


Abstract
Background/Aim: Metastasis to cervical lymph nodes of oral squamous cell carcinoma (OSCC) leads to a poor prognosis. The present study aimed at investigating the pathways and molecules associated with OSCC metastasis. Materials and Methods: The transcriptome between HSC-3 cells and their highly metastatic subline, HSC-3-M3 cells, was examined using gene expression microarray. Gene enrichment analyses and Ingenuity Pathway Analysis were performed. Kaplan-Meier plot analysis using a publicly available dataset was conducted to assess whether candidate molecules are prognosticators. Results: A total of 1,018 genes were differentially expressed, and the inflammatory pathway and NF-kB were predicted to be activated in HSC-3-M3 cells. CSF2 was suggested to be an indicator of poor prognosis in head and neck cancers. Conclusion: Inflammation and NF-kB may be involved in the metastasis of OSCC, and CSF2 is a promising diagnostic and therapeutic molecule. Moreover, HSC-3-M3 cells are a useful cell line model for studying OSCC progression.
Oral cancer is the most common epithelial malignancy among head and neck cancers, and its incidence has reportedly increased (1). It arises from the lining mucosa of the tongue, gingiva, buccal mucosa, oral floor, and hard palate, and most cases are histologically characterized as squamous cell carcinomas. In the treatment of oral squamous cell carcinomas (OSCC), surgical resection is the first choice, while radiotherapy and chemotherapy are also used depending on disease stage and patient condition. In recent years, the usage of molecularly targeted drugs and immune checkpoint inhibitors has become promising for advanced cases of OSCC (2).
Some clinical and pathological prognosticators can influence the survival of OSCC patients (3). Of these, lymph node metastasis is one of the most critical prognostic factors (4). Due to the rich lymph vessel plexuses in the head and neck region, OSCC often shows metastasis into the cervical lymph nodes, leading to metastasis to other organs that are further away and adding to a poor prognosis (4, 5). We have developed retrograde super-selective intra-arterial chemotherapy against advanced OSCC and showed its effectiveness in patients with cervical lymph node metastasis (6-9). However, despite the technological advances, the survival rate of patients with metastasis is still lower than that of the patients without it; therefore, it is clinically important to elucidate the molecular mechanisms of OSCC metastasis to improve the outcome.
Metastasis is a multi-step process involving the invasion of cancer cells into the surrounding tissues, intravasation, and proliferation at regional lymph nodes and distant organs (10). During these processes, dynamic phenotypic changes occur in cancer cells. Several factors such as transforming growth factor β (TGF-β) and tumor necrosis factor-α (TNF-α) stimulate cancer cells to activate intracellular signaling and gene-regulatory networks, leading to invasive and metastatic phenotypic changes such as Epithelial-Mesenchymal Transition (11).
Given that the expressions of numerous genes are dysregulated in the metastatic process, appropriate cancer cell line models are needed to reveal the intracellular molecular pathways and networks. Various cancer cell sublines with disease progressive phenotypes have been established from parental cells using in vitro or in vivo screening (12, 13). Because the genetic background between a parental cell and its subline is suggested to be similar, this cell line model would be useful in revealing specific pathways and molecules involved in cancer progression. We previously analyzed the transcriptome between the prostate cancer cell, LNCap, and its castration-resistant subline, C4-2B, and identified a novel tumor-suppressive, long non-coding RNA (lncRNA), DRAIC (14). Although some OSCC sublines have been established, few studies have examined their molecular characteristics (15-21).
HSC-3 cells are a tongue-derived OSCC cell line and are widely used for oral cancer research. The cells were originally isolated from the poorly differentiated OSCC of a 64-year old man (22). Matsui et al. established the highly metastatic subline from HSC-3 cells using in vivo screening in 1998 (15). They injected HSC-3 cells into the tongues of nude mice and the cells that metastasized into the lymph nodes were isolated and injected into the tongue again. After this procedure was repeated three times, the resultant cells were designated as HSC-3-M3 cells. When injected into the tongues of nude mice, HSC-3-M3 cells metastasize to cervical lymph nodes with a frequency of 90%, an incidence that is three times higher than that of parental HSC-3 cells (15). Although HSC-3-M3 cells have been used as a highly metastatic cell line model in a few articles (23, 24), to date, none have reported on the differences in mRNA transcriptome with parental HSC-3 cells. We hypothesized that comparing the transcriptome between HSC-3 and HSC-3-M3 cells would reveal specific intracellular pathways, gene-regulatory networks, and key molecules associated with OSCC metastasis.
