


Abstract
Background: Salivary gland cancer is a common type of head and neck cancer characterized by occasional deep invasion and lung metastasis. The precise role of sex steroid hormones in salivary gland cancer is unclear. To address this issue, we investigated whether the estrogen axis modulates salivary adenocarcinoma (SAC) and whether hormone therapy can be an effective treatment. Materials and Methods: The estrogen receptor (ER) was overexpressed in HSG human SAC cells that lack endogenous ER and the cells were treated with and without 17β-estradiol (E2). Results: E2 enhanced malignant phenotypes. Moreover, E2 treatment reduced E-cadherin expression, while increasing that of N-cadherin, vimentin, and inhibitor of differentiation 1 proteins that are associated with the epithelial–mesenchymal transition. Cell invasion was enhanced through activation of matrix metalloproteinase-9. Conclusion: These results indicate that hormone therapy used in breast cancer may also be effective for ER-positive SAC.
- 17β-Estradiol
- epithelial–mesenchymal transition
- AKT
- estrogen receptor
- hormone therapy
- MMP9
- salivary adenocarcinoma
Estrogen receptor (ER), a sex steroid hormone receptor, is intimately associated with cancer of the female reproductive organs (1, 2). ERs are steroid receptors located in the cytoplasm and on the nuclear membrane. Human, mouse, rat, dog, and cat ERα present more than 90% similarity, but ERα structure may differ among species. Estrogen can be an agonist and antagonist depending on its target organ (3). Estrogen binds to ERs, inducing conformational change and downstream cascades. While ERs are known to be expressed in tissues of the female reproductive tract and breast, recent studies indicate that ERs are also expressed in other tissues such as the bone, brain, liver, colon, skin, and salivary gland (4). Therefore, ER was investigated as a potential target in other organs such as the prostate, colon, lung, and stomach (5). ER influences many biological processes, not only in women, but also in men.
Head and neck malignancies, especially those of the oral cavity, account for about 3% of all cancer based on long-term studies (6). Salivary gland cancer (SGC) is the most dominant type of cancer after squamous cell carcinoma (6). SGC is known to morphologically mimic breast tumors with respect to histology and steroid hormone receptor status (7). Salivary adenocarcinoma (SAC) is among the common malignant SGC types, with an aggressive nature. Unfortunately, only few cell lines have been established from salivary cancer. We possess four cell lines of salivary cancer origin, but none of them expresses ERs. However, it was reported that ERα is expressed at higher levels in high-grade malignant SGCs than in low-grade malignant and benign SGCs (8).
Therefore, in this study, we transfected a SAC cell line with ERα and used this cell line to determine whether the estrogen–ER axis is functional in human SAC cells.
Materials and Methods
Cell culture. Human HSG cells were established from an adenocarcinoma of the submandibular gland (9) and were a generous gift from Professor Sato (Tokushima University, Tokushima, Japan). The human breast cell line, T47D, was obtained from the American Tissue Culture Collection (Manassas, VA, USA) and used as a control like. Cells were cultured in RPMI-1640 (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 5% charcoal-stripped fetal bovine serum (FBS) at 37°C in the presence of 5% CO2.
Transfection of HSG cells with ER. The pSG5-ER plasmid encoding human ERα was a gift from Professor P. Chambon (Institute of Genetics and Molecular and Cellular Biology, Strasbourg, France). The pBK-CMV vector (Stratagene, La Jolla, CA, USA) containing the neomycin resistance gene was co-transfected with pSG5-ER into HSG cells using Lipofectamine Plus reagent (Life Technologies, Carlsbad, CA, USA). Neomycin-resistant clones were selected with medium containing G418. Cells co-transfected with empty pSG5 and pBK-CMV vectors served as controls. HSG cells expressing ERα were pooled and after assessing ERα expression levels, clones with different expression levels of ERα were produced.
