one with a germline SUFU mutation and the other with a germline PTCH1 mutation. We aimed to detect similarities and differences at the expression levels of genes and miRs to better understand the biology of these two tumors that could help develop targets for future clinical use.
Patients and Methods
Patients and tumor collection. The study design adhered to the tenets of the Declaration of Helsinki and approved by the institutional and national review board of the Israel Ministry of Health. Informed consent was obtained. Primary tumor samples were collected at surgery, placed in RNAlater™ (AM7020; Thermo Fisher Scientific, Waltham, MA, USA), and stored at −80°C.
RNA and microRNA extraction and sequencing. Total RNA was extracted from freshly frozen tumor tissue samples as previously described (20). Library preparation and sequencing was performed using the Illumina TruSeq protocol on the HiSeq 2500 machine. Raw data deposited at the Sequence Read Archive (SRA) accession number SRP095882. The PTCH1-MB was included in our previous study (SRS1888277) (20) while SUFU-MB is newly deposited (SRS3694085).
RNA-seq data analysis. Raw reads were processed and analyzed as previously described (20). To obtain dispersion estimates for a count dataset, we used the ‘estimateDispersions’ function in the DESeq R package (21). Since we had no replicates, we defined the argument method=“blind”, which ignored the sample labels and computed the empirical dispersion value of the gene as if the two samples were replicates of a single condition. The argument sharing Mode was defined as “fit-only”. Genes with FDR corrected p<0.05 were noted as displaying different expression levels.
MicroRNA-seq data analysis. Raw reads were processed and analyzed as previously described (20). Unless specified otherwise, a p-value of 0.05 was used as the significance cutoff.
Ingenuity pathway analysis (IPA). Genes with FDR<0.05 and miRs with p<0.05 were uploaded to QIAGEN’s Ingenuity® Pathway Analysis (IPA®, QIAGEN Inc., https://www.qiagenbioinformatics.com/products/ingenuity-pathway-analysis) software (22). The IPA was used to gain insights into the overall biological changes introduced by the expression, miR target gene prediction, and miR and gene Integrated Analysis. Using the Ingenuity Pathways Knowledge Base, each gene was linked to specific functions, pathways, and diseases.
Analysis of an independent microarray data. The dataset GSE85217 (23) downloaded from the Gene Expression Omnibus (GEO) database (24), comprises 763 samples of which 223 are SHH. There is no information regarding germline mutations in the database, hence we had to chose the samples with the highest probability of representing GS patients. The likelihood of developing MB in patients with GS is higher in younger children (2), and as we were interested in SHH-MBs that are as similar as possible to those examined in our study, we first selected tumors from children under three years of age. Then we selected only those with a deletion in 10q, which includes the SUFU gene (n=3), and those with a deletion in 9q, which contains the PTCH1 gene (n=13). In this way we can compare tumors with similar mutations to ours, even if this is just in the tumor and not in the germline. We employed a moderated t-test, conducted using the limma (25) R package (version 3.38.3). Deletions, histology, and age were included as covariates in the linear model, and an FDR-corrected p-value of 0.05 was used as the significance cutoff.
Cell culture and siRNA transfections. Daoy cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM; Biological Industries Israel, Beit Haemek; 01-052-1A) with 4.5 g/l D-glucose, 4 mM L-glutamine, 10% fetal bovine serum (Biological Industries Israel; 04-007-1A), and 1% penicillin/streptomycin. Dicer-substrate 27mer short interfering RNAs (DsiRNAs) targeting human SUFU (hs.Ri.SUFU.13.1, hs.Ri.SUFU.13.2) and PTCH1 (TriFECTa RNAi kit, hs.Ri.PTCH1.13) were pre-designed by and purchased from IDT. The cells were transfected with siRNA at a final concentration of 30 nM per siRNA (SUFU siRNA, PTCH1 siRNA, or Scramble siRNA for control) by using the Avalanche® Everyday Transfection Reagent (EZT-EVDY-1), according to the manufacturer’s protocol. Briefly, the cells were passaged one day before transfection to reach a confluency of 60-70%. The next day, the selected siRNA was incubated in a serum-free medium with the recommended volume of transfection reagent for 20 min at room temperature. The transfection mixture was gently added to the prepared cell culture plate(s) for continued incubation at 37°C for 24-36 h, until harvesting and RNA extraction.