In the present study, a gene expression microarray was utilized to compare the transcriptome between HSC-3 and HSC-3-M3 cells. We identified over 1,000 differentially expressed genes (DEGs) in these cells. Gene enrichment analyses showed that inflammation-related genes and pathways were enriched in these DEGs. Ingenuity Pathway Analysis (IPA) predicted the activation of NF-kB in HSC-3-M3 cells. Moreover, from a gene-regulatory network enriched with HSC-3-M3 cells, we discovered that CSF2 could potentially be used as a prognostic marker. Our results suggest that the inflammatory pathway and NF-kB may be involved in OSCC metastasis, and CSF2 is a promising diagnostic molecule and therapeutic target.
Materials and Methods
Cell culture. HSC-3 and HSC-3-M3 cell lines were obtained from the Japanese Collection of Research Bioresources (JCRB) Cell Bank (Osaka, Japan), and maintained in Dulbecco’s modified Eagle’s medium containing 10% fetal calf serum.
Short tandem repeat (STR) analysis and sex-typing. DNA was isolated from HSC-3 and HSC-3-M3 cells using the QIAamp DNA Mini Kit (QIAGEN, Redwood City, CA, USA). A polymerase chain reaction (PCR) was conducted using 10 ng of DNA and the GenePrint 10 System (Promega Corporation, Madison, WI, USA) according to the manufacturer’s instructions.
Microarray analysis for mRNA profiling. Total RNA was isolated using a TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA). RNA integrity number (RIN) was assessed by an Agilent Technologies 2100 Bioanalyzer (Agilent Technologies, Carpinteria, CA, USA), and RIN scores of all the samples used for the microarray analysis were 10. A SurePrint G3 Human Gene Expression v3 8x60K Microarray (Agilent) was performed. Quantile normalization was performed using R. The expression dataset was deposited into the National Center for Biotechnology Information Gene Expression Omnibus (GEO) (https://www.ncbi.nlm.nih.gov/geo/) and are accessible through GEO Series accession number GSE160395.
Gene enrichment and pathway analyses. Gene Set Enrichment Analysis (GSEA, v4.1.0) (25) was run according to the default parameters. The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was performed using Metascape (https://metascape.org/gp/index.html#/main/step1) (26). QIAGEN Ingenuity® Pathway Analysis (IPA®, QIAGEN, version 52912811) was used to find functional and significant pathways. IPA Upstream Analysis, Canonical Pathway Analysis, and Regulator Effect Analysis were performed and visualized according to the manufacturer’s instructions. The list of DEGs obtained from the microarray analysis (false discovery rate (FDR) adjusted p<0.05 and absolute fold change >1.5) was used for GSEA, KEGG pathway analysis, and IPA.
Kaplan-Meier plot analysis using a publicly available dataset. OncoLnc (http://www.oncolnc.org/) (27) was used to analyze the prognosis of head and neck squamous cell carcinoma patients of The Cancer Genome Atlas (TCGA) dataset (28). The patients were divided into two groups based on the expression of MMP3, CXCL8, PTGS2, CSF2, and MMP13 (high expression group: top 75%, low expression group: bottom 25%).