Chemical treatment. 17β-Estradiol (E2) was obtained from Sigma-Aldrich and prepared as a 10-mM stock solution in ethanol. For most experiments, E2 was used at a concentration of 10 nM in 10 ml culture medium for a final ethanol concentration of 0.1%. Control cells were treated with 0.1% ethanol only. Cells were treated with ethanol or E2 (10 nM) once daily for 2 days.
Western blot analysis. Cells were lysed in 2× Laemmli buffer and stored at −70°C. Protein concentration was determined using the DC Protein Assay kit (Bio-Rad, Hercules, CA, USA). Samples (20-30 μg of total protein) were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Hybond P; Amersham Biosciences, Little Chalfont, UK). Membranes were blocked for 1 h at room temperature with Tris-buffered saline with Tween-20 (20 mM Tris, 137 mM NaCl, 3.8 mM HCl, and 0.1% Tween-20) containing 5% nonfat milk, and blots were probed with anti-ERα (EP1; DAKO, Glostrup, Denmark), anti-protein kinase B (AKT, 610860; BD Biosciences Pharmingen, San Diego, CA, USA), anti-pAKT (Thr308) (558275; BD Biosciences Pharmingen), anti-pAKT (Ser473) (560404; BD Biosciences Pharmingen), anti-E-cadherin (HECD-1; Abcam, Cambridge, UK), anti-N-cadherin (32; BD Biosciences, San Jose, CA, USA), anti-vimentin (V9; DAKO), anti-inhibitor of differentiation 1 (ID1) (Z-8; Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-matrix metalloprotease 9 (MMP9) (ab35326; Abcam plc), or anti-actin (C4; Chemicon International, Temecula, CA, USA) for 1 h. The membranes were then washed and incubated with a secondary antibody (either goat anti-rabbit or anti-mouse IgG-horseradish peroxidase) (Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight, washed again, and developed using enhanced chemiluminescence with the Amersham ECL-Plus kit according to the manufacturer's instructions.
Cell proliferation assay. After incubation of HSG cells for 24 h with E2 (10 nM) or ethanol, cell proliferative ability was determined by cell counting. Cells were digested with trypsin-ethylenedia-minetetra-acetic acid at 37°C for 5 min. Next, 8 ml of conditioned medium containing 5% FBS was added to the cells and the cells were collected. Then, 0.5 ml of the supernatant was diluted with 9.5 ml of balanced electrolyte solution. The cell number was determined by particle counting and analyzed using a Multisizer™ 3 system (Beckman Coulter, Brea, CA, USA). Assays were performed in triplicate and the results were averaged.
Boyden chamber invasion assay. Invasion assays were performed in modified Boyden chambers with 8-μm-pore filter inserts for 24-well plates (Collaborative Research, Waltham, MA, USA). The filters were coated with 10-12 μl of ice-cold Matrigel (Collaborative Research). Cells (40,000 cells/well) were added to the upper chamber in 200 μl of serum-free medium. The lower chamber was filled with 300 μl of conditioned medium from fibroblasts. After incubation for 20 h with E2 (10 nM) or ethanol, the HSG-ER cells were fixed with 2.5% glutaraldehyde and stained with 0.5% toluidine blue in 2% Na2CO3. Cells that remained in the Matrigel or attached to the upper side of the filter were removed with cotton tips. Cells on the lower side of the filter were counted under a light microscope. Assays were performed in triplicate or quadruplicate and the results were averaged.
