


Abstract
Background: Aberrant activation of the canonical Wingless-type MMTV integration site family (WNT)/β-catenin signaling pathway is critical for gliomas. Materials and Methods: In 74 gliomas of different histological grade and in 24 glioblastoma cell lines, protein expression of WNT member 3a (WNT3a), β-catenin and transcription factor 4 (TCF4) was investigated by immunohistochemistry, western blotting, immunofluorescence and immunocytochemistry. In tumors and cell lines, WNT3A expression was assessed at the mRNA level by quantitative real-time polymerase chain reaction. Results: WNT3a was overexpressed at the protein and mRNA levels in malignant astrocytic tumors and cell lines. Cytoplasmic expression of β-catenin was detected in high-grade gliomas and cell lines, with evidence of nuclear translocation on fractionated protein extracts. Activating mutations in the β-catenin encoding gene (CTNNB1) were excluded by direct sequencing. TCF4 was statistically correlated with Ki-67/MIB-1 and cyclin D1 labeling indices. Conclusion: Expression of WNT3a, cytoplasmic β-catenin and TCF4 was significantly associated with the histological malignancy grade and with a worse prognosis for patients with glioma.
The Wingless-type MMTV integration site family (WNT) signaling pathway is involved in different biological processes, such as embryonic development, cell polarity, fate and tissue homeostasis (1). The β-catenin (CTNNB1)-dependent pathway (often referred as “canonical”) includes β-catenin and transcription factor 4 (TCF4) as the key effectors and is reported as an important factor in the development and progression of several human cancer types (2-6).
The signaling transduction cascade is triggered by the binding of WNT ligands to Frizzled (Fz) protein and to low-density lipoprotein receptor-related proteins 5/6 (LRP5/6), followed by cytoplasmic β-catenin accumulation and subsequent nuclear translocation. In the nucleus, β-catenin binds to members of the TCF/lymphoid-enhancer-binding factor (LEF) family of transcription factors and modulates expression of target genes critical for cell proliferation, differentiation, survival and apoptosis, such as cyclin D1, AP-1 transcription factor (FRA1), V-MYC avian myelocytomatosis viral oncogene homolog (c-MYC), V-JUN avian sarcoma virus 17 oncogene homolog (c-JUN) and survivin (7, 8). In contrast, in the absence of WNT ligands, β-catenin is constantly sequestrated within a degradation complex composed of axin, tumor-suppressor adenomatous polyposis coli (APC), casein kinase Iα (CKIα) and glycogen synthase kinase-3β (GSK-3β) proteins. Upon phosphorylation by CKIα and GSK-3β, β-catenin is polyubiquitinated for the proteasomal degradation (Figure 1) (9).
The aberrant activation of the canonical WNT/β-catenin signaling pathway contributes to glioma development and malignant progression (10-15), as well as invasion (16) with prognostic implications (17-19). Among the WNT ligands present in neuronal stem cells, WNT3a increases proliferation and differentiation of neuronal progenitor cells in adult mice brain (20, 21) and it is required for the development of the mammalian hippocampus (22). Previous reports showed overexpression of WNT1 and WNT3a in glioblastoma multiforme and glioma stem cells, demonstrating pathway activation in the malignant transformation and progression of high-grade gliomas (23-25).
Compared to the normal brain, β-catenin and TCF4 are up-regulated in gliomas, while knock-down of β-catenin in human glioma cells by small interfering RNA (siRNA) inhibits cell proliferation and invasive ability, induces apoptotic cell death and delays tumor growth (26, 27). β-Catenin protein levels increase in high-grade astrocytomas and correlate with malignancy and with the gene expression of cyclin D1 and c-MYC (10-12, 18, 28).
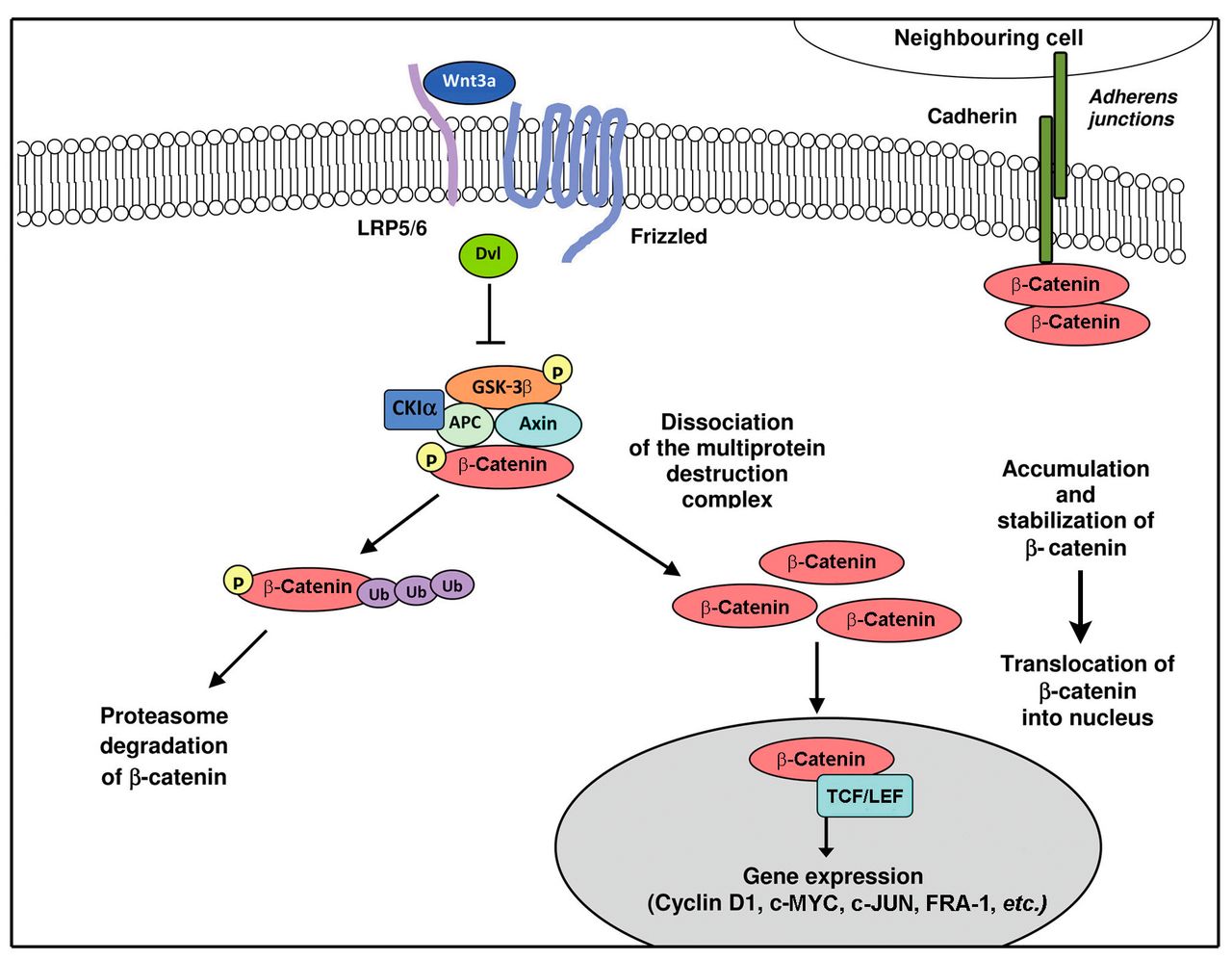
Overview of the canonical Wingless-type MMTV integration site family (WNT)/β-catenin signaling. In the absence of WNT signal, β-catenin is captured by adenomatous polyposis coli (APC) and axin within a destruction complex, phosphorylated (P) by casein kinase Iα (CKIα) and glycogen synthase kinase-3β (GSK-3β) and targeted for ubiquitination (Ub) and proteasomal degradation. Transcription by transcription factor/lymphoid-enhancer-binding factor (TCF/LEF) is off due to the binding of repressors. A pool of β-catenin is engaged in the interactions with cadherins for cell–cell adhesion. The binding of WNT3a ligand to frizzled and low-density lipoprotein receptor-related proteins 5/6 (LRP5/6) co-receptors determines the inactivation of the destruction complex. This allows β-catenin to accumulate in the cytoplasm and translocate to the nucleus, where it forms a transcriptionally active complex with TCF/LEF family members.