Reverse transcription (RT) and quantitative PCR (qPCR). Total RNA was isolated from Daoy cells by using the NucleoZol homogenizing reagent (Machery-Nagel 740404.200), according to the manufacturer’s protocol. Purified RNA samples were reverse-transcribed using the GoScript Reverse Transcription System (Promega, Madison, WI, USA, A5000) according to the manufacturer’s protocol. The cDNA product was diluted 1:5 and mixed with SYBR Green PCR Master Mix (ThermoFisher Scientific) for amplification on an AriaMX thermal cycler (Agilent Technologies, Santa Clara, CA, USA) using the gene-specific primer sets described below. Each qPCR reaction had a total volume of 12 μl. Three biological replicates were performed, and all reactions were run in triplicates. The comparative Ct method was used to analyze mRNA levels, using actin as the normalization control.
Primers used for qPCR reaction were: SUFU 5’ CAGCA AACCTGTCCTTCCACCA 3’ CAGATGTACGCTCTCAAGCTGC, PTCH1 5’ GCTGCACTACTTCAGAGACTGG 3’ CACCAGG AGTTTGTAGGCAAGG, MMP11 5’ GAGAAGACGGACCT CACCTACA 3’ CTCAGTAAAGGTGAGTGGCGTC, FOXL2 5’ CGGAGAAGAGGCTCACGCTGT 3’ CTGAGGTTGTGGCGGAT GCTAT, GSTM1 5’ CTATGATGTCCTTGACCTCCACCGTATA 3’ ATGTTCACGAAGGATAGTGGGTAGCTGA, GABRA4 5’ TC CTGGACAGTTTGCTCGATGG 3’ CAGAAACAGGTCCAAAG CTGGTG, NPNT 5’ GTAAGCACAGGTGCATGAACA 3’ GA ACCATCCGGCATGAGCATA.
Results
Patients. Primary tumor samples were obtained from two children diagnosed with non-metastatic, desmoplastic MB. Both tumors were identified as belonging to the SHH subgroup by using nanoString nCounter Technology, as previously described (26).
Patient 1: A boy, the first child of Yemenite origin parents who are not relatives, with no family history of cancer. Neonatal follow-up showed large head circumference with continued growth on the 98% percentile. There was some delay in motor and speech development. The boy was noted to have torticollis, unilateral dilation of the renal pelvis, and trivial pulmonary stenosis. At age 22 months, he presented a two-week history of recurrent falls followed by vomiting and apathy. MRI at diagnosis (Figure 1B and C) showed a large heterogeneous mass in the cerebellar vermis with obstructive hydrocephalus. No metastatic spread to the craniospinal axis was evident. The patient underwent gross total removal of the tumor, and pathology showed desmoplastic medulloblastoma (Figure 1F). Genetic testing was performed due to macrocephaly and showed a de novo germline mutation in PTCH1 NM_000264.5(PTCH1):c.379G>T (p.Glu127*) (Figure 1A). The boy was treated according to the COG 99703 protocol without irradiation. He is currently 10 years old, with no evidence of relapse. He developed keratogenic jaw cysts at the age of 5 years and has mild learning difficulties. He has palmar pits and multiple melanocytic nevi.

(A) Sequence chromatograms, showing the heterozygous PTCH1 de novo germline mutation NM_000264.5(PTCH1): c.379G>T (p.Glu127*) in patient 1 (p1), and the non-carrier mother (m) and father (f) of the patient, as well as a control from an unrelated individual (con). (B-E) MRI images at diagnosis, showing the tumor (short, thick arrows) and severe hydrocephalus (long, thin arrows): (B-C) patient 1, PTCH-MB, (D-E) patient 2, SUFU-MB. (B) Axial T2WI image and (C) sagittal T1WI post-Gadolinium image, showing a large 3.2×4.5×4.4 cm heterogeneous mass in the cerebellar vermis on T2, invading the 4th ventricle. The mass is enhanced and has areas of restriction and necrosis with secondary obstructive hydrocephalus. (D) Axial T2WI image and (E) sagittal T1WI post-Gadolinium image, showing extreme hydrocephalus caused by the large 3.5×6×6 cm mass in the posterior fossa (PF) and surrounding edema. The tumor was localized centrally in the PF, with the involvement of the superior portion of the 4th ventricle with cranial extension. The tumor is multicystic with multiple enhancing tumor nodules and restricted on diffusion. (F-G) H&E preparation from the PTCH-MB tumor of patient 1 (F), showing nodular, reticulin-free zones surrounded by densely packed undifferentiated cells with hyperchromatic nuclei producing a dense intercellular reticulin network. Macrophages and stroma were inconspicuous. The preparation from the SUFU-MB tumor of patient 2 (G) shows a prominent lobular architecture, large reticulin-free zones, and is rich in neuropil-like tissue. These zones showed population by cells with neurocytic differentiation. Macrophages and stroma were inconspicuous.