Zymography. Proliferating HSG cells (1×106 cells in 100-mm dishes) were transferred to serum-free medium for 2-3 days, after which the medium was replaced with 10 ml of fresh serum-free medium. Forty-eight hours later, the conditioned medium was collected and concentrated 10- to 15-fold using 10 kDa cutoff filters (EMD Millipore, Darmstadt, Germany). The concentrated medium was analyzed using gelatin substrate gels. Briefly, gels consisted of 8-10% (v/v) polyacrylamide and 3 mg/ml gelatin (Sigma-Aldrich). Concentrated conditioned medium was mixed with non-reducing Laemmli sample buffer and incubated at 37°C for 15 min. After electrophoresis, the gels were incubated for 1 h in 2.5% (v/v) Triton™ X-100 at room temperature, followed by 24-48 h in substrate buffer [100 mM Tris-HCl (pH 7.4) and 15 mM CaCl2]. The gels were stained with Coomassie Blue for 30 min and destained with 30% methanol/10% acetic acid (v/v). E2 (10 nM) or ethanol was also used in this experiment.
Statistical analysis. Statistical comparisons were performed using the Wilcoxon signed-rank test. p-Values of less than 0.05 were considered statistically significant. All statistical tests were performed using Statcel2 software (Statcel2, OMS, Tokyo, Japan).
Results
ER overexpression in HSG cells. HSG cells were transfected with a vector encoding ERα. Twenty four transfectants were established and screened by western blotting. T47D cells expressing human ER were used as positive controls in this experiment. We selected the HSG transfectant that expressed ER to a level similar to that of T47D cells for the subsequent experiments (Figure 1). HSG-ctl transfected with two empty vectors did not express ER.
Effect of E2 on cell proliferation and expression of AKT of HSG cells. Treatment with E2 (10 nM) for 24 h significantly enhanced the proliferation of HSG-ER cells when compared to that of ethanol-treated cells (p<0.01) (Figure 2A).
Western blotting results indicated that E2 treatment induced the phosphorylation of AKT on residues Thr308 and Ser473 in HSG-ER cells (Figure 2B). No effect on proliferation and AKT phosphorylation in HSG-ctl cells was observed after E2 treatment (data not shown).
