


Abstract
Background: miRNAs play important roles in multiple biological processes, and deregulation has been linked to several human diseases, including cancer. Studying changes in miRNA expression in cancer development is commonly performed in vitro in human cancer cell lines using quantitative polymerase chain reaction (qPCR), a method requiring the use of a robust reference gene that displays stable expression across all samples. Materials and Methods: Using the NormFinder software, a selection of commonly used endogeneous controls and miRNAs were tested in six human cancer cell lines to identify for the most suitable gene for use as a reference. Results: The frequently used endogenous control U6B small nuclear RNA (RNU6B) was found to be an unsuitable reference for normalization. The most suitable single endogeneous control identified was miR-25-3p, whereas the best combination of two endogeneous controls was miR-25-3p and miR-93-5p. Conclusion: We identified a single and a pair of miRNAs suitable for use as endogenous controls when performing qPCR-based miRNA expression analyses in human cancer cell lines.
MicroRNAs (miRNAs) are single-stranded RNAs which have roles in a wide range of fundamental cellular processes, including embryonal development, proliferation, apoptosis, and stress response (1, 2). These RNA molecules are produced from larger transcripts that are processed to form hairpin precursors that serve as substrates for Drosha and Dicer, members of the RNase III enzyme family, to generate mature 20-22 nucleotide miRNAs (3). These small non-coding RNA molecules lead to down-regulation of gene expression through translational inhibition or mRNA cleavage by binding to complementary sites in target transcripts (4). Precise control of miRNA expression is crucial for keeping cells in normal physiological states, and dysregulation of miRNAs may lead to disease, including cancer (5, 6). It has been shown that over 60% of human protein-coding genes are conserved targets of miRNAs (7) and most miRNAs are highly conserved across vertebrates (1, 8). miRBase, an online database of miRNA sequences, currently contains, precursors and, mature miRNAs of the human genome (9).
In recent years, several studies have shown that altered expression of specific miRNA genes contributes to the initiation and progression of cancer (10-13). The mechanism through which miRNAs contribute to cancer development and progression is often unknown but specific miRNAs have been shown to target genes that are critical regulators for the development of cancer, such as the E2 family (E2F) and RAS (14, 15). There is therefore an impetus for studying the expression patterns of individual miRNAs in cancer tissues, both to elucidate the molecular mechanisms involved in cancer development and progression, and to identify diagnostic and prognostic biomarkers. Due to their high availability, cancer cell lines are often used as experimental models of neoplastic disease in these studies. miRNA gene-expression analyses using cell lines can thus provide candidate molecular signatures characteristic of the tumor from which the cell line is derived.
One of the most commonly used techniques to study miRNA expression is quantitative polymerase chain reaction (qPCR). Due to the small size of miRNAs, the method is usually based on the addition of a universal tag sequence on the 5’ end during reverse transcription. This allows amplification of target miRNAs using a specific forward primer and a universal reverse primer. To correct for systematic variables, such as differences in quantity of starting template, RNA quality and enzymatic efficiency, qPCR data are normalized against those of an endogenous control gene. It has been reported that it is preferable to normalize target RNA levels with control genes belonging to the same RNA class (16), rendering the commonly used qPCR endogenous controls beta-actin (ACTB), glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and 18s unsuitable in quantitative miRNA assays. Rather, for normalization of miRNA levels, the endogenous control genes should belong to the small non-coding RNA family (ncRNA) of RNAs, such as small nuclear RNAs (snRNA). Ideally, the endogenous control should show invariable expression across the test sample set and exhibit storage stability, extraction efficiency and quantification efficiency equivalent to that of the target nucleic acids. Additionally, miRNAs represent as little as 0.01% of the total mass of RNA, but with concentrations greatly dependent on sample type (17). Thus, the endogenous control used should mirror this quantitative change in the target miRNA.
Cell line overview.
Primer overview.
There is no universal endogenous control suitable for every tissue type, as the expression of most RNAs varies with cell type and condition. A suitable endogenous control should, therefore, be validated for each sample set. The most commonly used endogenous control for miRNA expression studies is U6B small nuclear RNA (RNU6B), a snRNA molecule. A number of reports indicate that snRNAs are expressed both abundantly and stably, making RNU6B a good candidate as a miRNA endogenous control (17, 18). It has, however, been shown to be unsuitable for this purpose for many sample sets due to its low stability across tissues (19, 20). Instead, stably expressed miRNAs have been suggested as endogenous controls (21, 22).