Results
Identification of the origin of HSC-3 and HSC-3-M3 cells. The cellular morphologies of parental HSC-3 and its highly metastatic subline HSC-3-M3 cells in monolayer culture are cuboidal, and their differences were not evident under phase-contrast microscopy (data not shown). We performed a short tandem repeat (STR) analysis to determine whether these cells originated from the same individual. STR is a DNA sequence where 2-6 base pairs are tandemly repeated. An individual inherits an allele from each of their parents. Therefore, the two alleles contain either the same repeat number (homozygote allele) or different numbers (heterozygote allele) in an STR marker. Nine STR markers on the maternal and paternal chromosomes were amplified using PCR that used genomic DNA isolated from HSC-3 and HSC-3-M3 cells. As shown in Table I, one repeat number was detected in D13S317, D7S820, D16S539, CSF1PO, and TPOX while different repeat numbers were detected in TH01, D21S11, D5S818, and vWA in both cells. We also analyzed the differences in DNA sequence length of the sex-typing marker, Amelogenin gene, in X and Y chromosomes by PCR. The results indicated that the sex chromosome constitution was XY in these cells (Table I). The STR profiling and sex-typing were completely matched between these cells, suggesting that they were derived from the same male individual, and not contaminated by other cell lines. It was also verified that profiling and sex-typing were the same as the data registered in Cellosaurus (https://web.expasy.org/cellosaurus/), which is an online knowledge resource on cell lines.
Short tandem repeat (STR) profiling and sex-typing in HSC-3 and HSC-3-M3 cells used in this study.
Transcriptomic analysis by gene expression microarray. We isolated the total RNA from these cells and performed a gene expression microarray. A total of 1,018 genes were differentially expressed in HSC-3 vs. HSC-3-M3 cells with a significant level of FDR-adjusted p-value <0.05 and absolute fold change >1.5. These differentially expressed genes (DEGs) are presented as a volcano plot in Figure 1A and a heat map in Figure 1B. The DEGs were analyzed by GSEA to identify hallmark gene sets. The most significant gene sets of gene