In the absence of WNT ligands, the oncogenic activation of β-catenin may result from genetic mutations affecting key components of the canonical pathway [APC, β-catenin-encoding gene (CTNNB1), axin] (29). Activating mutations in the CTNNB1 oncogene contribute to neoplastic transformation in several types of human cancer, such as medulloblastoma, hepatocellular carcinoma, colon and ovarian cancer, with the common halimark of nuclear accumulation of β-catenin (30-33). Unlike other malignancies, however, genetic events in high-grade glioma are rare (14, 31, 34).
Finally, the WNT/β-catenin signaling pathway is involved in the maintenance of tumor stem cells by inhibiting differentiation, conditioning radio- and chemoresistance, and inducing an invasive phenotype in gliomas (35-40).
The aim of the present study was to verify the activation of the canonical pathway by determining the expression of the key players WNT3a, β-catenin and TCF4, in a series of 74 astrocytic and oligodendroglial gliomas and 24 glioblastoma cell lines. We also studied the relationships of the WNT/β-catenin signaling pathway with malignancy and the correlation of TCF4 with cell proliferation and cyclin D1 expression.
Materials and Methods
Brain tumor specimens. A total of 74 adult human brain tumors (31 glioblastomas and 43 grade I-III gliomas) were analyzed. All cases were operated on at the Neurosurgery Unit of CTO Hospital/Health and Science City (Turin, Italy). Surgical tumor specimens, from either total or partial removal, were split into three consecutive fragments. The first one was formalin fixed, paraffin embedded (FFPE) and cut into 5 μm-thick sections. The second one was minced and enzymatically dissociated for expansion in culture, and the third fragment was stabilized in RNAlater® solution (Thermo Fisher Scientific Inc., Waltham, MA, USA) and stored at −80°C for molecular genetics and proteins. Two brain samples from the periventricular region were obtained from patients with epilepsy and diagnosed as normal nervous tissue. The histological diagnosis was performed according to the World Health Organization (WHO) guidelines (41). Matched constitutional DNA from blood/saliva was obtained from each patient. Patient demographics are summarized in Table I.
Patients' demographics.
List of primary antibodies used for immunohistochemistry.
Ethics statement. Human brain specimens for molecular genetics, proteins and cultures were obtained and used in compliance with the local Institutional Review Board and Committee on Human Research and with the ethical human subject principles of the World Medical Association Declaration of Helsinki Research. Written informed consent was obtained from all patients after ethics approval of the CTO Hospital/Health and Science City, Turin, Italy (n. 487/2012).
Immunohistochemistry. In addition to hematoxylin and eosin (H&E) staining, immunohistochemistry was performed with the primary antibodies listed in Table II on a Ventana Full BenchMark® XT automated immunostainer (Ventana Medical Systems Inc., Tucson, AZ, USA). Ultra-View™ Universal DAB and Alkaline Phosphatase Red Detection Kits were used for visualization of antibody reactions (Ventana Medical Systems Inc.). Heat-induced epitope retrieval (HIER) was obtained with Tris–EDTA (pH 8). CD34 immunostaining was used to distinguish isolated endothelial cells from tumor cells.
In vitro cultures. A total of 24 glioblastoma cell lines were established from glioblastoma tumors removed at the Neurosurgery Unit of CTO Hospital/Health and Science City of Turin, Italy. Culture conditions have been previously described (42, 43). Nine cell lines (CV13 NS, CV21 NS, NO3 NS, NO6 NS, CTO3 NS, CTO11 NS, CTO12 NS, CTO15 NS, CTO22 NS) were isolated in Dulbecco's modified Eagle's medium (DMEM)/F-12 supplemented with 20 ng/ml epidermal growth factor (EGF) and 10 ng/ml basic fibroblast growth factor (bFGF) for the neurosphere (NS) assay. Thirteen cell lines (CV2 AC, CV6 AC, CV8 AC, CV9 AC, CV10 AC, CV21 AC, NO3 AC, NO4 AC, CTO3 AC, CTO15 AC, CTO16 AC, CTO20 AC, CTO21 AC) were developed in DMEM with 10% fetal bovine serum (FBS) for conventional monolayer growth and named adherent cells (AC). Both types of cultures were maintained in a humidified atmosphere of 5% O2 and 5% CO2.
Cell line authentication from the respective primary tumor was obtained by short tandem repeat (STR) profiling.
Two malignant glioma cell lines (U87 MG and 010627) were kindly supplied by Dr. Rossella Galli (DIBIT San Raffaele, Milan, Italy) and maintained as AC and NS, respectively.
All experiments on primary glioblastoma cell lines were carried out with cells from passages 10-20. All cultures were checked for Mycoplasma contamination before experimental use (e-Myco™ Mycoplasma PCR Detection kit; iNtRON Biotechnology Inc., Gyeonggi-do, South Korea).