Patient 2: A girl, the fourth child of unrelated parents of Iraqi-Moroccan/Yemenite origin, with no family history of malignancy. At age 3 months, the local well bay clinic noticed increasing head circumference, new-onset strabismus, and lethargy. MRI (Figure 1D and E) showed extreme hydrocephalus and a multicystic mass in the posterior fossa (PF). There were no metastases visible in the brain or spine. She underwent partial resection of the mass, leaving a supratentorial residue. The pathology result was MB with extensive nodularity (MBEN) (Figure 1G). The tumor was positive for both GAB1 and YAP in more than 80% of cells. Due to her extremely young age, she underwent genetic testing, which showed a de novo heterozygous loss of exon 3 in the SUFU gene at the DNA level. She was treated according to the ACNS 1221 protocol (before suspension of enrolment) without intrathecal chemotherapy. She is now 6 years and four months old, with no evidence of relapse. She shows some residual mild ataxia and dysmetria and attends a regular kindergarten. She has no other physical findings of GS.
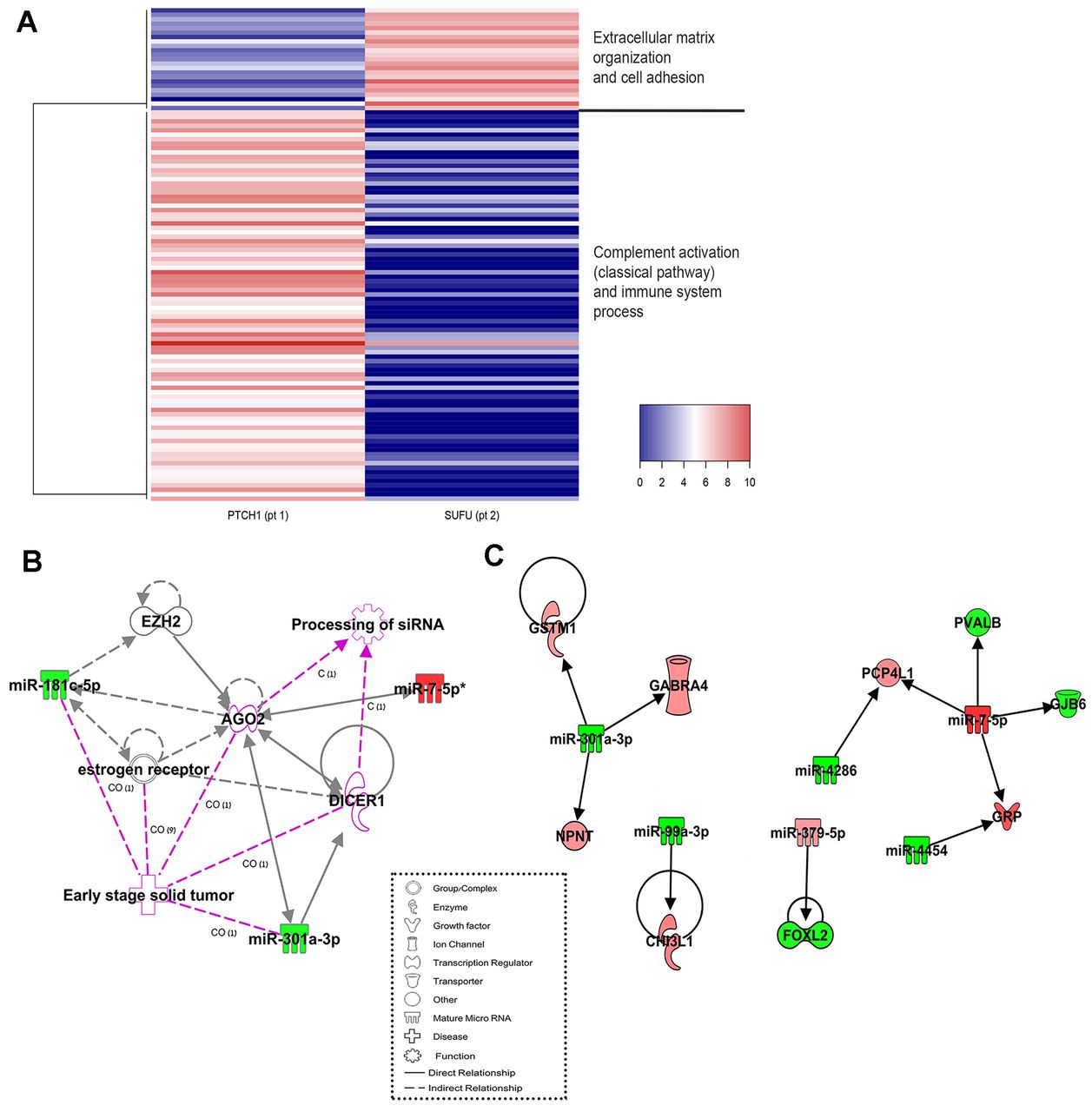
Expression profiling of genes. Comparison of SUFU-MB against PTCH1-MB detected 111 genes displaying different expression levels. Of these 23 were up-regulated, and 88 were down-regulated in SUFU-MB, compared to PTCH1-MB (Figure 2A). The up-regulated cluster was associated with extracellular matrix organization and cell adhesion, while the down-regulated cluster was associated with complement activation (classical pathway) and immune system processes. Of the MB-related genes, we detected a significantly lower expression of OTX2 in the SUFU-mutated tumor. OTX2 plays an essential role in normal cerebellar development (27) and aberrant OTX2 expression is implicated in several malignancies (28), including MB (29). Indeed, lower OTX2 expression is one of the biomarkers used to differentiate SHH-MB from other MB subgroups in a real-time PCR assay panel (30). In MB, the effect of overexpressed OTX2 as an oncogene is predominantly observed in Group 3/4, and participates in tumor localization and migration (28). In contrast, in SHH tumors, overexpression of OTX2 inhibits tumor progression (27). The lower expression of OTX2 may contribute to the increased risk of local relapse and, therefore, to poorer event-free survival which has been found in SUFU-mutated patients (10).