In order to select suitable reference genes for normalization, many analysis tools, such as Normfinder, geNorm and Bestkeeper can be used (23-25). In the present study, we used Normfinder, as well as evaluation of the raw Ct values to assess 12 candidate reference genes in six human cancer cell lines. Small RNA extraction was used to efficiently recover all classes of small RNA populations. Our aim was to identify suitable endogenous controls for qPCR analysis of miRNA gene expression in selected human cancer cell lines.
Materials and Methods
Cell lines and RNA extraction. Six human cancer cell lines were used in this study (Table I). The cells were cultured under normal conditions, trypsinized and harvested for RNA isolation. Small RNA was isolated using the mirVana miRNA isolation kit (Life Technologies, Carlsbad, CA, USA) from all cell lines according to the manufacturer's protocol. RNA concentrations were determined by measuring the A260/280 absorbance on a Nanodrop 1000 (Thermo Fisher Scientific, Wilmington, DE, USA).

Raw Ct values for 12 candidate endogenous controls tested in six human cancer cell lines. The error bars represent the standard deviation (SD) of the eight replicates used for each sample.
Quantitative PCR. cDNA synthesis was performed on 6 ng small RNA using the miScript II RT kit (Qiagen, Valencia, CA, USA). The cDNA samples were amplified using the miScript SYBR® Green PCR kit (Qiagen), the provided miScript Universal Primer (reverse primer) and a small RNA specific primer (forward primer). Primers targeting candidate control genes were ordered from the miScript Primer Assay catalogue (Qiagen). Candidate endogenous controls were selected from the literature (21, 31, 32) and from suggested miRNA endogenous controls in the miScript Primer Assay catalogue (Qiagen). An overview of the primers used is given in Table II. The PCR reactions were performed in a final volume of 20 μl, and a total of 0.1 ng cDNA was used per reaction. The reactions were performed on a Stratagene Mx3000p instrument using the PCR program recommended in the miScript SYBR® Green PCR kit protocol.
All reactions were carried-out using eight replicates. Ct values >35 were regarded as negative and the melting point curves were manually inspected for all assays to verify primer specificity (data not shown).
Data analysis. To obtain a crude measure of the stability for the candidate reference genes, the raw Ct values for each gene were evaluated across all cell lines. The standard deviation between the eight replicates and the PCR amplification efficiencies were calculated for all assays. PCR amplification efficiencies were calculated for each assay using the formula, E=(10−1/slope−1)×100. Assays with efficiency >90% or <110% were included for further analysis. Furthermore, stability of the endogenous control gene expression was analyzed using the Normfinder analysis tool (23) (http://moma.dk/normfinder-software). Firstly, all the candidate endogenous controls (n=12) were tested in all cell lines (n=6). All raw Ct values from measurements by qPCR were transformed to linear scale before used as input in NormFinder.
Results
Expression levels of candidate reference genes. To evaluate the potential endogenous normalizers of miRNA genes in human cancer cell lines, a selection of candidate reference genes was tested for stable expression across six cell lines (Table I). To limit the chance of large variability in storage stability, extraction efficiency and quantification efficiency between the reference and target gene, the candidate reference genes were limited to small non-coding RNAs only. The potential reference gene set consisted of nine miRNA genes and three small RNA genes (Table II).
As shown in Figure 1, the candidate gene Ct values ranged from 20.75 to 33.53. miR-16 was the most highly expressed gene, with a mean Ct value of 21.86, while miR-455 had the lowest level of expression, with a mean Ct value of 32.41. The Ct value range for one candidate reference gene among the cell lines is a crude measure of the stability for this gene. The mean Ct value range for miR-455, miR-17, miR-93, miR-103, miR-345, RNU6B and small nucleolar RNA D68 (SNORD68) was >3 cycles, whereas only miR-25 had a mean Ct value range <2. Of particular interest is the large variation in Ct values across the cell lines for RNU6B, with a mean Ct value range of 3.44 cycles. Due to amplification efficiency outside the wanted range, the RNU6B and miR-140-3p assays were omitted from further analyses (Table II), whereas the remaining 10 candidate reference genes were analyzed for expression stability across the different cell lines. The standard deviation for inter-assay replicates ranged from 0.077-0.744.