Immunofluorescence. Expression of key WNT/β-catenin pathway molecules was examined on a panel of 12 glioblastoma cell lines (seven NS and five AC). Cells were fixed for 20 min with 4% paraformaldehyde at room temperature, rinsed three times with phosphate-buffered saline (PBS), blocked/permeabilized for 30 minutes with 1× PBS containing 2% of the appropriate serum and 0.1% Triton X-100 and, finally, stained with the primary antibodies listed in Table II. Negative controls were obtained by omitting the primary antibody. Alexa Fluor® 488-AffiniPure goat anti-rabbit IgG and Alexa Fluor® 594-AffiniPure rabbit anti-mouse IgG (Jackson ImmunoResearch Laboratories Inc., West Grove, PA, USA) were used as secondary antibodies. Cell nuclei were stained with 4’,6-diamidino-2-phenylindole (DAPI). Images were examined and acquired on a Zeiss Axioskop fluorescence microscope (Karl Zeiss, Oberkochen, Germany) equipped with an AxioCam5MR5c and coupled to an imaging system (AxioVision Release 4.5, Zeiss).
On CTO3 NS, CTO3 AC, NO3 NS and NO3 AC lines, immunofluorescence was also carried out after treatment with 50 μM temozolomide (Sigma Aldrich Co., St. Louis, MO, USA) for 48 h.
Immunocytochemistry. The analysis was performed using a Ventana Full BenchMark® XT automated immunostainer (Ventana Medical Systems Inc.) and UltraView™ Universal DAB Detection Kit (Ventana Medical Systems Inc.) as detection system. HIER was performed in Tris-EDTA (pH 8). Primary antibodies were the same used for immunofluorescence with the same dilutions. Negative controls were obtained by omitting the primary antibody. Images were examined and acquired on a Zeiss Axioskop fluorescence microscope.
Protein extraction and western blotting analysis. Whole-protein extracts from six cell lines as NS and six as AC were obtained using a RIPA lysis buffer containing protease and phosphatase inhibitor cocktails (Sigma Aldrich Co.). After protein quantification (BCA™ Protein Assay; Pierce Biotechnology, Rockford, IL, USA), 30 μg of total protein extracts were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene difluoride membranes (Immobilon-P; Millipore, Bedford, MA, USA) with standard procedure. Primary antibodies used were anti-WNT3a (NBP1-19050, dilution 1:1000; Novus Biologicals, Littleton, CO, USA) and anti-β-catenin (610153, dilution 1:500; BD Transduction Laboratories™, Franklin Lakes, NJ, USA); secondary antibodies were horseradish peroxidase-labeled (Dako, Glostrup, Denmark). Proteins were detected by enhanced chemiluminescence using Immobilon Western reagents (ECL; Millipore). A rabbit polyclonal antibody to human α-tubulin (LF-PA0146, dilution 1:5,000; LabFrontier, Seoul, Korea) was used to normalize sample loading and transfer. Band intensities were quantified using NIH Image J software (National Institutes of Health, Bethesda, MD, USA).
In order to analyze the subcellular localization of β-catenin, nuclear and cytoplasmic fractions of frozen tissues and cells were prepared using the NE-PER® Nuclear and Cytoplasmic Extraction reagents (Pierce Biotechnology) as per manufacturer's instruction. Thirty micrograms of protein for cells and 50 μg for tissue analysis were subjected to western blotting as described above, using antibody to β-catenin. Antibody to human α-tubulin and an anti–TATA-box binding protein (TBP/TFIID; Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) were used as loading controls for cytosolic and nuclear fractions, respectively.
DNA extraction. Genomic DNA (gDNA) from frozen or FFPE tumor samples and cell lines was isolated using the QIAamp® DNA Mini kit (Qiagen Inc., Valencia, CA, USA), according to the manufacturer's instructions. Constitutional gDNA was extracted from peripheral blood by a salting-out protocol and from saliva using the Oragene® DNA Collection kit (DNA Genotek Inc., ON, Canada, USA).
CTNNB1 mutation analysis. CTNNB1 exon 3 encodes the N-terminal regulatory sequence of the β-catenin protein that contains the consensus phosphorylation sites for GSK-3β critical for its proteasomal degradation. Two primer pairs designed on gDNA were used to amplify CTNNB1 (GenBank reference sequence NM_001098209) exon 3 and the adjacent splice sites (including at least 100 bp of the flanking intronic sequences) by polymerase chain reaction (PCR). The primer sequences are available on demand. PCR amplification was performed in a total volume of 10 μl containing 50 mM KCl, 10 mM Tris–HCl (pH 8.3), 1.5 mM MgCl2, 250 μM of each dNTP, 0.365 U of AB Taq Polymerase (AB Analitica, Padova, Italy), 10 pmol of each primer and 50 ng of genomic DNA. A standard touchdown PCR protocol was used.
Sanger direct sequencing. Prior to sequencing, PCR products were purified by 0.5 U shrimp alkaline phosphatase (SAP) and 5 U exonuclease I (GE Healthcare BioSciences, Auckland, New Zealand), according to manufacturer's instructions. Cycle sequencing was performed using the BigDye® Terminator v1.1 Cycle Sequencing Kit (Thermo Fisher Scientific Inc.). Sequencing products were analyzed by Sanger sequencing on an ABI® 3130 Genetic Analyzer and data collected by Sequencing Analysis v.5.3.1 software (Thermo Fisher Scientific Inc.).
RNA isolation and quantitative real-time PCR (qRT-PCR). Total RNA from frozen tissues and cell lines was isolated using the RNeasy® Mini Kit (Qiagen Inc.). One microgram of RNA was reverse transcribed into cDNA using random primers with the High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific Inc.), according to the manufacturer's protocol.
qRT-PCR for WNT3A was performed on a StepOne Real-Time PCR system using the StepOne software v2.3 (Thermo Fisher Scientific Inc.). Reactions were performed in triplicate using TaqMan® Gene Expression Master Mix and TaqMan® Gene Expression Assay (WNT3A Hs00263977_m1) (both from Thermo Fisher Scientific Inc.).
A suitable endogenous reference gene was validated by comparing the stability of three different housekeeping genes [glyceraldehyde-3-phosphate dehydrogenase (GADPH), beta-2-microglobulin (B2M) and hypoxanthine guanine phosphoribosyltransferase 1 (HPRT1)] in a series of glioblastoma tumor specimens. Relative fold changes of WNT3A gene expression were then normalized against that of the endogenous HPRT1 gene and determined by calculation of the 2−ΔΔct. A commercial FirstChoice® human brain total RNA (Ambion®, Thermo Fisher Scientific Inc.) was used as reference and two normal nervous tissues as internal control.
