


Abstract
Background/Aim: Cancers are selectively sensitive to methionine (MET) restriction (MR) due to their addiction to MET which is overused for elevated methylation reactions. MET addiction of cancer was discovered by us 45 years ago. MR of cancer results in depletion of S-adenosylmethionine (SAM) for transmethylation reactions, resulting in selective cancer-growth arrest in the late S/G2-phase of the cell cycle. The aim of the present study was to determine if blockade of the MET-methylation axis is a highly-effective strategy for cancer chemotherapy. Materials and Methods: In the present study, we demonstrated the efficacy of MET-methylation-axis blockade using MR by oral-recombinant methioninase (o-rMETase) combined with decitabine (DAC), an inhibitor of DNA methylation, and an inhibitor of SAM synthesis, cycloleucine (CL). We determined a proof-of-concept of the efficacy of the MET-methylation-axis blockade on a recalcitrant undifferentiated/unclassified soft-tissue sarcoma (USTS) patient-derived orthotopic xenograft (PDOX) mouse model. Results: The o-rMETase-CL-DAC combination regressed the USTS PDOX with extensive cancer necrosis. Conclusion: The new concept of combination MET-methylation-axis blockade is effective and can now be tested on many types of recalcitrant cancer.
- Cancer
- methionine addiction
- methionine restriction
- methionine-methylation-axis blockade
- decitabine
- cycloleucine
- MAT2A
- soft-tissue sarcoma
- PDOX
Methionine (MET) addiction (1-14) is a fundamental and general hallmark of cancer discovered by us 45 years ago (3). MET addiction is observed in the clinic where the high demand for MET by cancers, results in a strong signal from [11C]-MET-PET imaging (2). MET addiction involves elevated MET flux in cancer cells (3-6) due to excess levels of transmethylation reactions (4), known as the Hoffman effect (7). MET addiction is a general phenomenon in cancer and a crucial target for cancer therapy by MET restriction (MR) which results in depletion of free MET and S-adenosylmethionine (SAM) (5, 6, 10) and selective S/G2-phase cell-cycle arrest in cancer cells (11-13). MET addiction is tightly linked to other hallmarks of cancer (14).
Recombinant methioninase (rMETase) targets MET addiction of cancer cells by severely depleting the sources of cellular MET (1, 15). We demonstrated the effectiveness of oral administration of rMETase (o-rMETase) on many types of chemotherapy-resistant cancers with patient derived orthotopic xenograft (PDOX) mouse models (15-19).
Decitabine [5-aza-2’-deoxycytidine, (DAC)], which is in clinical use for myelodysplastic syndrome and leukemia treatment, is a DNA-methylation inhibitor that incorporates irreversibly into DNA and causes DNA hypomethylation (20, 21). The efficacy of DAC alone for cancer treatment was unsatisfactory (22-26). We recently reported that the combination of o-rMETase and DAC inhibited a recalcitrant undifferentiated/unclassified soft-tissue sarcoma (USTS) PDOX model (27). Cycloleucine (CL), an inhibitor of MET adenosyltransferase-2A (MAT2A), which catalyzes SAM synthesis from MET and ATP, has been shown to lower intracellular SAM (28-30).
Soft-tissue sarcoma is a rare cancer which arises from mesenchymal cells (31). Chemotherapy is used in combination with surgery to treat soft-tissue sarcoma (32). Doxorubicin (DOX) has been used for soft-tissue sarcoma for the past 40 years, but with limited efficacy (31, 33, 34). USTS, formerly known as malignant fibrous histiocytoma, is a frequent type of soft-tissue sarcoma seen in the middle-aged and the elderly (35). The outcome of USTS is usually unsatisfactory when the tumor is resistant to DOX (33, 34).
Based on MET addiction due to excess transmethylation reactions, we hypothesized that CL could be combined with o-rMETase and DAC for MET and methylation blockade for highly effective therapy of the USTS PDOX model. The present report is a proof-of-concept that MET-methylation-axis blockade can effectively target a recalcitrant cancer.
Materials and Methods
Mice. Athymic nude mice, at 4-6 weeks of age, were from AntiCancer Inc. (San Diego, CA, USA). An IACUC protocol was approved for the present study following the principles and procedures described in the National Institutes of Health Guide for the Care and Use of Animals under Assurance Number A3873-1 (36). All surgical procedures were conducted under appropriate anesthesia and analgesia (36).
Patient-derived tumor. A fresh surgical sample from the USTS patient not otherwise specified (NOS), who underwent surgery at UCLA, was previously brought to AntiCancer, Inc. for establishment in nude mice (34). The patient provided written informed consent with UCLA IRB#10-001857 approval (34).