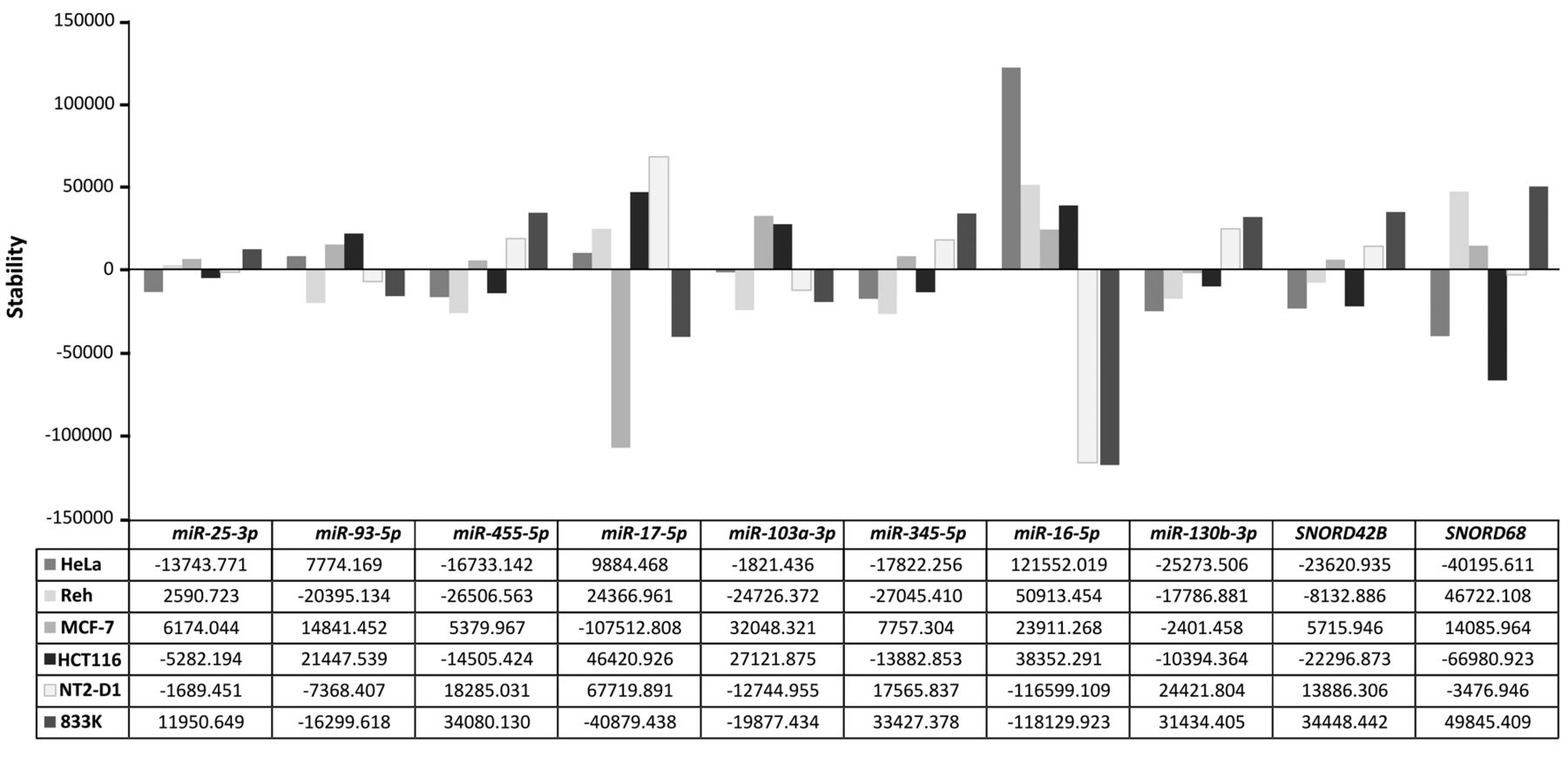
Variation across six human cancer cell lines in 10 candidate endogenous controls. The stability of each candidate endogenous control as measured by NormFinder is displayed in the bar charts in the upper panel. The corresponding inter-cell line stability values are given in the lower panel.
Expression stability of candidate reference genes. NormFinder was used to assess the stability of the 10 candidate references. This method ranks the candidate genes based on intra- and inter-group variations through an algorithm calculating a ‘stability value’ that is inversely correlated with the stability of gene expression (i.e. a high stability value indicates low stability). NormFinder ranked the 10 candidate references from lowest to highest stability value (Figure 2). This ranking of each gene by the stability showed that the best single gene was miR-25-3p, while the best two-gene combination was that of miR-25 and miR-93 (Table III).