Statistical methods. Associations between categorical variables were evaluated using 2×2 contingency tables by the two-tailed Fisher's exact test. The Pearson's correlation coefficient was used to examine correlations among TCF4, cyclin D1 and Ki-67/MIB-1 labeling indices (LIs).

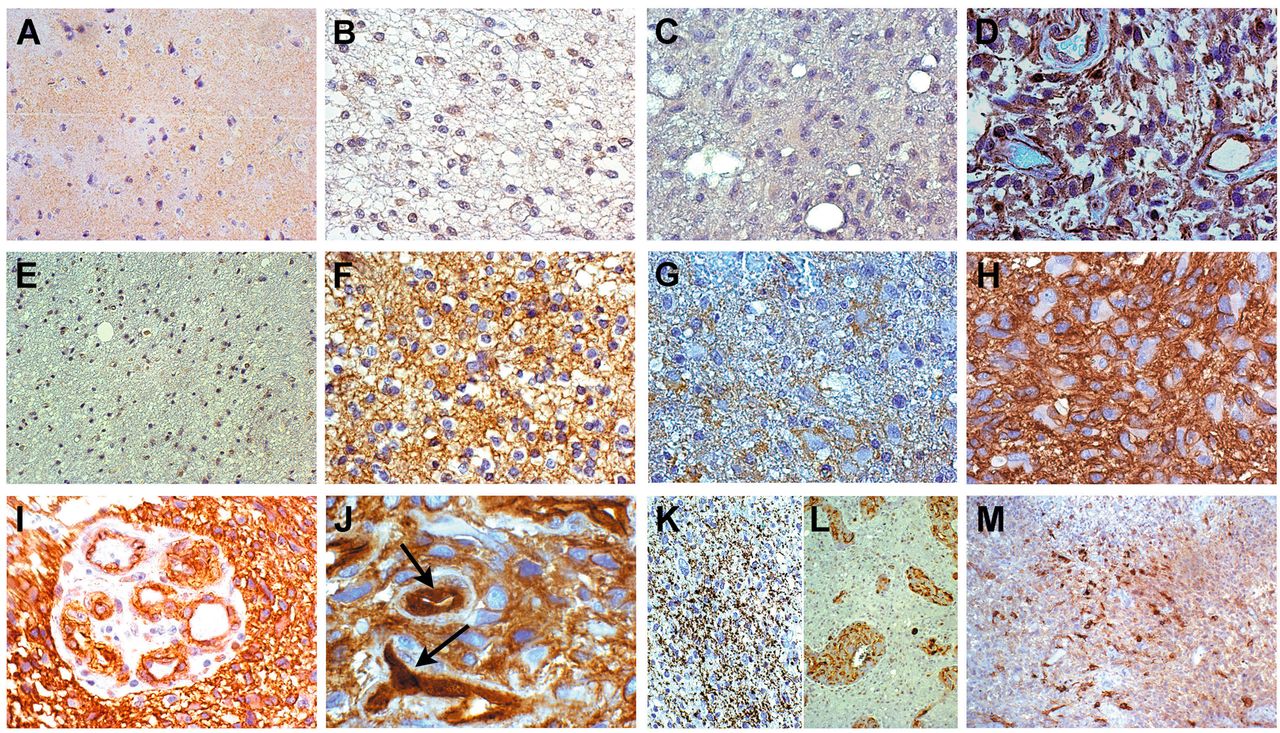
Immunohistochemistry of Wingless-type MMTV integration site family (WNT)/β-catenin signaling pathway. A: Normal cortex. No WNT3a expression, ×200. B: Grade II oligodendroglioma. Moderate WNT3a expression on the cell membranes, ×400. C: Grade II astrocytoma. No WNT3a expression, ×400. D: Glioblastoma. Diffuse cytoplasmic and membranous WNT3a expression, ×400. E: Normal white matter. Slight membranous β-catenin expression, ×200. F: Grade II oligodendroglioma. Diffuse membranous β-catenin expression, ×400. G: Grade III astrocytoma. Moderate cytoplasmic and membranous β-catenin expression, ×400. H: Glioblastoma. High cytoplasmic and membranous β-catenin expression, ×400. I: Glioblastoma. Cytoplasmic and membranous β-catenin expression in proliferative endothelial cells, ×200. J: Glioblastoma. Nuclear and cytoplasmic β-catenin expression in hyperplastic endothelial cells, as indicated by arrows, ×630. K: Grade III astrocytoma. Granular expression of phospho-β-catenin, ×200. L: Glioblastoma. Phospho-β-catenin expression in the cytoplasm of proliferative endothelial cells, ×200. M: Glioblastoma. Positivity for phopsho-GSK-3β, ×200. All specimens stained with 3,3’ diaminobenzidine.
Overall survival (OS) was defined as the time between the histological diagnosis and patient's death or last follow-up. Patients alive at last follow-up were considered censored events. Survival curves were estimated using the Kaplan–Meier method and differences between them were compared by the log-rank test (Mantel–Cox). Analysis was carried out by SPSS v21.0 software (SPSS Inc., Chicago, IL, USA). p-Values <0.05 were considered as statistically significant.
Results
Immunohistochemistry. The immunohistochemical demonstration of WNT3a expression was technically very difficult because of its variability. However, WNT3a was negative in the normal nervous tissue (Figure 2A), occasionally and slightly positive on the cell membranes of grade II and III oligodendrogliomas (Figure 2B), negative in grade II and III astrocytomas (Figure 2C), whereas it was highly expressed in the cytoplasm and on the cell membranes of glioblastomas (Figure 2D). WNT3a expression was not found in pilocytic astrocytomas.