Surgical orthotopic implantation (SOI). A single tumor fragment (2-3 mm), harvested from a subcutaneously-grown tumor in nude mice, was implanted into the nude-mouse biceps, to establish USTS PDOX models, as described in our previous reports (27, 31).
Treatment protocols. The PDOX mouse models (n=6/group) were treated as follows (Figure 1): G1, Untreated control; G2, CL (50 mg/kg, intraperitoneal injection, twice per week); G3, o-rMETase (50 units/mouse, oral gavage, twice per day); G4, DOX (3 mg/kg, intraperitoneal injection, once per week); G5, CL+ DAC (50 mg/kg, intraperitoneal injection, once per day) ; G6, o-rMETase + CL + DAC. Treatment was initiated after all tumors reached at least 100 mm3. Tumor volume and mouse body weight were measured twice per week.
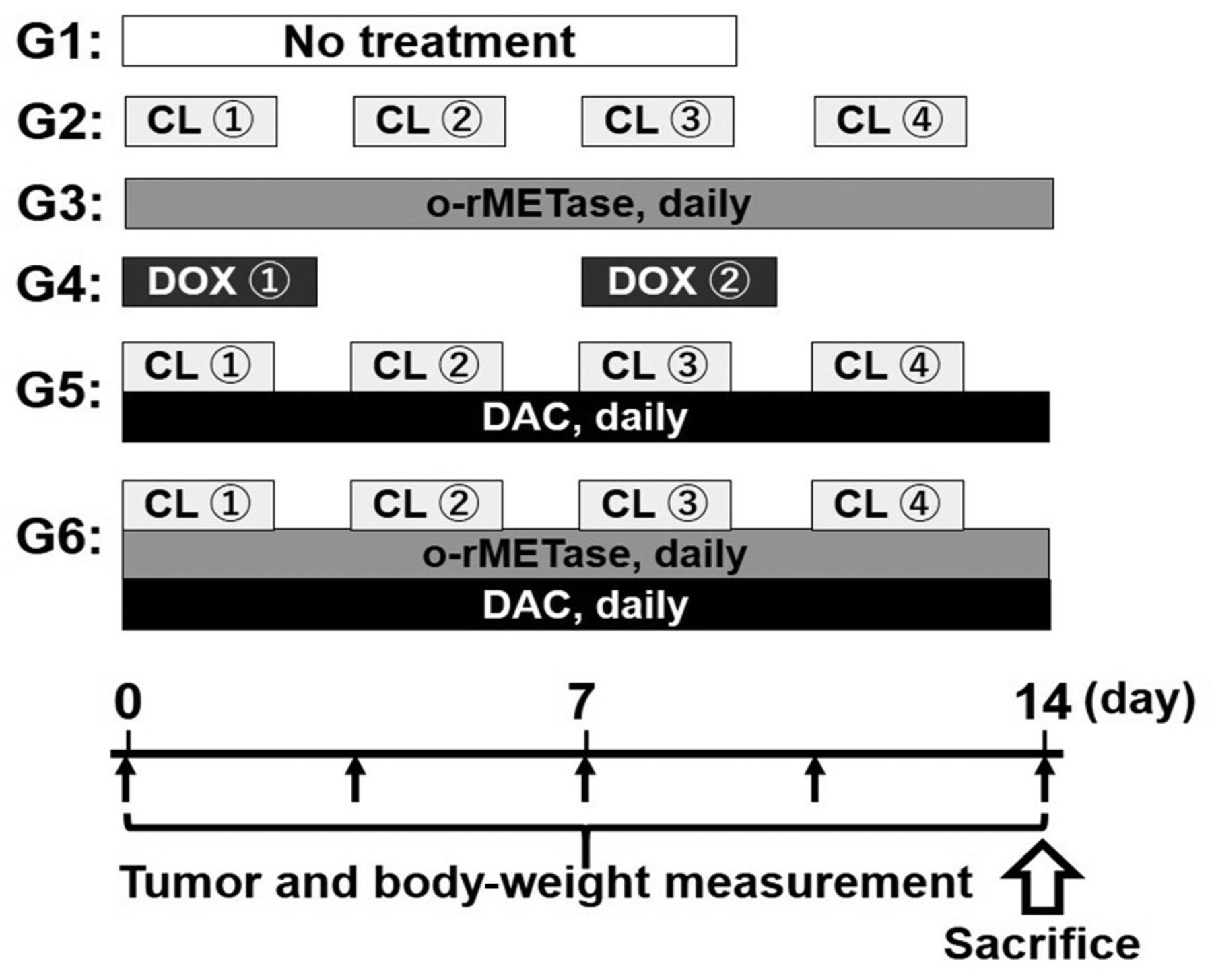
Treatment schema. DOX: Doxorubicin; o-rMETase: oral recombinant methioninase; DAC: decitabine.