(A) A Heatmap of 23 up-regulated genes (red) and 88 down-regulated genes (blue) in the SUFU-MB compared to the PTCH1-MB. Heatmap was generated with heatmap3 (51) using complete as the linkage method, and Euclidean as the distance. (B) MicroRNAs expression in network no. 1 (Table III). (C) Gene and miR inverse expression in the SUFU-MB as compared with the PTCH1-MB. Green nodes represent decreased expression and red nodes represent increased expression. White nodes represent no significant change in expression. Pink line indicates diseases or functions related to the molecules. CO: Correlation, C: causation/leads. The figures were generated using Ingenuity® Pathway Analysis.
Recently, mTORC1 signaling was detected as a downstream effector of OTX2 in Group 3 MB (31). A relationship between mTOR pathway and SHH was also demonstrated for SHH MB (32, 33). However, the mTOR signaling molecules did not show differential expression between the two patients, demonstrating that mTOR probably does not have a role in regulating the pathophysiology differences of the two SHH tumors.
We found a decreased expression of matrix metallopeptidase 11 (MMP11) in SUFU-MB, compared to PTCH1-MB. MMPs are endopeptidases, responsible for the degradation of extracellular matrix components (34) and play a significant role in cancer (35). Increased expression of some MMPs, including MMP11, are correlated with the tumor WHO-grading classification of human malignant gliomas (36). It will be of interest to test whether the expression of MMPs contributes to the increased probability of local relapse in SHH tumors.
Immune signaling pathways dominated the list of genes whose expression differed between the tumors from the two patients (Table I).
Top five canonical signaling pathways identified by the ingenuity pathway analysis (IPA) for the genes whose expression differs between the two patients.
Over-represented diseases and biological functions included skeletal and muscular disorders, cell death and survival, embryonic development, and nervous system development and function (Table II).
Top diseases and biological functions identified by the ingenuity pathway analysis (IPA), for the genes whose expression differs between the two patients.
Top associated network functions included cancer-related functions, such as cellular development, DNA replication, recombination, and repair (Table III).
Top five associated network functions identified by the ingenuity pathway analysis (IPA) for the genes whose expression differs between the two patients.
Genes belonging to these networks include OTX2 and MYC, which are part of the expression panel used to differentiate MB subgroups (30). The genes GSTM1 and HLA-G, which belong to network number 2 (Table III), are known to be associated with BCC. Genetic variants in GSTM1 might contribute to the variation in the number of BCC jaw cysts and its presentational phenotypes in patients with PTCH-mutated as opposed to SUFU- mutated GS (37), while the expression of the HLA-G gene is high in BCC and decreases following radiotherapy (38).
Expression profiling of microRNAs. Overall, 778 miRs were expressed in both tumors, of which the expression level of 11 miRs was different between SUFU-MB and PTCH1-MB, three miRs displayed higher expression and eight lower expression levels (Table IV). IPA analysis of differentially expressed miRs identified cancer-related functions, such as cell morphology, cellular development, and cellular growth and proliferation as top diseases and biological functions (Table V).
MicroRNAs displayed different expression levels SUFU-MB compared to those expressed in PTCH1-MB.
Top diseases and biological functions identified by the ingenuity pathway analysis (IPA) for the miRs represented different expression in our patients.