Changes in the expression of epithelial–mesenchymal transition (EMT) markers and cell morphology in E2 or ethanol treated HSG-ER cells. We next investigated whether E2 modulated the expression of genes known to be up-regulated (N-cadherin, vimentin, and ID1) or down-regulated (E-cadherin) during EMT. E-Cadherin expression decreased whereas that of N-cadherin, vimentin and ID1 increased in E2-treated HSG-ER cells when compared with ethanol-treated cells (Figure 3A).
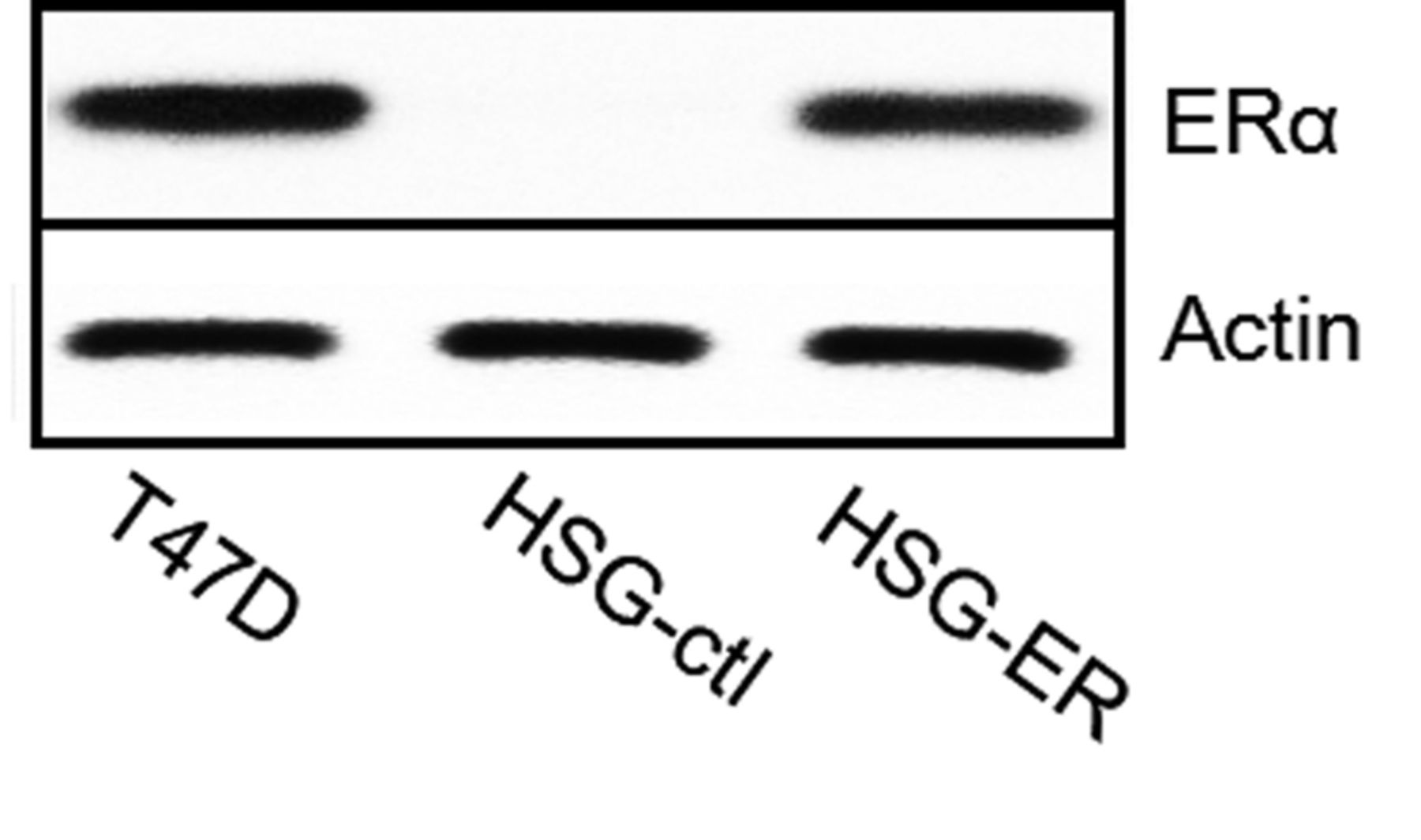
Estrogen receptor (ER) overexpression in ER-deficient HSG cells. Actin served as a loading control. These HSG-ER cell populations were used for further experiments. HSG-ctl cells were transfected with an empty vector.
Moreover, the cell morphology drastically differed between E2- and ethanol-treated HSG-ER cells (Figure 3B). Ethanol-treated cells had a more flattened appearance and formed monolayers, while E2-treated cells grew in a disorderly fashion and a large fraction of the cells formed multilayers.
There was no effect of E2 treatment on gene expression or cell morphology in HSG-ctl cells (data not shown).
Effect of E2 treatment on cell invasion and MMP9 activity. Next, we compared the invasive phenotype of HSG-ER cells upon treatment with E2 and ethanol using the Boyden chamber invasion assay. Invasiveness was significantly enhanced in E2-treated HSG-ER cells when compared to that of ethanol-treated cells (p<0.01) (Figure 4A).
Accordingly, the expression of MMP9, one of the major gelatinases secreted by aggressive cancer cells, was also induced in E2-treated HSG-ER cells when compared with ethanol-treated cells (Figure 4B).
Discussion
The prognosis of aggressive SGC is poor and the only reliable treatment is surgery because SGCs a relatively rare and heterogeneous group of tumors with variable pathological and phenotypic characteristics (10). New treatment modalities are urgently needed.
A previous report indicated that ERβ inhibits the migration and invasion of breast cancer cells and up-regulated E-cadherin expression in an ID1-dependent manner (11). Sometimes, ERβ, in contrast to ERα, does not act as an oncogene. High ERβ expression is associated with a significantly decreased risk of breast cancer in women with atypical hyperplasia (12). Moreover, while ERα is a prognostic marker of breast cancer, ERβ is not. (13). ERα is an oncogenic transcription factor and treatment with 4-hydroxytamoxifen is well established (14, 15).