Histological analysis. The procedures for fixation, sectioning, deparaffinizing, and staining of harvested tumor samples were conducted as described previously (37).
Statistical analysis. Data are presented as mean±standard error of the mean (SEM). One-way ANOVA with Tukey’s range test and the Student’s paired t-test were used for statistical analyses.
Results
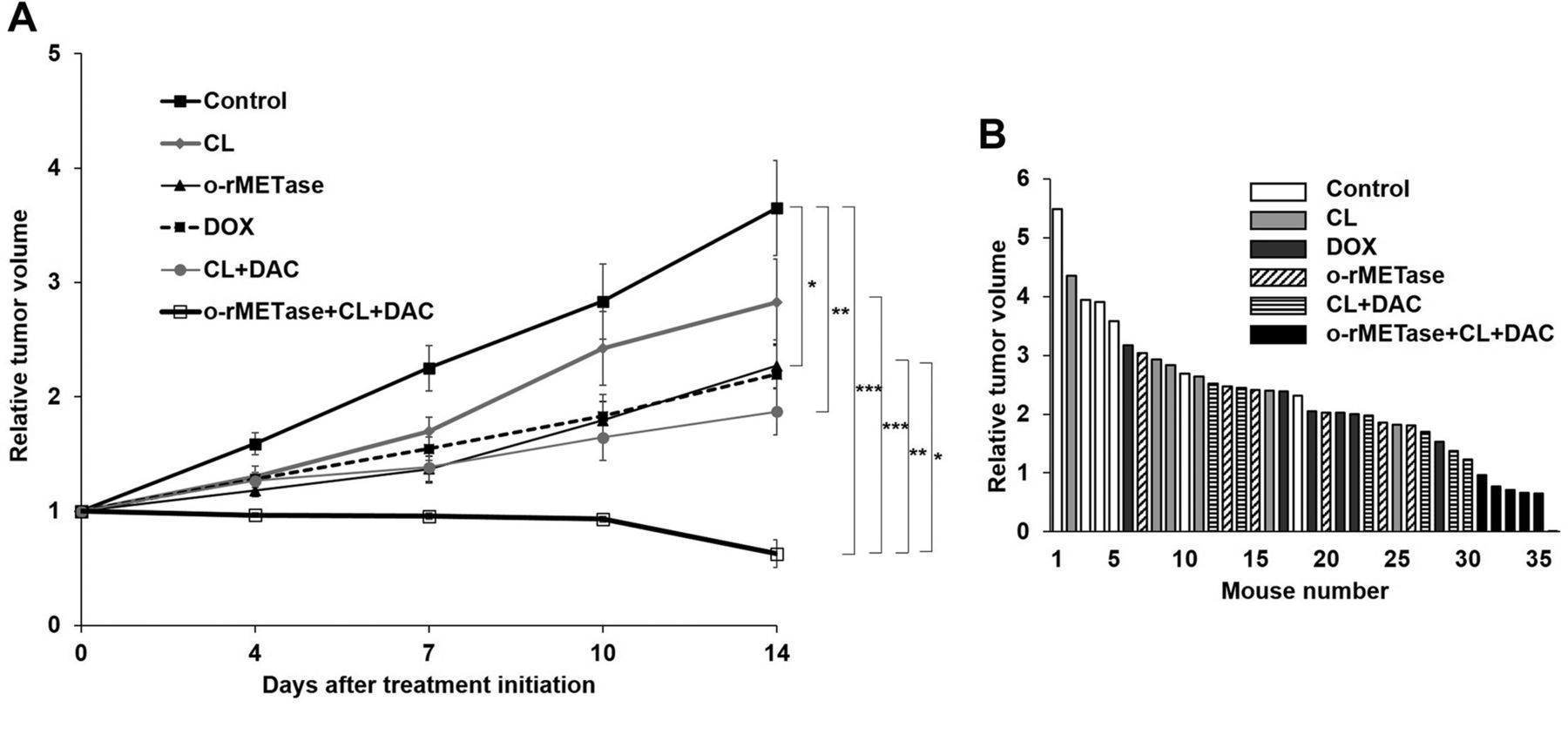
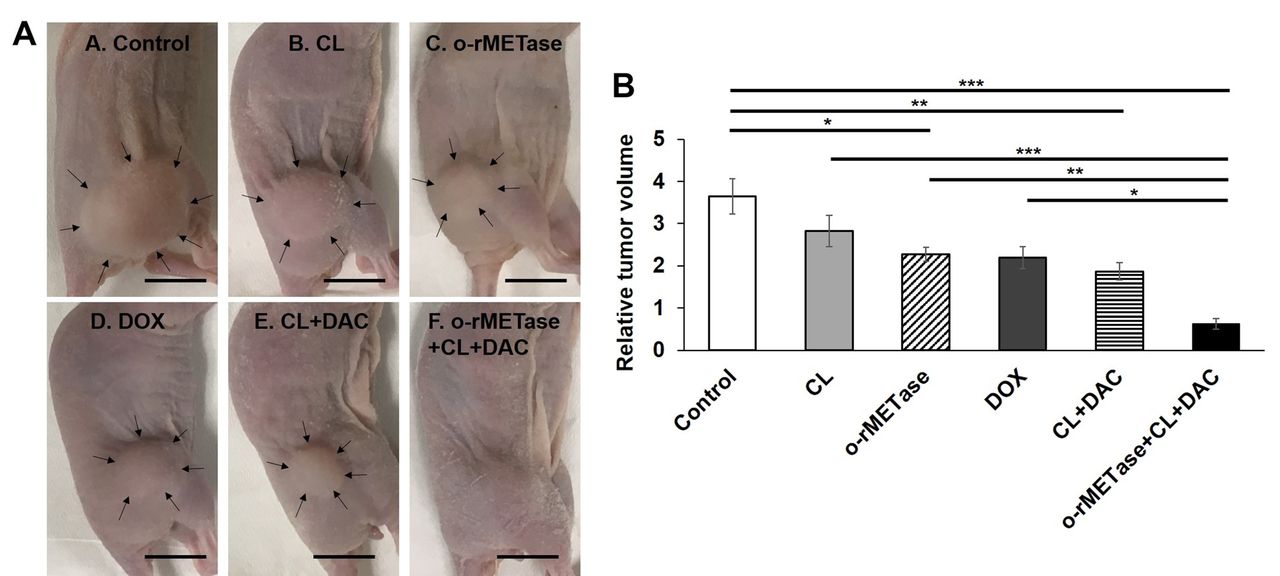
MET-methylation-axis blockade regressed the sarcoma PDOX. Only the MET-methylation-axis blockade regressed the recalcitrant USTS PDOX tumor (p<0.001). o-rMETase alone (p=0.03), the CL-DAC combination (p=0.003), as well as