Top network functions included RNA post-transcriptional modification and cancer, cardiovascular diseases, and connective tissue disorders (Table VI). Among the miRs that showed a lower expression in SUFU-MB were miR-301a-3p and miR-181c-5p. These miRs are related to early stages of solid tumor processes and the processing of siRNA networks, and they regulate, either directly or indirectly, the expression of DICER1 (Figure 2B).
Top three associated network functions identified by the ingenuity pathway analysis (IPA) for the miRs represented different expression in our patients.
Integrated analysis of miRs and mRNA expression. Of the 111 differentially regulated genes, 16 are targets for the 11 miRs. Of these, nine target genes demonstrated an inverse expression with six miRs (Table VII, Figure 2C). We found that miR-301a-3p demonstrates lower expression levels in SUFU-MB, compared to PTCH1-MB. miR-301a-3p acts as an oncomiR (39, 40), as it down-regulates the expression of the SMAD4 gene. Inhibiting miR-301a-3p reversed gemcitabine treatment resistance in pancreatic cancer cells in vitro by regulating the expression of PTEN (41). The role of miR-301a-3p in MB is yet unclear, and it may have different effects in different tissues. One of the target genes of miR-301a-3p is GABRA4, which is downregulated in MB (42). We found that the expression of GABRA4 was higher in SUFU-MB, possibly resulting from the lower expression of its regulator, miR-301a-3p. GABRA4 is a part of the GABA receptor signaling pathway and, considering our findings; it may not be downregulated in all MBs as previously thought. The specific role of GABRA4 in MB tumorigenesis, in general, or in SUFU-mutated MB is yet to be determined.
MicroRNAs and their targeted genes that exhibit inverse expression, predicted by ingenuity pathway analysis (IPA).
The expression level of miR-7-5p was higher in SUFU-MB than in PTCH1-MB and, correspondingly, the expression of its target gene, Connexin 30 (GJB6), was lower. Connexins play a role in the gap-junction signaling pathway, and they function as tumor suppressors (43). The expression of Connexin 30 in human glioblastoma cells was found to reduce their growth in vitro, but, at the same time, it made them resistant to the effects of radiation therapy (44). Increasing the levels of Connexin 30 in SUFU tumors may serve as a therapeutic option to decrease cell proliferation, while resistance to radiation therapy will be irrelevant in these young patients, whose up-front treatment is planned to be radiation-free.