ontology enriched with HSC-3-M3 and HSC-3 cells were “cytokine secretion” (Figure 1C) and “cell surface receptor signaling pathway involved in cell-cell signaling” (Figure 1D), respectively. The KEGG pathway analysis characterized some significant pathways such as “platinum drug resistance” and “TNF signaling pathway” in up-regulated DEGs in HSC-3-M3 cells (Figure 1E), and “inflammatory mediator regulation of transient receptor potential channels” and “primary immunodeficiency” in down-regulated DEGs in HSC-3-M3 cells (Figure 1F). In the TNF signaling pathway, a typical inflammatory pathway, inflammation-related genes such as CCL20, CSF2, IL18R1, MMP3, and PTGS2 were up-regulated in HSC-3-M3 cells (Figure 1G).
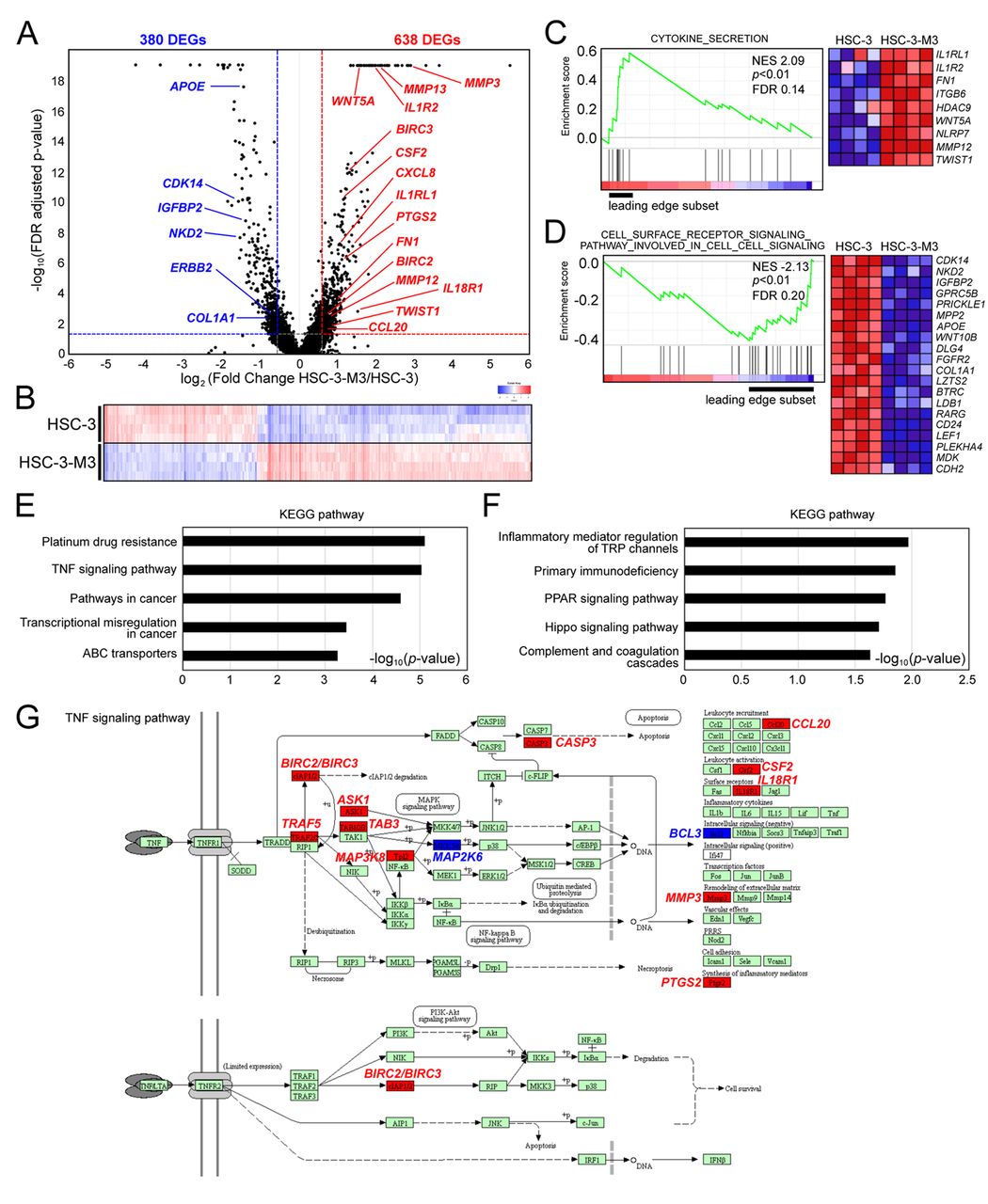
Gene expression profile and gene enrichment analyses of HSC-3 vs. HSC-3-M3 cells. (A) Volcano plot of log2 fold change (X-axis) and - log10 false discovery rate (FDR) adjusted p-value (Y-axis) of HSC-3 (four replicates) vs. HSC-3-M3 (four replicates) cells. The red and blue dotted lines indicate the threshold cutoff of absolute 1.5-fold change and FDR adjusted p-value 0.05. Genes with -log10 FDR adjusted p-value higher than 18 are shown at the top of the Y-axis. DEGs; differentially expressed genes. (B) Heat map representing DEGs of HSC-3 vs. HSC-3-M3 cells. Each column represents the relative expression level for individual DEGs. Increased gene expression is shown by a deeper red while decreased gene expressions by a deeper blue. (C) and (D) Histogram of Gene Set Enrichment Analysis (GSEA) of the enriched gene set in HSC-3-M-3 cells (C) and HSC-3 cells (D). Normalized enrichment score (NES), nominal p-value, and FDR are indicated. The heat map images show individual gene expression of the leading-edge subset. (E) and (F) Kyoto Encyclopedia of Genes and Genomics (KEGG) pathway analysis using up-regulated (E) or down-regulated (F) DEGs in HSC-3-M3 cells compared with HSC-3 cells. (G) TNF signaling pathway registered in KEGG database was visualized using KEGG Mapper (https://www.genome.jp/kegg/mapper.html). The molecules highlighted in red are up-regulated DEGs in HSC-3-M3 cells, and the molecules highlighted in blue are down-regulated DEGs in HSC-3-M3 cells.