the rMETase-CL-DAC combination MET-methylation-axis blockade (p<0.001) significantly inhibited the USTS PDOX tumor compared to the control. The o-rMETase-CL-DAC combination had significantly better efficacy than the other treatments: vs. CL alone (p=0.001); vs. o-rMETase alone (p=0.007); or vs. DOX alone (p=0.03). CL alone did not inhibit the tumor (p=0.44) (Figures 2 and 3).
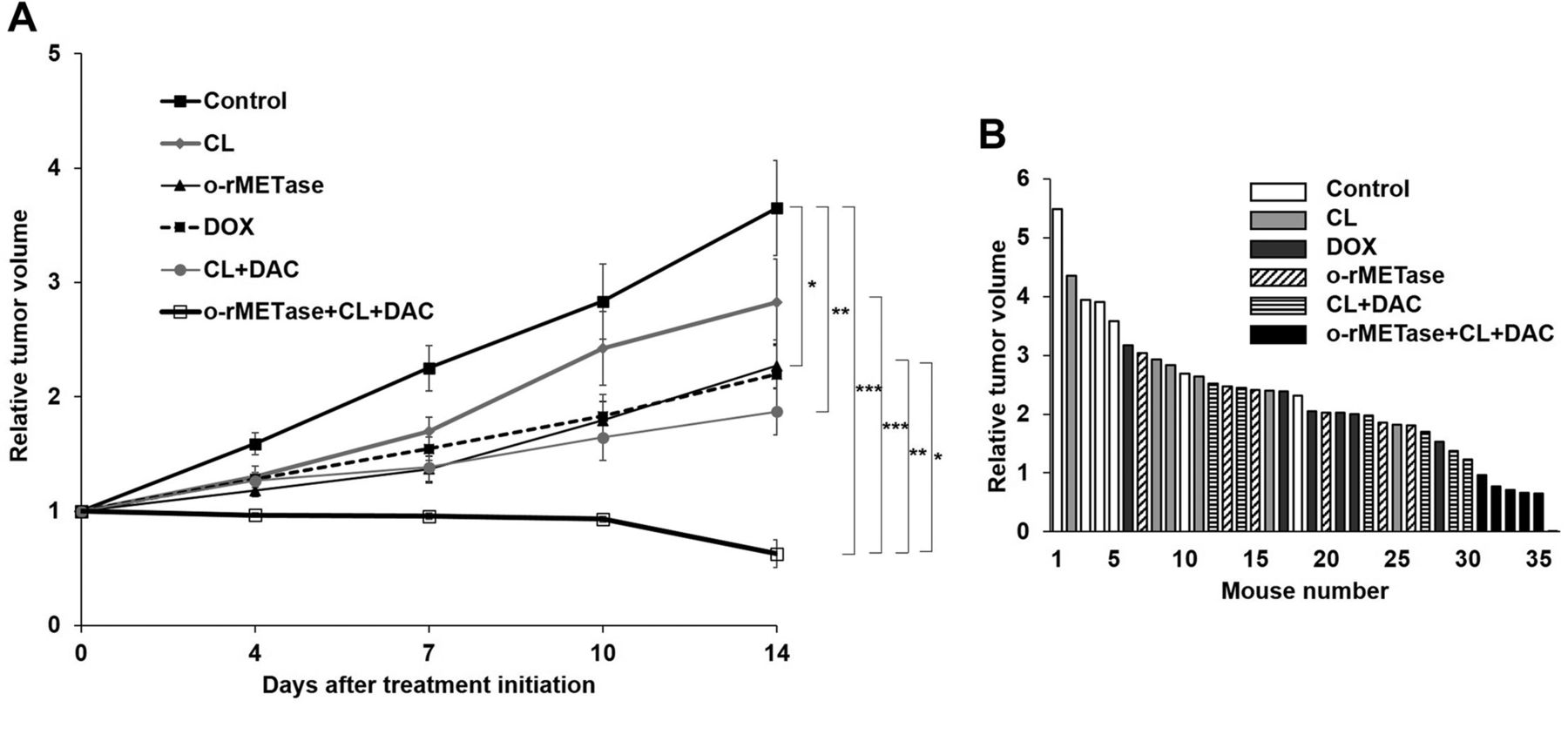
(A) Relative tumor volume of the USTS-PDOX model. (B)Waterfall plot of relative tumor volume. Six mice were in each group. *p=0.05; **p=0.01; ***p=0.001. Error bars: ±SEM. USTS, Undifferentiated/unclassified soft-tissue sarcoma; PDOX, patient-derived orthotopic xenograft; o-rMETase, oral recombinant methioninase; CL, cycloleucine; DOX, doxorubicin; DAC, decitabine; SEM, standard error of the mean.

(A) Representative photographs of treated USTS PDOX mouse models on day 14. Arrows show the margin of the tumors. (B) Bar graphs indicate relative tumor volume of each group on day 14. Scale bars: 10 mm. 6 mice were in each group. *p=0.05; **p=0.01; ***p=0.001. Error bars: ±SEM. USTS, Undifferentiated/unclassified soft-tissue sarcoma; PDOX, patient-derived orthotopic xenograft; o-rMETase, oral recombinant methioninase; CL, cycloleucine; DOX, doxorubicin; DAC, decitabine; SEM, standard error of the mean.