MiR-379-5p showed higher expression in the SUFU-MB, compared to the PTCH1-MB, while its targeted gene, FOXL2, demonstrated lower expression levels. FOXL2 is a transcription factor involved in congenital disorders (45). It directly modulates the expression of the estrogen receptor 2 (ESR2) (46). A recent study found that 17β-estradiol, via ESR2, exerts chemoprotective effects in some MB cell lines (47). In the SHH pathway, SUFU and GLI interact and bind to PIAS1 (48), which activates estrogen receptors, including ESR2 (49). It may be instrumental to try and decrease the expression of miR-379-5p in SUFU-mutated tumors, which would increase the expression of FOXL2 and, therefore, the expression of ESR2, which may have chemoprotective effects.
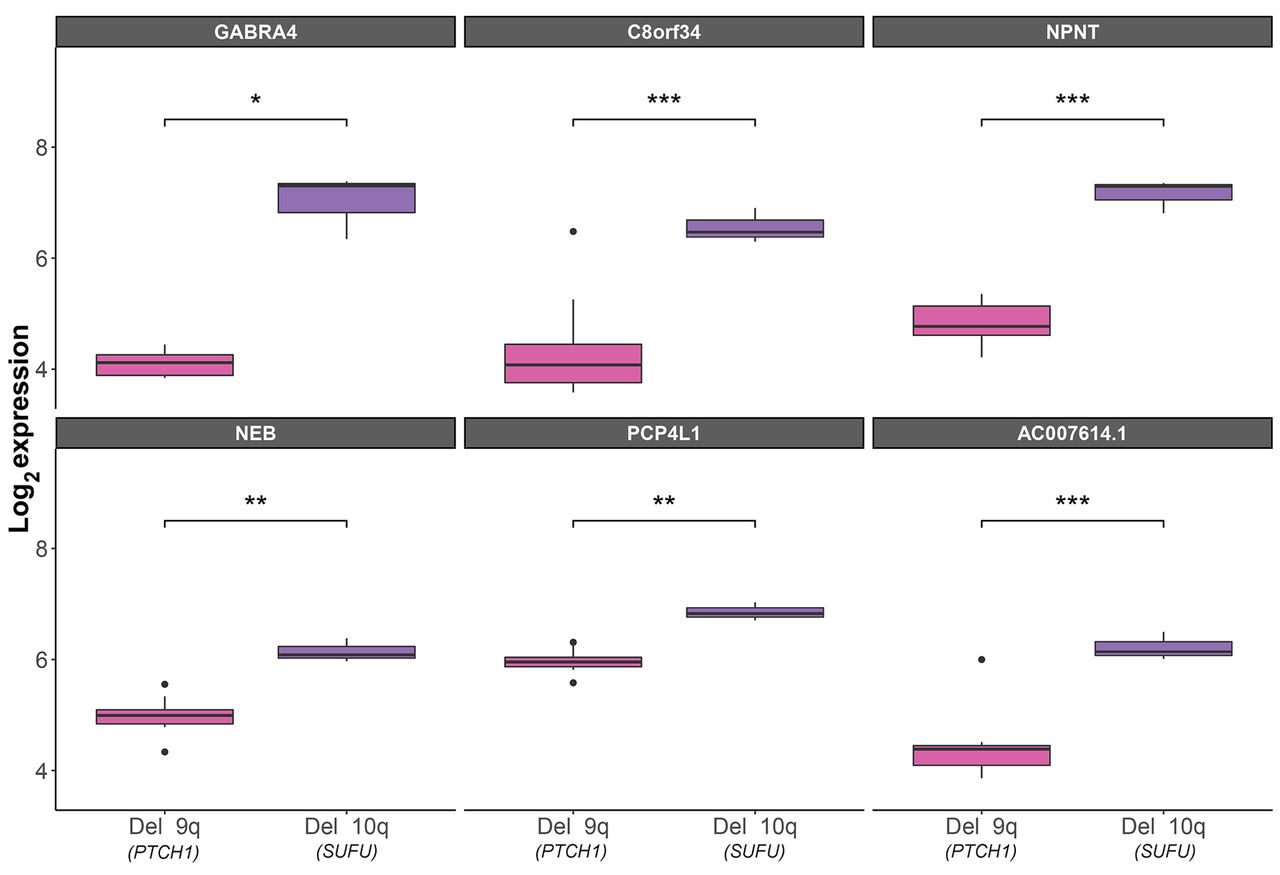
Supporting evidence from an independent cohort. Since the current study reports only two patients, it needs to be repeated in a larger cohort. Larger independent SHH cohorts of tumors with known germline mutations are not publicly available, but we were able to detect an independent dataset of SHH tumors. Although this independent dataset does not show which patients have a germline mutation (i.e., Gorlin’s syndrome), it does include important phenotypic data, such as age and chromosomal deletions. We know from the literature that the majority of SHH MBs occur in infants younger than 3 years (8). We chose to analyze data from these infants with tumors who carry a deletion in 10q or 9q (which include SUFU and PTCH1, respectively), adding significant support to our results. We assumed that the PTCH1- and SUFU-loss tumors will have a genetic expression similar to that in our patients. Of the 111 differentially expressed genes that we detected in our patients, 54 were also included in the microarray used in the GSE85217 dataset (available upon request). Of these, the expression of six genes (11%) was significantly different between PTCH1- and SUFU-loss tumors. These six genes are upregulated in the SUFU-loss tumors, which is in agreement with their expression in our SUFU-MB patient. Three of the six genes (GABRA4, NPNT, and PCP4L1) are targets that demonstrated an inverse expression with miRs in our patients (Figure 3).