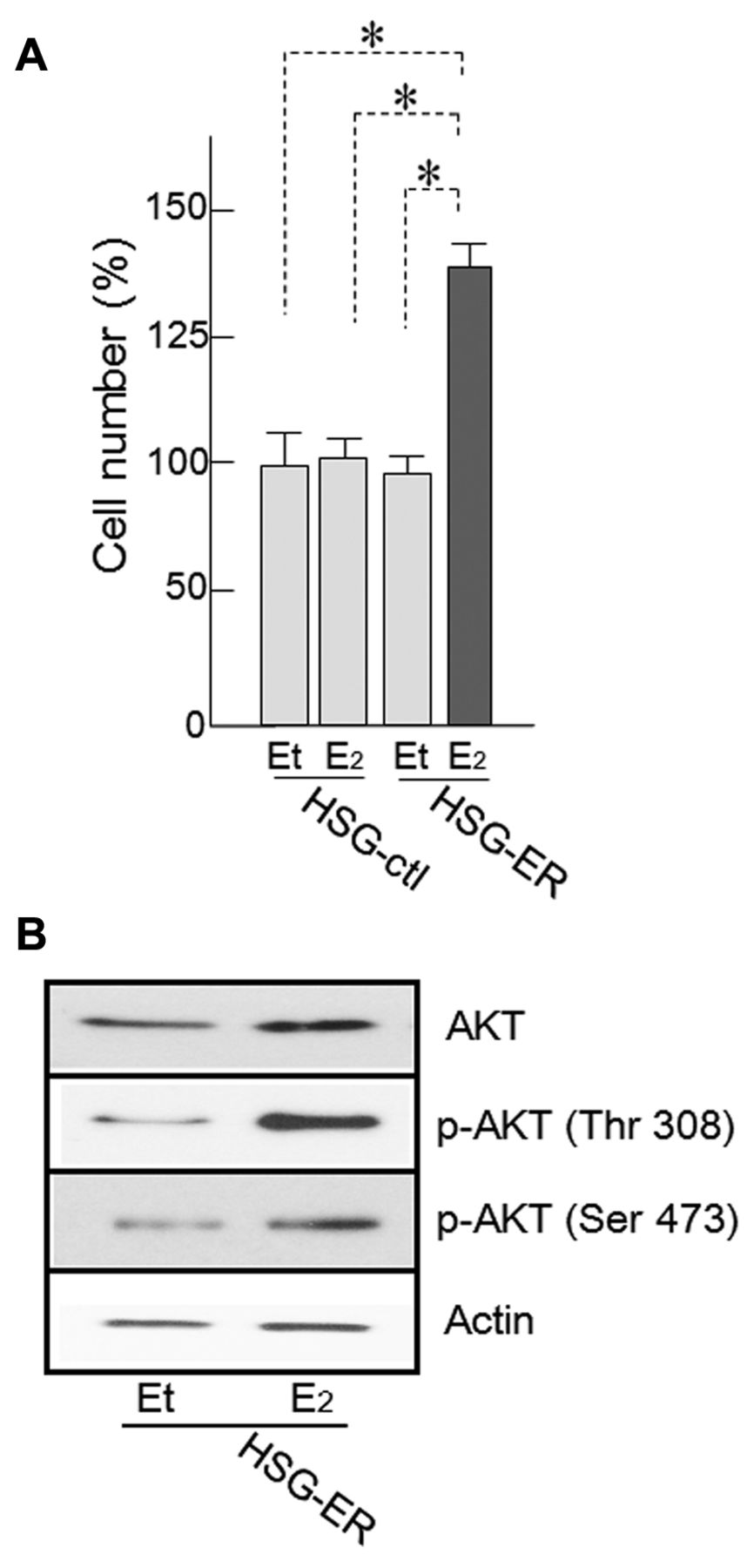
Effect of 17β-estradiol (E2) treatment on HSG-estrogen receptor (ER) cell proliferation. A: HSG cell proliferation was enhanced by E2 treatment. Treatment with 10 nM E2 for 24 h significantly enhanced the proliferation of HSG-ER cells and HSG-ctl cells (*p≤0.01). B: Protein kinase B (AKT) phosphorylation was also up-regulated. Western blotting indicated that the quantity of phosphorylated (p)-AKT (Thr308, Ser473) increased. Et: Ethanol (vehicle).