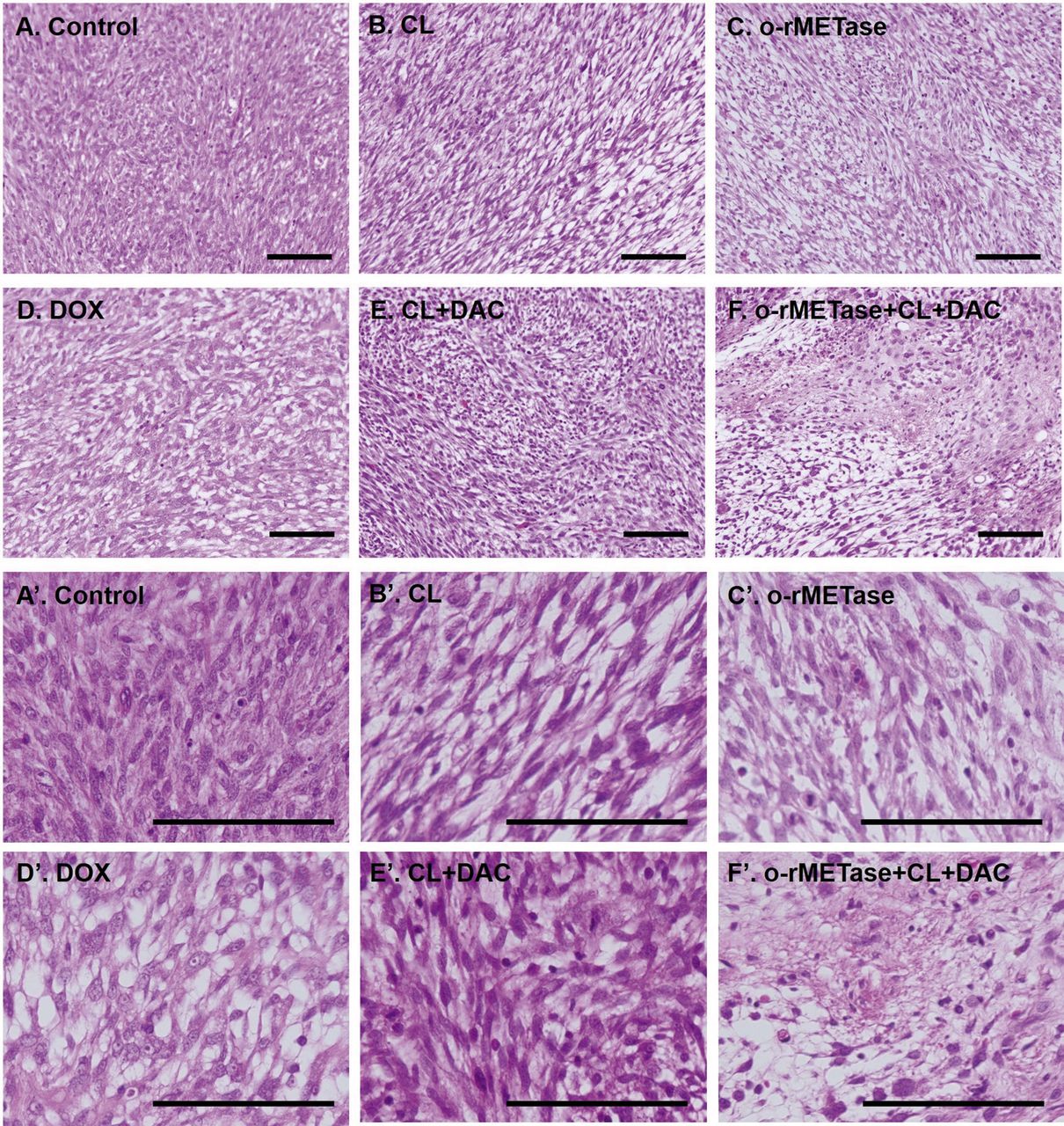
The combination MET-methylation-axis blockade caused extensive necrosis in the sarcoma PDOX. Only the PDOX tumors treated with the MET-methylation-axis blockade had extensive tumor necrosis. The control USTS PDOX tumor had a high cell density comprising atypical spindle-shaped cancer cells (Figure 4A, A’). USTS PDOX tumors treated with CL alone (Figure 4B, B’), o-rMETase alone (Figure 4C, C’), DOX alone (Figure 4D, D’), and the CL-DAC (Figure 4E, E’) combination showed viable pleomorphic cancer cells; however these groups had less viable cancer cells than the untreated control. The o-rMETase-CL-DAC combination induced widespread necrosis (Figure 4F) with non-viable cancer cells and stroma replaced by degenerative scars (Figure 4F’). These results further showed the efficacy of MET-methylation-axis blockade.

H&E staining. (A, A’) Control. (B, B’) CL alone. (C, C’) o-rMETase alone. (D, D’) DOX alone. (E, E’) CL+DAC. (F, F’) o-rMETase+CL+DAC. Scale bars: 100 μm. o-rMETase, oral recombinant methioninase; DOX, doxorubicin; CL, cycloleucine; DAC, decitabine.
Effect of treatment on body weight. Only treatments containing CL caused body-weight loss: CL alone (p=0.001), the CL-DAC combination (p=0.001), and the o-rMETase-CL-DAC combination (p=0.001) (Figure 5).