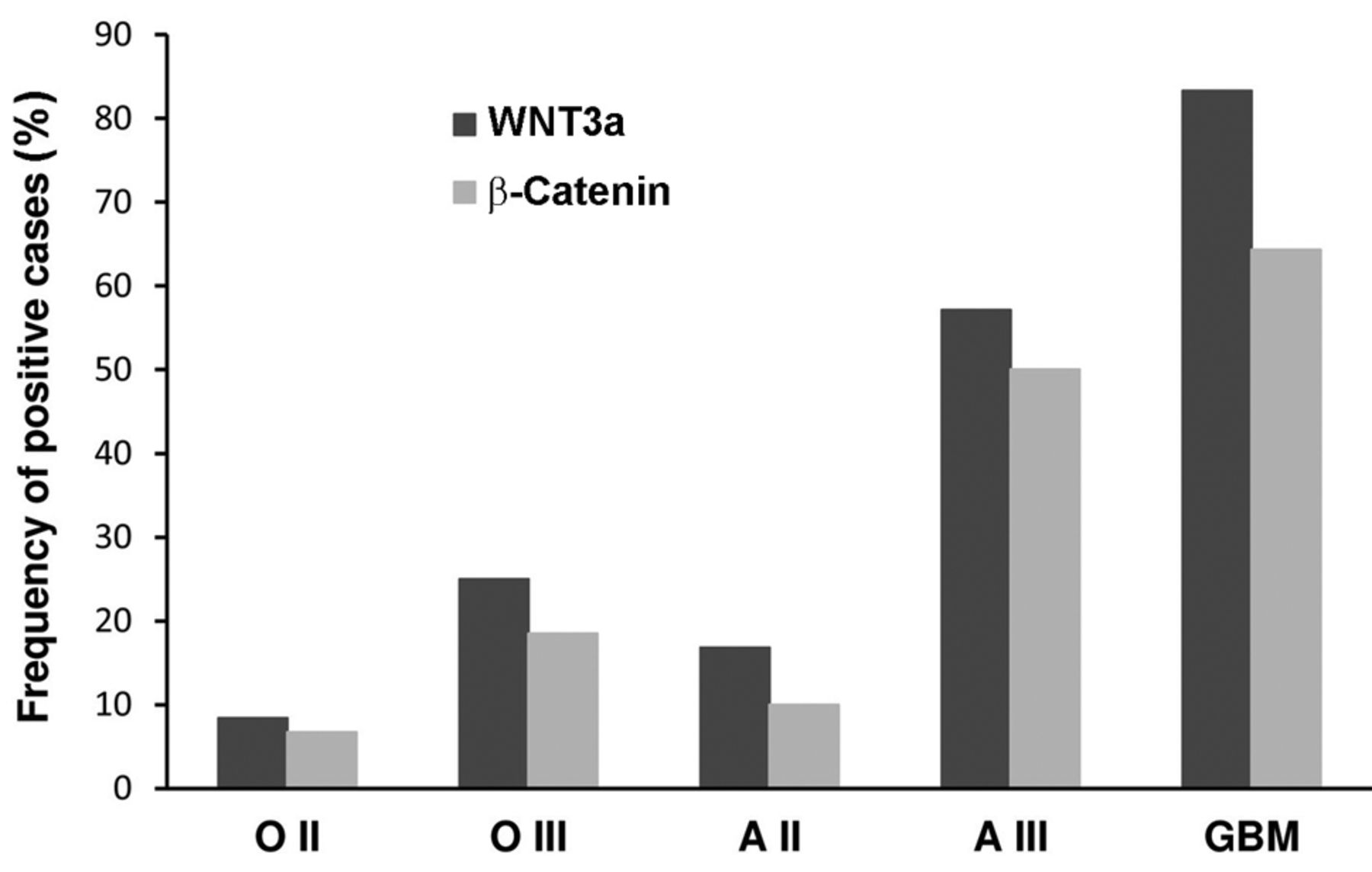
β-Catenin was weakly expressed in the normal nervous tissue only on the cell membranes (Figure 2E). It was expressed in the cytoplasm and on the cell membranes more intensely with increasing histological grade (Figure 2F-H). The immunostaining intensity was higher and prevailingly cytoplasmic in high- rather than in low-grade astrocytomas, while it was limited to the cell membranes in oligodendroglial tumors. β-Catenin expression was not found in pilocytic astrocytomas. Clear-cut nuclear staining was not found, with the exception of rare, scattered and barely identifiable positive nuclei. In proliferative endothelial cells of microvascular proliferation (MVPs), cytoplasmic and membranous positive staining was diffusely present (Figure 2I). Occasionally, nuclear staining was detected, more frequently and reliably compared to tumor cells (Figure 2J). The cytoplasmic accumulation of β-catenin was regarded as indicative of aberrant pathway activation. Phospho-β-catenin (Ser33/37 /Thr41), considered as a destabilized and inactive form of β-catenin, was variably expressed in all gliomas and in endothelial cells with a granular staining pattern, inversely correlated to the histological grade (Figure 2K and L). Expression of phospho-GSK-3β (Tyr216), the active form of GSK-3β in the degradation complex of β-catenin, followed the immunostaining pattern of phospho-β-catenin (Figure 2M). The frequency of WNT3a and β-catenin immuno histochemical expression among gliomas of different tumor types and histological malignancy grade is reported in Figure 3.
TCF4 was expressed in nuclei of tumor cells, with a variable staining intensity, and more in high- than in low-grade gliomas (Figure 4A and B). Nuclei with the highest staining intensity corresponded to those positive for Ki-67/MIB-1 (r=0.761, p<0.0001) and cyclin D1 (r=0.739, p<0.0001) (Figure 5). Nuclear cyclin D1 expression was significantly higher in high- than in low-grade gliomas and correlated with the Ki-67/MIB-1 LI (r=0.873, p<0.0001) (Figure 4C and 5). By visual analysis of serial sections, the areas with the maximum Ki-67/MIB-1 LI corresponded to those positive for cyclin D1 and with the highest TCF4 expression (Figure 4D and E). Nuclei intensely positive for TCF4 were also detected in proliferative endothelial cells (Figure 4F). TCF4 expression was found neither in the normal nervous tissue nor in pilocytic astrocytomas.
Immunofluorescence. In NS, WNT3a was variably positive in the cytoplasm (Figure 6A), β-catenin was more consistently positive in the cytoplasm and on the cell membranes (Figure 6B), and TCF4 consistently positive in the nuclei (Figure 6C); phospho-GSK-3β was variably positive in the cytoplasm (Figure 6D). In AC, WNT3a was slightly positive, and β-catenin and TCF4 negative in the cytoplasm, that, in contrast, was positive for phospho-GSK-3β.
After treatment with 50 μM temozolomide for 48 h, WNT3a, β-catenin, TCF4 and phospho-GSK-3β expression was revealed in AC (Figure 6E-H), thus demonstrating the pathway activation. Their expression did not change in NS.
Immunocytochemistry. The immunocytochemical expression patterns of the key molecules studied confirmed the observations obtained by immunofluorescence (Figure 6I and J). In NS, β-catenin expression was also clearly appreciable as a nuclear staining (Figure 6K). TCF4 was also positive in the nuclei of AC (Figure 6L).