Potential upstream regulators activated in HSC-3-M3 cells. An IPA was performed to determine the upstream regulators controlling the DEGs. The IPA Upstream Regulator Analysis predicts the potential upstream regulators using p-values. And z-scores >2 (activated state) or <2 (inhibited state) are considered statistically significant. The top 22 upstream regulators ranked by p-value are shown in Figure 2A. Among them, TNF (z-score=3.752), lipopolysaccharide (z-score=3.231), SMARCA4 (z-score=2.982), IL1B (z-score=2.664), FOS (z-score=2.838), RELA (z-score=2.725), EPAS1 (z-score=2.283), and NFkB (complex) (z-score=2.57) were predicted as activated regulators in HSC-3-M3 cells compared with HSC-3 cells. All these factors are well-known for their involvement in cytokine and inflammatory-related pathways. In particular, the RELA/NFkB (complex) is an inflammation-related transcriptional factor in numerous cancer progression conditions. In IPA Canonical Pathway Analysis, it was shown that some signal transducers such as IkB kinase (IKK) complex in NF-kB signaling were predicted to be activated in HSC-3-M3 cells (Figure 2B).

Involvement of NF-kB pathway in the establishment of gene expression pattern in HSC-3-M3 cells. (A) Upstream regulator analysis using IPA. The results were sorted by p-value in ascending order, and the top 22 ranked results are shown with activation z-score. The left Y-axis indicates -log10 (p-value) while the right Y-axis shows activation z-score. Upstream regulators with activation z-score >2, which means predicted activation in HSC-3-M3 cells, are highlighted in red. (B) Canonical pathway analysis using IPA. The illustration shows NF-kB signaling. The meanings of each shape and color are also presented.
Functional networks enriched in HSC-3-M3 cells. Functional networks enriched with HSC-3-M3 cells were analyzed next. The IPA Regulator Effect Analysis shows upstream regulators and gene sets from the DEGs, which theoretically induce significant diseases and functions. It calculates consistency scores, where higher scores are given in networks that are directionally consistent with published literature from the IPA library. The top five ranked networks were summarized in Table II. Among them, top-ranked network had 19 upstream regulators, 34 DEGs, and 5 diseases and functions. This outcome illustrated in Figure 3 indicated that in HSC-3-M3 cells, upstream regulators including NFkB1-RelA may control the 34 DEGs, followed by the cancer progressive phenotypes such as “Metastasis of tumor cell lines” and “Migration of cancer cells”. “Migration of smooth muscle cells”, “Cell movement of vascular smooth muscle cells,” and “Neoplasia of breast cancer cell lines” were also predicted. Other enriched functional networks also predicted several functions such as “immune response” and “production of reactive oxygen species” (Table II).

Prediction of the functional network enriched with HSC-3-M3 cells. The top-ranked network results analyzed by the regulator effect analysis of IPA are shown. The regulators (top) are predicted to be associated with the diseases and functions (bottom) via gene sets consisting of DEGs (middle) in HSC-3-M3 cells. The meanings of each shape and color are also shown below.
Top 5 ranked regulator effect based on consistency score calculated by ingenuity pathway analysis (IPA).
Survival analysis of five candidate mRNAs. Among the 34 DEGs in the top-ranked network shown in Figure 3, MMP3, CXCL8, PTGS2, CSF2, and MMP13 were predicted to be under the control of NFkB1-RelA and involved in the metastasis and migration of cancer cells. In our microarray analysis (Figure 1A and B), the fold changes of MMP3, CXCL8, PTGS2, CSF2, and MMP13 were 7.49, 2.14, 2.16, 2.22, and 4.37, respectively. Because these mRNAs were up-regulated in HSC-3-M3 cells, one would expect that a high expression of these mRNAs may predict a poor prognosis. A Kaplan-Meier plot based on the dataset from “Head-Neck Squamous Cell Carcinoma” including OSCC showed that a higher expression of CSF2 mRNA predicts a lower probability of overall survival although the p-values of other mRNAs did not reach statistical significance (Figure 4A-D).