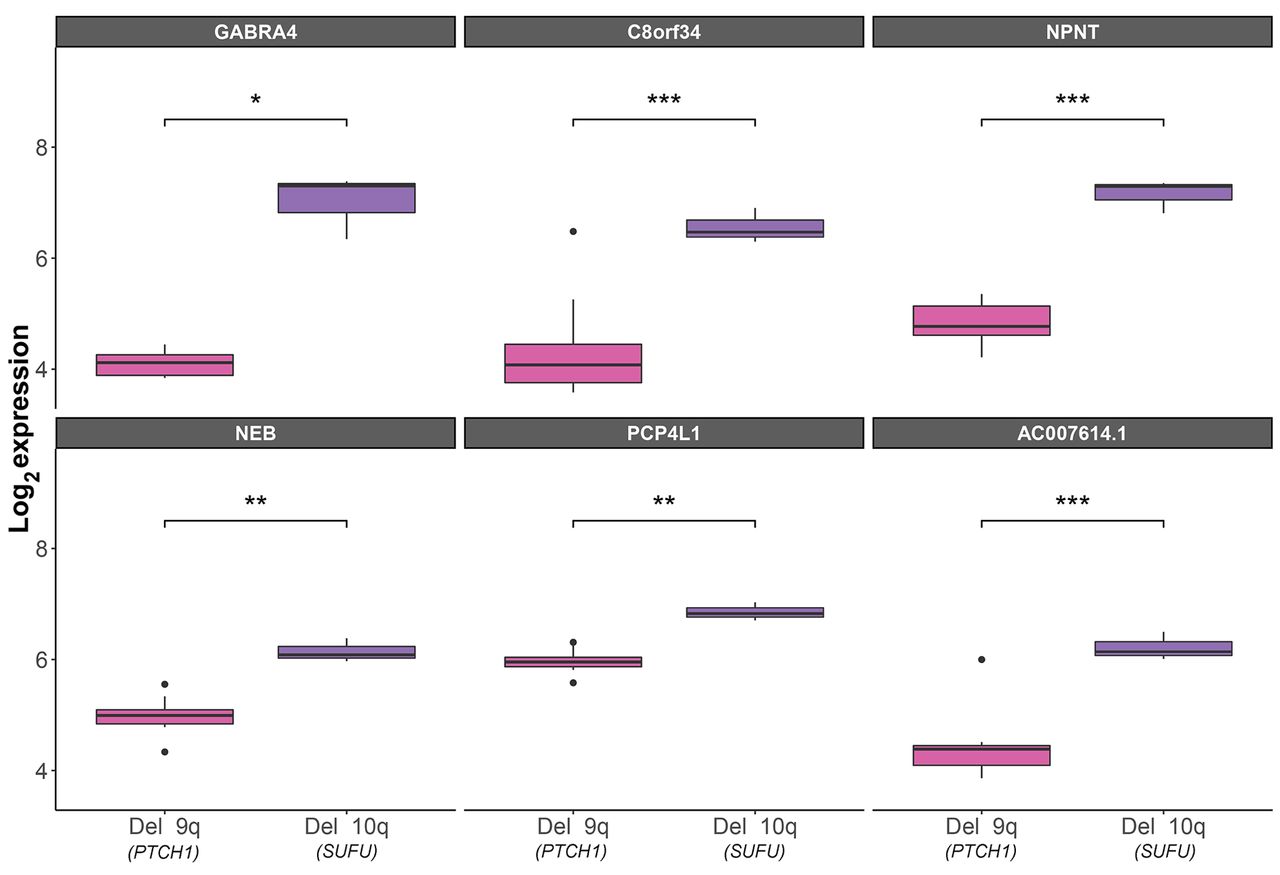
Box plots of differentially expressed genes in patients under the age of three, who carry a deletion in 10q, which includes the SUFU gene (n=3), or in 9q, which contains the PTCH1 gene (n=13), according to the independent GSE85217 dataset (23). t-Test statistical significance: *p≤0.05, **p≤0.01, ***p≤0.001.
In vitro validation. To validate our findings, we utilized short interfering RNA (siRNA) to knockdown SUFU or PTCH1 expression on a Daoy MB cell line, and we tested the effect of the expression of selected genes that were differentially expressed between PTCH1- and SUFU-loss tumors. To test knockdown efficiency, we quantified the PTCH1 and SUFU expression in transfected cells. The expression of PTCH1 and SUFU was reduced by 60% and 80%, respectively, in siRNA transfected cells (Figure 4A). Five genes were selected and their expression following PTCH1 and SUFU down-regulation was tested. The direction of change observed in the in vitro model was consistent with their change expression as observed in the PTCH1- and SUFU-loss tumors.