Relationships of WNT3a and β-catenin protein expression with clinical and molecular features. The WNT3a and β-catenin protein expression was not associated with sex, patient age, tumor location or age at diagnosis in the whole series of gliomas (p>0.05 for all categories). In contrast, their expression was significantly associated with glioblastomas compared to low-grade gliomas (p=0.0113 and p=0.0349, respectively for WNT3a and β-catenin).

Wingless-type MMTV integration site family member 3a (WNT3a) and β-catenin immunohistochemical expression with respect to tumor type and histological malignancy grade.
The comparison of WNT3a and β-catenin expression with typical genetic and epigenetic alterations of gliomas [epidermal growth factor receptor (EGFR) gene amplification, O6-methylguanine-DNA methyltransferase (MGMT) promoter hypermethylation, isocitrate dehydrogenase 1/2 (IDH1/2), tumor protein p53 (TP53), phosphatase and tensin homolog (PTEN) and telomerase reverse transcriptase (TERT) mutations], already published (44, 45), revealed a significant association of WNT3a immunoreactivity with TERT promoter mutations (p=0.0208).
Western blotting. Western blotting analysis of whole-cell lysates from glioblastoma cell lines revealed clear bands at approximately 40 kDa for WNT3a and 92 kDa for β-catenin. The two proteins were expressed variably and more intensely in NS than in AC (Figure 7A). Densitometric quantification of the bands is shown in Figure 7B.
Upon fractionated extraction, nuclear expression of β-catenin was detected in both NS and AC (Figure 7C). In fractionated glioblastoma tissue extracts, β-catenin expression was appreciable in the nuclei, although to a lesser extent compared to cytoplasmic expression (Figure 7D).
CTNNB1 mutation analysis. Search for mutations in CTNNB1 exon 3 and in the adjacent splice sites was performed in all malignant astrocytic tumor samples and glioblastoma cell lines. Mutation analysis revealed genetic mutations neither in tumor samples nor in glioblastoma cell lines.
qRT-PCR. By qRT-PCR, the WNT3A expression level was evaluated in 11 glioblastomas and six grade II-III tumor specimens, in 24 glioblastoma cell lines (10 NS and 14 AC), and in two normal nervous tissue samples. When available, the analysis was performed on NS and AC developed from the same primary tumor (n=4). A fold change in the relative gene expression of more than two was regarded as evidence of WNT3A up-regulation.

Correlations among transcription factor 4 (TCF4), Ki-67/MIB-1 and cyclin D1 labeling indices (LIs). A: Glioblastoma. High nuclear TCF4 expression in several tumor cells, ×200. B: Grade II oligodendroglioma. TCF4 intensely positive nuclei, ×400. C: Glioblastoma. Several cyclin D1-positive nuclei in the same area as in (A), ×200. D: Several Ki-67/MIB-1-positive nuclei in the same area as in (A), ×200. E: Several Ki-67/MIB-1-positive nuclei in the same area as in (B), ×400. F: Glioblastoma. Nuclei of proliferative endothelial cells with high TCF4 expression, as indicated by arrows, ×400. All stained with 3,3’ diaminobenzidine.
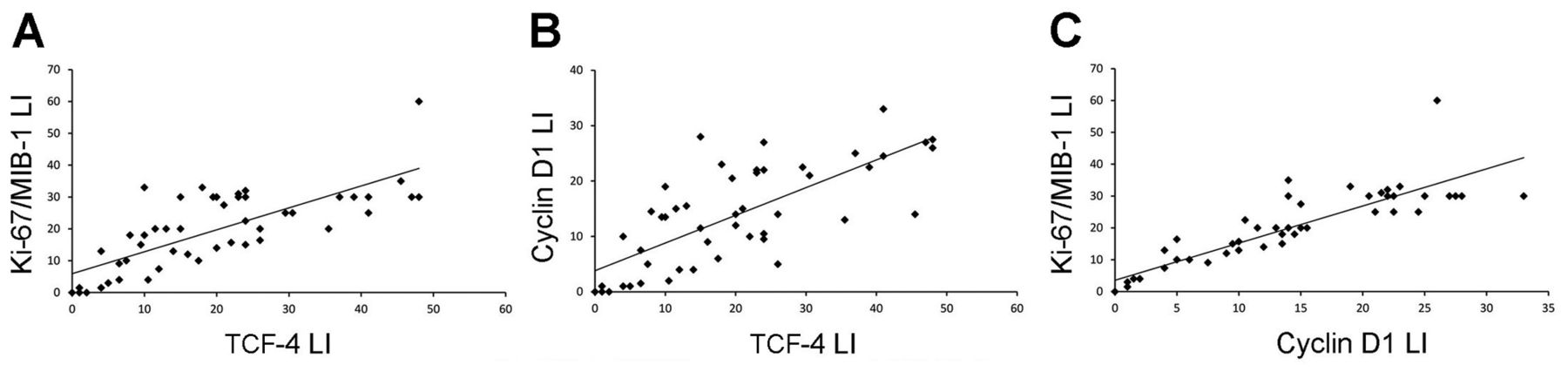
Correlation among transcription factor 4 (TCF4), Ki-67/MIB-1 and cyclin D1 labeling indices (LIs). A: Linear regression analysis between TCF4 and Ki-67/MIB-1 LIs (r=0.761, p<0.0001). B: TCF4 and cyclin D1 LIs (r=0.739, p<0.0001). C: Cyclin D1 and KI-67/MIB-1 LIs (r=0.873, p<0.0001).
WNT3A expression levels were significantly up-regulated in glioblastomas (7/11, 63.6%) and in anaplastic astrocytomas (2/2, 100%), with increasing fold change from grade III to grade IV (Figure 8A). Variable and weak WNT3A expression was detected in grade II and III astrocytic and oligodendroglial tumors. Compared to the normal nervous tissue, glioblastomas showed up-regulation of WNT3A level up to 80-fold and grade II-III gliomas up to 9-fold. The up-regulation rate observed in glioblastoma tumor specimens was consistent with previous data from the Cancer Genome Atlas portal, documenting WNT3A overexpression in 55% of glioma samples (25).