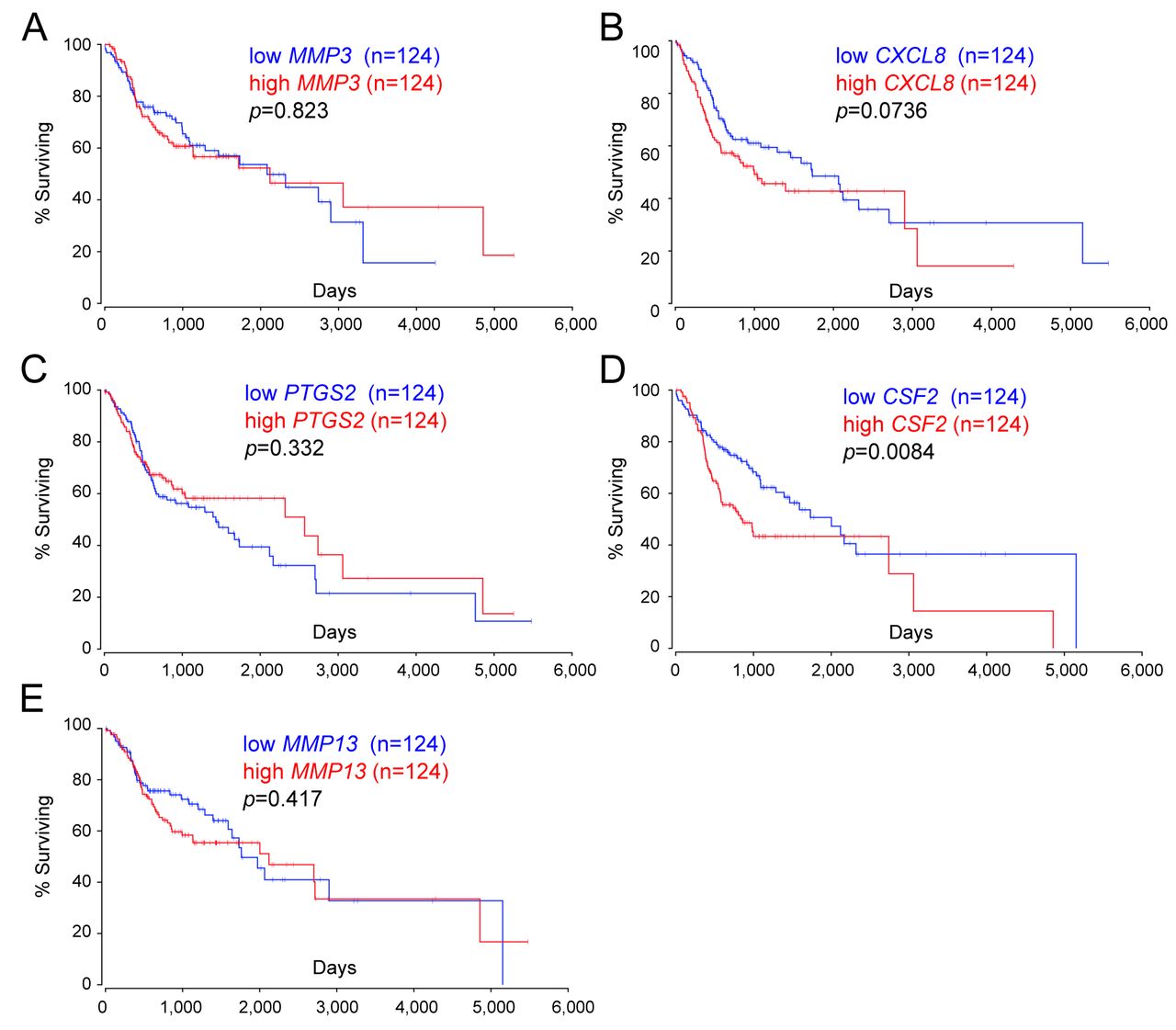
Survival analysis of five mRNAs in head and neck squamous cell carcinoma patients of The Cancer Genome Atlas (TCGA). The overall survival analysis based on the expression of (A) MMP3, (B) CXCL8, (C) PTGS2, (D) CSF2, and (E) MMP13 mRNAs in head and neck squamous cell carcinoma patients including OSCC patients was performed using OncoLnc (http://www.oncolnc.org/).