Discussion
miRNAs are recognized as important factors in gene regulation of physiological and pathological processes. To date, 35,828 mature miRNAs from 223 species are registered in the miRBase (Release 21, June 2014) (http://www.mirbase.org) (9). As miRNAs regulate gene expression through a ‘fine-tuning’ mechanism, studying the role of individual miRNAs in a specific physiological or pathological experimental situation depends on a reliable and accurate technique for measuring expression levels. qPCR-based relative quantification of gene expression is considered the gold standard for gene expression analysis in tissues and cell cultures. This method, however, strongly relies on the selection of one or more stably expressed reference genes. Indeed, normalization to an unsuitable reference gene has resulted in biased results (33). Since no universal endogenous control exhibiting stable expression in all analytical settings exists, a suitable reference gene should be verified for each analytical system. In this study, we experimentally evaluated 10 candidate reference genes for use in normalization in qPCR measurements of miRNA expression across six different cancer cell lines.
Evaluation of 10 candidate reference genes based on average stability values across six cancer cell lines as calculated by NormFinder.
Although most miRNA expression studies used miRNAs or other small RNAs as reference genes, several studies also normalized the miRNA expression against mRNAs such as GAPDH. This raises the question whether mRNAs can be used as reference genes in miRNA expression studies. Recently, mRNAs have been found to be unsuitable as reference genes in miRNA studies, as different groups of RNAs may have different properties affecting storage stability, extraction efficiency and quantification efficiency (34). Additionally, mRNAs are reverse transcribed using a different technique from that for miRNAs. Interestingly, our results indicate that even the much used miRNA endogenous control gene RNU6B is unsuitable as a reference gene under our experimental settings. This is both due to high variability in Ct values across the cell lines tested, and amplification efficiency outside the accepted range. Our results thus support previous results stating that RNU6B should be avoided for normalization purposes in many experimental settings (17, 18).
Prior to performing NormFinder stability analysis, we analyzed the efficiency of each assay using a standard curve. Two assays (RNU6B and miR-140-3p) were found to have too high or low efficiencies and were therefore omitted. The other assays, with efficiency between 90-110%, were included in the NormFinder analysis. Here, the efficiency is assumed to be 100% when calculating the stability values. This generates a discrepancy between estimated and real values, and is a limitation in our study. However, this discrepancy is of limited importance, as an assay efficiency of 90-110% is considered adequate in most studies. Researchers may take the efficiency into account when performing gene-expression analyses using these endogenous controls in the future.
The candidate reference genes regarded as the most suitable for use in the described cancer cell lines, miR-25-3p and miR-93-5p, are both members of the miR-106b-25 cluster (consisting of miR-106b, miR-93, and miR-25). These miRNAs have been found to display changes in expression coinciding with the epithelial-to-mesenchymal transition during carcinogenesis in vitro (35). Similarly, dysregulation in the expression of the miR-106b-25 cluster and the related miR-17-92 cluster has been reported for various tumors, including B-cell lymphoma and solid tumors derived from breast, colon, lung, pancreas, prostate and stomach (10, 36-38). Due to their oncogenic properties, great care must be taken when using these miRNAs as endogenous controls in cancer tissues or cell lines. However, in our setting they performed well and displayed stable expression patterns, indicating that they are suitable as reference genes when performing miRNA expression analyses.
Despite belonging to the same cluster, miR-25-3p and miR-93-5p do not share the same seed sequence, indicating that they have different target sequences. The differences in expression patterns between these two miRNAs in the cell lines tested also indicate that they are not co-regulated, increasing the chance of successfully using them as paired endogenous controls.
In conclusion, we have identified a single and a pair of miRNAs suitable for use as endogenous control when performing qPCR-based miRNA expression analyses in human cancer cell lines. Using NormFinder, miR-25-3p alone or in combination with miR-93-5p were identified as having high expression stability across the cell lines tested.
Acknowledgements
The Authors thank research assistant, Herman Sebastian Folkestad Evensen (Faculty of Health Sciences, Oslo and Akershus University College of Applied Sciences) for his valuable technical assistance. We are also grateful to Professor Heidi Kiil Blomhoff (Faculty of Medicine, University of Oslo) and Dr. Nur Duale (Norwegian Institute of Public Health, Oslo) for kindly providing the cell lines used in this study. This work was supported by Faculty of Health Sciences, Oslo and Akershus University College of Applied Sciences, Norway.
- Received September 25, 2015.
- Revision received October 28, 2015.
- Accepted October 30, 2015.
- Copyright© 2016, International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved




{kind=link}
{kind=link}