Immunofluorescence (IF) and immunocytochemistry (ICC) of glioblastoma stem cells. IF, Neurospheres: A: High cytoplasmic Wingless-type MMTV integration site family member 3a (WNT3a) expression. B: High cytoplasmic β-catenin expression. C: High nuclear transcription factor 4 (TCF4) expression. D: Cytoplasmic phopsho-glycogen synthase kinase-3β (GSK-3β) expression. IF, Adherent cells: E: Very poor cytoplasmic WNT3a expression. F: Negative β-catenin expression. G: Negative TCF4 expression. H: Positive phopsho-GSK-3β expression. IF, Adherent cells after 50 μM temozolomide treatment for 48 h: I: Positive cytoplasmic WNT3a expression. J: Positive cytoplasmic β-catenin expression. K: Nuclear and cytoplasmic TCF4 expression. L: Poor cytoplasmic phopsho-GSK-3β expression. Nuclei were counterstained with 4’,6-diamidino-2-phenylindole. All ×400 magnification. ICC, Neurospheres: M: Cytoplasmic WNT3a expression. N: Cytoplasmic and nuclear β-catenin expression. O: Nuclear TCF4 expression. ICC, Adherent cells: P: Prevailing nuclear TCF4 expression. All 3,3’ diaminobenzidine, ×400 magnification.
In glioblastoma cell lines, WNT3A up-regulation was detected in 8/24 (33.3%) cases, with a similar distribution in NS (3/10, 30%) and AC (5/14, 35.7%) (Figure 8B). By comparing NS and AC with the matched primary tumors, WNT3A gene expression levels were equally up-regulated in NS and in the primary tumor, and they were, generally, higher in NS than in AC. The maximum fold change observed in the CTO3 NS was in agreement with its high proliferative rate (data not shown). WNT3A gene expression was undetectable in the cell lines derived from WNT3A-negative primary tumors. Higher relative fold changes were observed in AC obtained from tumors that did not develop NS from primary cultures.
Weak WNT3A overexpression was found in 010627 NS, whereas it was not detected in U87MG AC, as reported in the literature (24, 25). In the two normal nervous tissue samples, WNT3A was not expressed.
Survival analysis. The relationship of WNT3a and β-catenin expression with OS was evaluated in a sub-group of 25 patients including 16 glioblastomas, seven grade II and two grade III gliomas. By univariate analysis, WNT3a (p=0.033) and cytoplasmic or cytoplasmic/nuclear β-catenin (p=0.042) expression were significantly associated with a worse prognosis in the whole group of gliomas (Figure 9). The median survival time was 16 months for WNT3a-negative cases and 10 months for WNT3a-positive cases. Censored cases were 4/11 (36.4%) for the former and 2/14 (14.3%) for the latter. This was expected since WNT3a and β-catenin correlated with malignancy.

Western blot analysis. A: Representative analysis of two independent experiments displaying Wingless-type MMTV integration site family member 3a (WNT3a) and β-catenin expression in whole-cell extracts from glioblastoma cell lines, cultured as neurospheres or adherent cells. B: Quantitative analysis of WNT3a and β-catenin levels normalized to α-tubulin, showing higher amounts of the two proteins in neurospheres than adherent cells. C, D: β-Catenin expression by western blotting analysis in cytosolic and nuclear extracts from glioblastoma cell lines and tumor tissues.
Discussion
The canonical WNT/β-catenin signaling pathway plays a pivotal role in tumorigenesis and progression of malignant astrocytic gliomas (10-15), increases the expression of stem cell genes (35-40), promotes the migratory and invasive capacity of glioblastomas, and influences epithelial to mesenchymal transition activators (16, 46-48). Our findings are in line with these observations.
The key step in the pathway is the cytoplasmic stabilization of β-catenin and its translocation to the nucleus to form a β-catenin/LEF/TCF complex that regulates the transcription of multiple genes involved in cellular proliferation, differentiation, survival and apoptosis, such as FRA1, c-MYC, and cyclin D1. Importantly, the oncogenic function of β-catenin primarily depends on its subcellular localization, with its nuclear accumulation being responsible for malignant progression (49).

WNT3A expression levels by quantitative real-time polymerase chain reaction in gliomas and glioblastoma cell lines. A: Relative WNT3A expression levels in glioma tumor samples of different types and histological malignancy grades. B: Relative WNT3A expression levels in a panel of 24 glioblastoma cell lines, cultured as neurospheres (NS) or adherent cells (AC). Reported values are the average ± standard deviation (SD) of at least two independent experiments in triplicate.

Relationship of Wingless-type MMTV integration site family member 3a (WNT3a) and β-catenin expression with survival in glioma patients. Kaplan–Meier survival curves for overall survival (OS) with respect to WNT3a expression (A) and subcellular localization of β-catenin expression (B). M: membranous; C: cytoplasmic; N: nuclear.
In our series, we report on the aberrant pathway activation in malignant astrocytic gliomas by demonstrating the overexpression of the WNT3a ligand at both the protein and mRNA level. At the same time, high protein expression levels of β-catenin and TCF4 were detected in anaplastic astrocytomas and glioblastomas, but not in diffuse astrocytomas or oligodendrogliomas, compared to normal nervous tissue. WNT3a, β-catenin and TCF4 expression was prevalent in astrocytic than in oligodendroglial tumors with a significant correlation with the histological malignancy grade, as reported in the literature (10-13, 18, 19, 25, 26, 28). Contrary to high-grade glioma, where the cytoplasmic subcellular localization of β-catenin prevails, in low-grade glioma, β-catenin expression is mainly limited to the cell membranes, thus suggesting a more structural than oncogenic function.
In our material, immunohistochemical detection of WNT3a was rather unsatisfactory, even though occasionally positive on the cell membranes of high-grade gliomas. However, WNT3A up-regulation was demonstrated by qRT-PCR in nine out of 11 (81.8%) malignant astrocytic gliomas and in eight out of 24 (33.3%) glioblastoma cell lines, as in previous observations (23-25). On the other hand, in agreement with the activation of the WNT/β-catenin pathway in glioma progression, frequent aberrant promoter hypermethylation of WNT pathway inhibitor genes has been reported in malignant astrocytic gliomas, with secreted frizzled-related protein (SFRP1, SFRP2) and naked (NKD2) inhibitors prevailing in primary glioblastomas (40%) and dickkopf (DKK1) inhibitor in secondary glioblastomas (50%) (50). Another inhibitor of the pathway is the WNT inhibitory factor 1 (WIF1) gene, epigenetically silenced in 75% of glioblastomas (51-54).