The proliferation of HSG cells overexpressing ERα was up-regulated by E2 treatment, indicating, as far as we are aware for the first time, that the E2–ERα axis is functional in this experimental system. This increase in proliferation was accompanied by AKT phosphorylation. Similar results were obtained in different breast cancer cells expressing ERα (16), and the up-regulation of cyclin D1 was also mentioned, explaining the effect of E2 on cell proliferation. ERα-dependent AKT activation was previously reported (17, 18, 19). Thus, this pathway might be the main signaling pathway underlying the proliferation of SGC cells. Moreover, E2 treatment affected cell morphology. Therefore, we also investigated the expression of genes associated with EMT. The results indicated that the expression of E-cadherin was down-regulated, while that of N-cadherin, vimentin, and ID1 was up-regulated by E2 stimulation, consistent with the EMT process.

A: Expression profiles of proteins associated with the epithelial–mesenchymal transition are presented. B: The change in HSG-estrogen receptor (ER) cell morphology after E2 administration was also observed. Et: Ethanol (vehicle).
In addition, E2 treatment significantly increased the invasive capacity of HSG-ER cells and up-regulated MMP9 expression. AKT is known to be associated with MMP9 up-regulation (20, 21). However, up-regulation of ID1 cannot be ignored in this study. ID1 can enhance tumor cell migration, which is associated with the secretion of MMP9 (22, 23). Moreover, ID1 is also associated with the EMT (24, 25).
A recent study reported that 66.7% of high-grade SGCs were positive for ERα, while only 20% of the low-grade lesions were ERα-positive (8). Another report mentioned that ERα-dependent protein kinase, CK2α, regulates AKT expression and activation in oncogenesis (26). Taken together, these findings indicate that the E2–ERα axis might be a target for the treatment of SGC.