Discussion
This is the first study analyzing the mRNA transcriptome between HSC-3 and HSC-3-M3 cells. Although various researchers have carried out transcriptomic analyses of OSCC, information on the mRNA profile between parental and its subline OSCC cells is limited. Given that HSC-3 and HSC-3-M3 cells were derived from the same individual as shown in our STR and sex-typing analyses (Table I), the distinct gene expression pattern in HSC-3-M3 cells was considered to be epigenetically established during the metastatic process. Although it remains unclear how the HSC-3-M3 cells were selected during the in vivo screening from parental HSC-3 cells (15), signaling cascades activated by microenvironmental stimuli may affect gene expressions related to metastasis.
Aberrant inflammation is a typical example of microenvironmental stimuli that are involved in various diseases. Mounting evidence indicates that the inflammatory signal is linked to cancer initiation and progression in various cancer cell types (29). It is reported that TNF-α, a key regulator of the inflammatory response, promotes metastasis of OSCC cells via NF-kB (30). Our analyses suggested that TNF signaling and NF-kB are predicted to be activated in HSC-3-M3 cells compared to HSC-3 cells (Figures 1 and 2). Almeida et al. reported that NF-kB is constitutively activated in advanced head and neck cancer cells including OSCC cells and this activation is involved in the acquisition of anti-cancer drug resistance (31). Interestingly, our KEGG pathway analysis showed that up-regulated DEGs in HSC-3-M3 cells were associated with the “Platinum drug resistance” pathway (Figure 1E). Cancer progression phenotypes such as metastasis and anti-cancer drug resistance should be analyzed in considering inflammation and NF-kB activation.
NF-kB is a protein complex consisting of RelA (p65), RelB, c-Rel, p105/p50 (NF-kB1), and p100/p52 (NF-kB2) (29). These proteins regulate the transcription of downstream genes. As shown in Figure 3, IPA predicts that NF-kB might control MMP3, CXCL8, PTGS2, CSF2, and MMP13. Several reports have shown that the transcription of these genes is directly regulated by NF-kB (29, 32, 33). The expression of these mRNAs, except CSF2, was not significantly associated with patient prognosis (Figure 4). However, each factor is reported to be involved in the metastasis of OSCC (17, 34-36). Therefore, the cooperative regulation of OSCC progression by these factors together with upstream regulators remains to be investigated.
The inflammatory cytokine, CSF2 (also known as GM-CSF), is involved in the differentiation of various cells such as dendritic cells (37). It plays a crucial role not only in the physiological process but in the development and progression of a variety of malignancies. It has been reported that CSF2 mRNA is expressed in some cancer cells, and serves as a poor indicator of prognosis. Recurrent and metastatic head and neck cancer patients including OSCC were shown to secrete high levels of CSF2 (38). Consistent with this report, our survival analysis showed that a high expression of CSF2 mRNA leads to poor prognoses of head and neck cancer patients (Figure 4D). Importantly, the transcription of CSF2 gene is directly regulated by NF-kB, AP-1, and ETS1 (39). Reports have also indicated that chemo-resistant pancreatic cancer cells overexpress CSF2 via activation of the NF-kB pathway (40). This NF-kB-mediated CSF2 secretion enhances the differentiation of monocytes into myeloid-derived suppressor cells, leading to T-cell suppression and pancreatic cancer progression (40). Moreover, secreted CSF2 is thought to enhance the growth of urothelial carcinoma using autocrine and paracrine mechanisms (41). On the other hand, the tumor-suppressive effects of CSF2 on tumor progression should be considered. For example, high levels of CSF2 are associated with an improved 5-year survival rate in colorectal cancer (42). We should carefully analyze the diverse roles of CSF2 on tumor progression in various cancer cell types and disease stages (43).