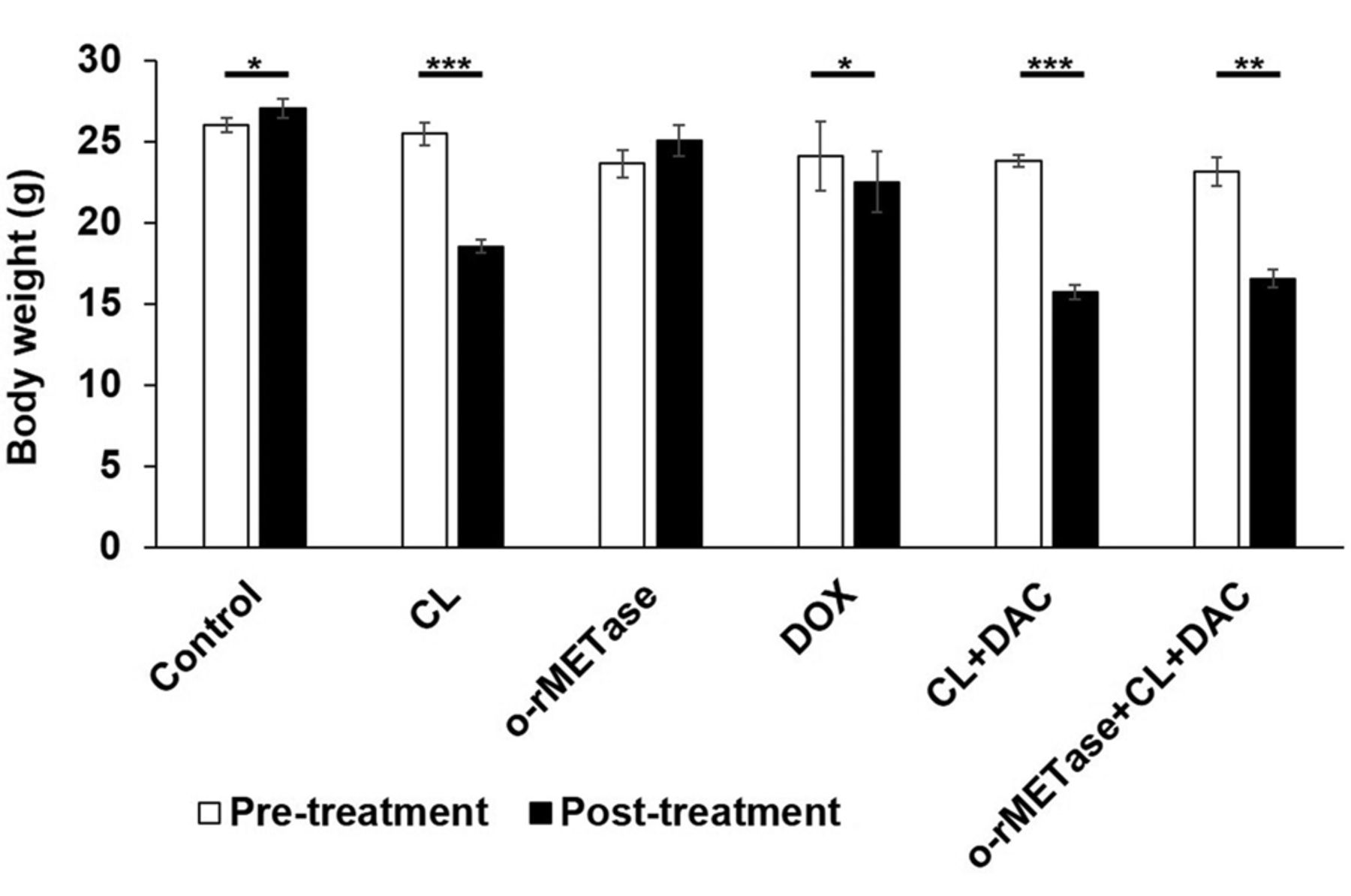
Mouse body-weight. *p=0.05; **p=0.01; ***p=0.001. Error bars: ±SEM. o-rMETase, oral recombinant methioninase; CL, cycloleucine; DOX, doxorubicin; DAC, decitabine; SEM, standard error of the mean.
Discussion
MET addiction of cancer is due to excess and aberrant transmethylation reactions, that consume much larger than normal amounts of MET (3-6, 10). Lowered DNA methylation is a hallmark of cancer cells, originally discovered in our laboratory (39), and may be due to diversion of methyl groups to other substances such as histones (please see below). Several demethylating agents have been used for cancer treatment (21, 40) including DAC, as well as azacitidine which has been used to treat myelodysplastic syndrome (41). These inhibitors cause hypomethylation of DNA, but had limited efficacy in the clinic (25, 40, 41). In our recent study, a DNA hypomethylating drug alone did not inhibit a USTS-PDOX, but o-rMETase combined with a DNA-hypomethylating drug arrested the PDOX tumors and decreased the cancer-cell density, suggesting the combination of rMETase and a DNA hypomethylating agent could be effective to inhibit tumor growth (27, 37).