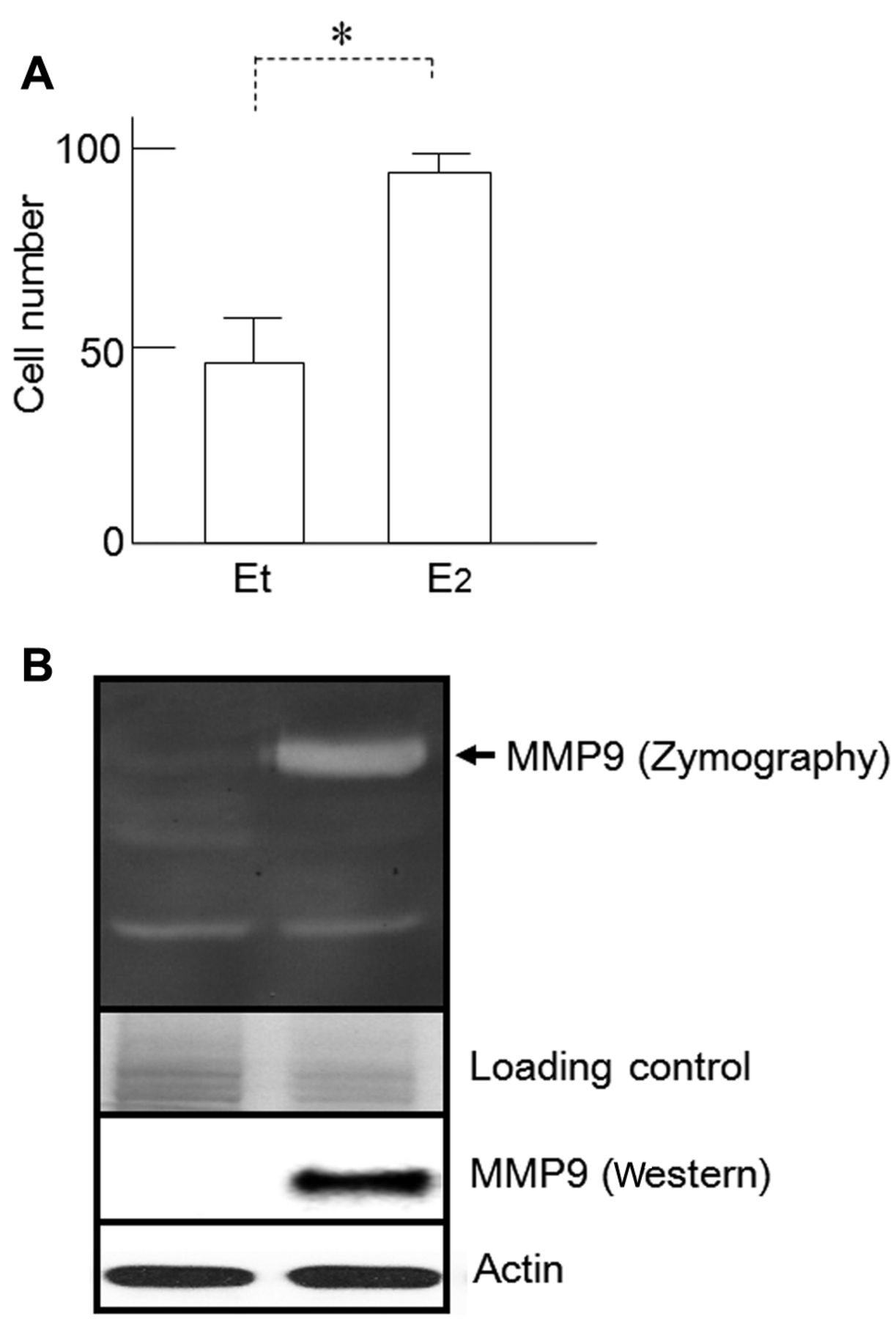
Effects of E2 on HSG cell invasiveness and matrix metalloproteinase 9 (MMP9) secretion. A: Cell counts of HSG-estrogen receptor (ER) and HSG-ctl cell populations as determined by Boyden chamber invasion assays. *p<0.01. B: Gelatin zymography indicated that MMP9 secretion by HSG-ER cells was increased by E2 administration. Western blot analysis indicated that MMP9 protein was strongly up-regulated in HSG-ER cells after E2 treatment. Et: Ethanol (vehicle).
In conclusion, our results clearly show that the E2–ERα axis is functional in HSG-ER cells, and that E2 treatment results in an increase in malignant phenotypes of these SGC cells such as increased proliferation and invasion. ER has recently garnered attention as a potential therapeutic target in malignancies arising from organs other than the female reproductive tract, such as gallbladder (27), prostate (28), lung (29), and hepatocellular (30) carcinomas. Future studies will be designed to determine whether 4-hydroxytamoxifen is an effective treatment approach for ER-positive SGCs.
Acknowledgements
This work was supported by JSPS KAKENHI Grant Number 15K11257. The Authors gratefully acknowledge this financial support. They would also like to thank Editage (www.editage.jp) for English language editing.
Footnotes
Conflicts of Interest
None of the Authors has a financial conflict of interest to disclose in relation to the content of this article.
- Received April 9, 2016.
- Revision received May 12, 2016.
- Accepted May 17, 2016.
- Copyright© 2016, International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.