One crucial point concerns the immunohistochemical demonstration of the nuclear translocation of β-catenin. In our series, in the absence of clear nuclear expression, cytoplasmic immunopositivity for β-catenin was regarded as indicative of WNT/β-catenin signaling pathway activation, as for previous authors (14, 17). Indeed, the cytoplasmic stabilization of β-catenin precedes the nuclear translocation and the subsequent transcription of target genes. In fresh/FFPE malignant astrocytic gliomas, nuclear or nuclear/cytoplasmic positivity of β-catenin has been found (10-14, 16, 19). In our experience, the scattered and rare nuclear expression was always found in association with the cytoplasmic expression. Technical problems may have hampered the immunohistochemical detection of nuclear β-catenin in FFPE tumor samples. As a matter of fact, western blotting analysis successfully revealed β-catenin expression in the nuclear fractions from fresh glioblastoma tumor samples and glioblastoma cell lines. The nuclear translocation of β-catenin in glioblastoma cell lines has already been reported (11-13, 18, 19, 24-26), also in medulloblastoma cell lines (55).
In endothelial cells of MVPs, the cytoplasmic and membranous expression of β-catenin was easily detectable. It has been already observed in proliferative endothelial cells of glioblastomas and related to angiogenesis and cell proliferation (56). Expression of TCF4 in nuclei of endothelial cells is in line with this interpretation. TCF4-positive nuclei of hyperplastic endothelial cells significantly correlated with the Ki-67/MIB-1 LI.
TCF4 is the main effector of the WNT/β-catenin signaling pathway in the control of gene regulation. At its maximum immunostaining intensity, it statistically correlated with the histological malignancy grade and with Ki-67/MIB-1 and cyclin D1 LIs (11, 13, 26, 28). Therefore, it might be used as a proliferative marker both in astrocytic and oligodendroglial glioma. On the other hand, TCF4 expression levels correlated with V-AKT murine thymoma viral oncogene homolog 1 and 2 (AKT1 and AKT2), both direct target genes of TCF4. Down-regulation of β-catenin/TCF4 activity thus inhibits glioma cell proliferation and invasion in vitro and tumor growth in vivo (13).
Phospho-GSK-3β and phospho-β-catenin, both instrumental for the proteasomal degradation of β-catenin, had a similar expression pattern, opposite to that for oncogenic WNT3a and β-catenin, and inversely correlated to histological grade.
Key components of the canonical WNT/β-catenin signaling pathway are commonly affected by genetic mutations in several cancer types (29). Thus, we focused on the search for potential activating mutations in CTNNB1 exon 3 that encodes the N-terminal regulatory sequence of β-catenin, containing the consensus phosphorylation sites for GSK-3β, and reported as a frequent event in different tumor entities. Unlike other malignancies, however, the absence of CTNNB1 activating mutations excludes genetic events as responsible for the oncogenic activation of β-catenin in malignant astrocytic gliomas, as previously observed (14, 31).
The WNT/β-catenin signaling may contribute to the maintenance of stem cell-like properties, inhibition of differentiation, radioresistance and development of an invasive phenotype in glioma (35-37, 57). As a matter of fact, the canonical WNT/β-catenin signaling is crucially involved in embryonic development and controls stem cell biology by inducing self-renewal of both embryonic stem cells (ESCs) and adult stem cells (21, 58-60). NANOG, octamer-binding transcription factor 4 (OCT4), SRY (sex determining region Y)-box 2 (SOX2) and c-MYC, transcriptional factors responsible for pluripotency and self-renewal in ESCs, are all TCF4 target genes (61, 62). The activity of c-MYC and cyclin D1 target genes is promoted by the downstream modulator of the WNT/β-catenin pathway Forkhead box M1 (FOXM1) (24). Knock-down of FOXM1 and β-catenin in glioma stem cells (GSCs) abolishes their self-renewal and tumor-initiation abilities (34).
A novel proto-oncogene, pleomorphic adenoma gene-like 2 (PLAGL2), induces activation of WNT receptors and ligands and increases the accumulation of β-catenin. Enhanced PLAGL2 expression by gene amplification strongly suppresses differentiation of normal neural stem cells and GSCs, while it promotes their self-renewal capacity (36). Another gene regulating the WNT/β-catenin signaling in glioma is paternally expressed gene 3 (PEG3), an imprinted gene with a tumor -suppressor activity. Aberrant PEG3 promoter hypermethylation was significantly associated with high-grade glioma. Low PEG3 expression increases β-catenin expression and thus proliferation of GSCs (63). Consistently with these observations, no wonder, therefore, that WNT3a and β-catenin are variably expressed in NS and AC, more in the former than in the latter, as demonstrated by immunofluorescence, immunocytochemistry, western blotting and qRT-PCR. The reduced expression of WNT3a and β-catenin in AC is in line with their more differentiating character.
Furthermore, WNT/β-catenin pathway activity confers chemoresistance to cancer stem-like cells in different tumor types (64, 65), glioblastomas included (39, 40). In our series, treatment with temozolomide induced activation of the WNT/β-catenin pathway in AC, in line with previous observations (25). Thus, the inhibition of the pathway may represent a potential tool to enhance the response of GSCs to temozolomide.
Our findings also demonstrate that WNT3a and cytoplasmic/nuclear β-catenin expression were significantly associated with a worse prognosis in patients with malignant glioma (12, 14, 17-19), as in other cancer types (66, 67). WNT3a and β-catenin may thus be regarded as potential prognostic factors in the entire group of gliomas.
WNT3a and β-catenin expression was not associated with any of the well-known genetic and epigenetic features of gliomas, except for TERT promoter mutations.
Finally, the canonical WNT/β-catenin signaling pathway can be regarded as a stemness regulator and enhancer of cell proliferation in gliomas, strongly associated with the histological malignancy grade.
Conclusion
The canonical WNT/β-catenin signaling pathway contributes to tumorigenesis and progression of malignant astrocytic gliomas and promotes stem cell properties. TCF4 activates transcription of target genes and significantly correlates with the Ki-67/MIB-1 LI. The crucial point in the pathway is the nuclear translocation of β-catenin.
Acknowledgements
This work was supported by Grant N. 4011SD/cv 2011-0438 of Compagnia di San Paolo, Turin, Italy.
Footnotes
Conflicts of Interests
The Authors declare that they have no conflict of interests.
- Received September 21, 2015.
- Revision received October 10, 2015.
- Accepted November 4, 2015.
- Copyright© 2016, International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Proteogenomics of glioblastoma associates molecular patterns with survival
- WNT vampirization by glioblastoma leads to tumor growth and neurodegeneration
- Investigation of dacomitinib on reducing cell necrosis and enhancing cell apoptosis in C6 glioma rat model by MRI
- Moesin Up-regulation Is Associated with Enhanced Tumor Progression Imaged Non-invasively in an Orthotopic Mouse Model of Human Glioblastoma
- Anticancer Activity of Artemisinin and its Derivatives