In order to enhance MR, we examined the efficacy of the SAM-synthesis inhibitor, CL (30) along with o-rMETase and DAC in the present study. SAM, which plays an important role in transmethylation reactions in all cells, is synthesized from MET by MAT2A (28). CL inhibits SAM synthesis by inhibiting MAT2A, resulting in hypomethylation in the tumor and may increase the DNA hypomethylation efficacy of DAC (28, 30).
CL may be too toxic at the current dose, leading to mouse body-weight loss. Future studies will examine the most effective non-toxic dose of CL for optimal MET-methylation-axis blockade.
o-rMETase has now shown clinical efficacy in a pilot study (42). Recently, a study showing efficacy of a human enzyme engineered to be a METase was published, stating it is superior to the bacterial METase used in the present study, due to its long half-life in the circulation (43). However due to the hardiness of the bacterial enzyme to survive stomach acidity, it could be dosed orally as shown in the present and previous studies (15-19, 27, 37), giving it superiority over the injected engineered human METase. Bacterial o-rMETase is much superior to injected bacterial rMETase (44). Future studies will include determination of the efficacy of MET-methylation-axis blockade on the major cancer types in PDOX models. Recent papers have come out claiming novelty on MET addiction (45, 46), about which we published long ago (2-12, 47-50). Targeting a central aspect of metabolism such as MET and methylation has far more potential for cancer therapy than targeting peripheral metabolism, despite claims of “metabolic dependence” (51). MET addiction, discovered by us, is found in all cancer types and is linked to other hallmarks of cancer (14). All cancer types tested are sensitive to MR, whether by methionine-free media (8, 52), diet (47-49) or methioninase (15-19, 27, 37). A recent study in our laboratory (53) has demonstrated that the combination of o-rMETase, CL and azacytidine arrested a pancreatic cancer PDOX, further demonstrating that blocking the MET-methylation-axis is highly-effective chemotherapy against recalcitrant cancers and has clinical potential. Other recent studies from our laboratory have shown that excess transmethylation that causes MET addiction in cancer cells results in hypermethylation of histone H3 lysine marks (54, 55). These facts suggest MET addiction may be the very basis of cancer (3). Orthotopic mouse models of sarcoma (31) such as that used in the present study, are clinically relevant, unlike ectopic subcutaneous sarcoma mouse models (56). Recently, more potent inhibitors of MAT2A, which is regulated differently in normal and cancer cells (50), have been developed (57) and may be used in MET-transmethylation-axis blockade, which directly targets the elevated flux of methionine in cancer cells, termed the Hoffman effect (58).
Acknowledgements
This paper is dedicated to the memory of AR Moossa, MD, Sun Lee, MD, Professor Jiaxi Li, and Masaki Kitajima, MD.
Footnotes
Authors’ Contributions
Conception and design: TH and RMH. Acquisition of data: TH, QH, NS, JY, NY, KH, HK, SM, and KI. Analysis and interpretation of data: TH, NY, KH, HK, SM, MB, HT, and RMH. Writing, review, and/or revision of the manuscript: TH, HT, SRS, and RMH.
This article is freely accessible online.
Conflicts of Interest
The Authors declare no competing interests.
- Received August 16, 2020.
- Revision received December 31, 2020.
- Accepted January 19, 2021.
- Copyright© 2021, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- High Clinical Concordance of Drug Resistance in Patient-derived Orthotopic Xenograft (PDOX) Mouse Models: First Step to Validated Precise Individualized Cancer Chemotherapy
- Oral-recombinant Methioninase Converts an Osteosarcoma from Methotrexate-resistant to -sensitive in a Patient-derived Orthotopic-xenograft (PDOX) Mouse Model
- Over-methylation of Histone H3 Lysines Is a Common Molecular Change Among the Three Major Types of Soft-tissue Sarcoma in Patient-derived Xenograft (PDX) Mouse Models
- Lowering and Stabilizing PSA Levels in Advanced-prostate Cancer Patients With Oral Methioninase
- Oral-recombinant Methioninase Converts an Osteosarcoma from Docetaxel-resistant to -Sensitive in a Clinically-relevant Patient-derived Orthotopic-xenograft (PDOX) Mouse Model