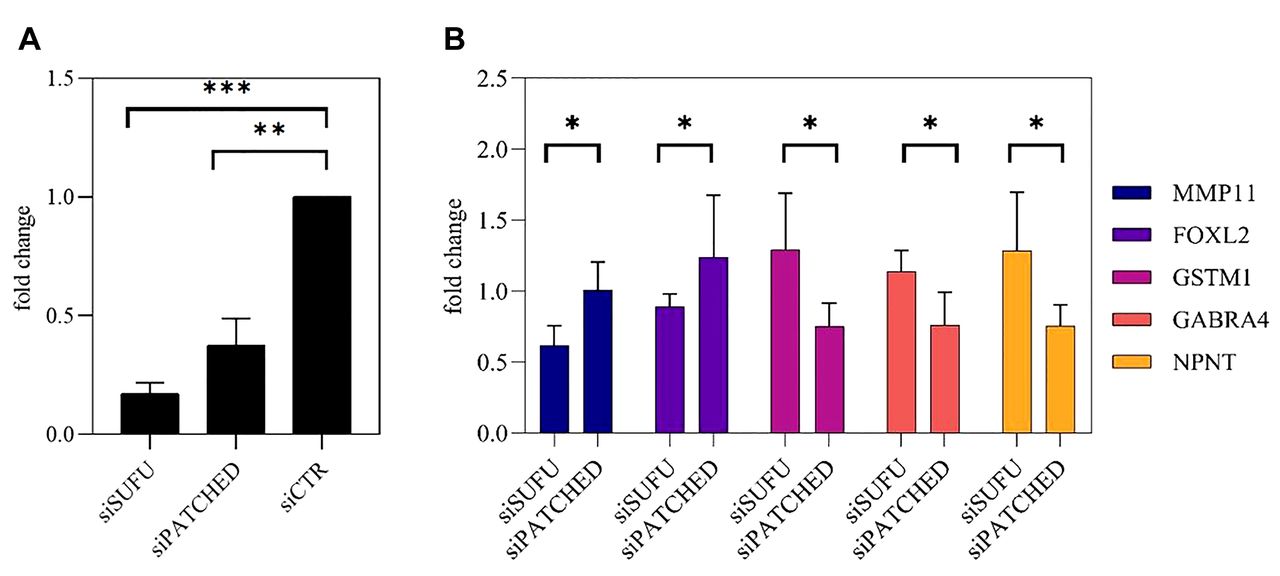
In vitro validation of gene expression. (A) Efficiency of SUFU and PTCH siRNA silencing. (B) SUFU and PTCH silencing influence on gene expression *p<0.05, **p<0.01, ***p<0.001 (one-tailed t-test), n=3.
Conclusion
Most MBs are thought to develop sporadically, but inherited forms also exist, most often in children with SHH MB. Due to the rarity of germline mutations that predispose to MB our knowledge of heritable predisposition is incomplete. Indeed, while 30% of all MB patients belong to the SHH subgroup, only 2% are defined as having Gorlin syndrome and show germline mutations in the SHH pathway. Hence, it is inherently difficult to collect samples from a large number of patients with such mutations. However, despite this caveat, by the detection of molecular differences between two SHH-MB tumors having different germline mutations, this study contributes to our understanding of the biology of heritable MB and suggested potential drug targetable pathways.
Treatment of MB usually involves chemotherapy and craniospinal irradiation with severe long-term effects on memory and cognition, growth and development, hearing and the risk of secondary malignancies. Much research is devoted to finding more successful and less damaging treatments for this disease. There are four subgroups of MB but the commonest in infancy is the SHH subgroup (50) of which approximately 20% will have germline mutations in PTCH1 or SUFU (Gorlin syndrome) (9). Children with GS should not receive radiation and general protocols omit/delay radiation for infants until they reach three years of age. Alternative treatments for infant with MB and in particular in the context of GS are desperately needed. Children with GS-SUFU differ phenotypically from those with GS-PTCH1 and even their MB differ with a poorer prognosis noted in the former and a higher rate of secondary malignancies (10).
Herein we explored the differences between the tumors from two infants both with SHH-MB that bear different germline mutations causing GS, and we used an unbiased whole-transcriptome sequencing to identify previously undetected potential therapeutic targets. Often the study of a rare genetic disease can have implications for research and treatment of a wider cohort of patients such as SHH-MB as a whole. In the same way that targeted therapy has been developed for PTCH1-mutated tumors (although, at the moment, relevant only for skeletally mature patients), we hope that a suitable target will be found for those with downstream mutations, such as SUFU and GLI and for infant with MB in general. This report may stimulate interest among the MB community and hopefully result in international collaborations for further delineating the unique features of different groups within MB and the SHH group in particular.
Acknowledgements
This research was funded by Zanvyl and Isabelle Krieger Fund (Baltimore, MD) (N.G-C), and the Israel Cancer Association (N.G-C), and the Levi-Eshkol Fund, Ministry of Science, Technology & Space, Israel (grant number 3-12624) for providing S.G scholarship. The Authors also wish to acknowledge Dr. Ruty Mehrian-Shai from Sheba Mediacl Center for providing the Daoy cell line.
Footnotes
* These Authors contributed equally to this work.
↵& Current address: Arthur and Sonia Labatt Brain Tumour Research Centre, Hospital for Sick Children, Toronto, ON, Canada.
↵Authors’ Contributions
Conceptualization, M.S-D and H.T; supervision, M.S-D, A.P, T.L and N.G-C; recruiting patients, H.T, S.M and N.G-C; clinical information, H.T and S.F; formal analysis and investigation, S.G; laboratory experiments, N.P; writing—original draft preparation, S.G, H.T and M.S-D; writing—review and editing, all Authors.
This article is freely accessible online.
Conflicts of Interest
The Authors declare no conflicts of interest.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon request.
- Received January 19, 2021.
- Revision received February 11, 2021.
- Accepted February 15, 2021.
- Copyright© 2021, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.