Considering that NF-kB pathway activity is correlated with tumor metastasis in patients with OSCC, its signaling effectors may act as biomarkers to detect and treat metastasis (44). Regarding the roles of CSF2 on OSCC progression, Wagai et al. showed that CSF2 positively regulates OSCC proliferation (45). Notably, secreted CSF2 is also known to stimulate NF-kB signaling (37); therefore, it is assumed that CSF2 and NF-kB pathways may crosstalk and maintain the gene expression pattern in HSC-3-M3 cells through a positive feedback mechanism. Figure 3 also shows that CSF2 and NF-kB pathways are indirectly connected via several other pathways, such as P38 mitogen-activated protein kinase and epidermal growth factor pathways that are well known for inducing OSCC progression (46, 47). Based on these findings, further detail analyses are needed to demonstrate how the CSF2 or NF-kB pathways regulate each other and the metastatic phenotype of HSC-3-M3 cells. As CSF2 is also produced by hemopoietic cells such as lymphocytes, we must to consider the effects of CSF2 secreted by inflammatory cells in microenvironment. Breaking the regulatory cycle between cancer cells and inflammatory cells has been suggested as a promising strategy against metastasis (48, 49).
In the present study, we focused on the analysis of HSC-3-M3 cells as a highly metastatic subline, but other OSCC sublines should also be considered (16-21). Muramatsu et al. established a highly metastatic OSCC cell subline, HOC313-LM, from parental HOC313 cells using in vivo screening (21). Similar to HSC-3-M3 cells, HOC313-LM cells have a higher metastatic potential compared to parental HOC313 cells. The same study also led to the identification of a hypusine cascade and RhoA signaling in HOC313-LM cells (21). It remains to be investigated whether these cascades and pathways are involved in the activation of inflammation and NF-kB in both HSC-3-M3 and HOC313-LM cells. We hypothesize that integrative transcriptomic analyses among these different OSCC sublines would help identify novel pathways and molecules associated with cancer progression. It has been reported that non-coding RNAs like microRNA (miRNA) and lncRNA regulate gene expression in cancer cells (14, 50-52). Recent report showed that the expression of some miRNAs are differentially expressed in HSC-3 and HSC-3-M3 cells (53). Thus, it would be of interest for future studies to assess the entire transcriptome including non-coding RNAs in these cell lines.
We analyzed HSC-3-M3 cells, whose molecular characteristics have not yet been determined. Our results also indicated that they are a useful cell line model for studying OSCC progression. The limitation of this study is that the predicted pathways and networks were not sufficiently validated. Various other experimental approaches using animal and cell line models are needed to verify our results. These results should also be confirmed by clinical studies on patients with OSCC. We plan to conduct further in vitro and in vivo studies to analyze and validate how the pathways and networks affect downstream gene expression and the metastatic phenotype.
In conclusion, transcriptomic analysis using HSC-3 and HSC-3-M3 cells showed the potential involvement of inflammatory and NF-kB pathways in OSCC metastasis. And CSF2 might be a diagnostic as well as therapeutic molecule. Although more detailed biochemical analyses to validate the predicted pathways, networks, and molecules are needed, the results presented herein provide certain insights into the molecular mechanisms of OSCC metastasis.
Acknowledgements
The Authors are grateful to Advanced Medical Research Center, Yokohama City University for technical support of IPA. The authors thank members of the Department of Nutrition and Dietetics, Kamaukura Women’s University for helpful cooperation and discussion. And the authors also thank Enago (https://www.enago.jp/) for the English language review. This research was funded by Grants-in-Aid for Scientific Research from Japan Society for the Promotion of Science (grant number 18K09545 and 20K18705).
Footnotes
Authors’ Contributions
K.S. conceived and supervised the study. Y.I. and K.S. performed the experiment, and interpreted all results. T.T. contributed to analyze and interpret the results of microarray and IPA. Y.H., J.B., K.T., and K.M. participated in planning the study. All Authors involved in writing the paper, and approved the final manuscript.
This article is freely accessible online.
Conflicts of Interest
The Authors declare that they have no competing interests.
- Received October 29, 2020.
- Revision received November 25, 2020.
- Accepted December 4, 2020.
- Copyright© 2021, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}