


Abstract
Background: Prostate cancer (PC) represents the most frequently diagnosed cancer in men. Exposure to infectious agents has been considered to induce prostatic inflammation and cancerous transformation. Controversial data exist concerning the role of the human polyomaviruses BK (BKV) and JC (JCV) in PC etiology. Therefore, a possible association between these polyomaviruses and PC was investigated. Materials and Methods: Urine, blood and fresh prostatic tissue specimens were collected from 26 patients with PC. The presence of BKV and JCV, the possible non-coding control region (NCCR) variations and the genotyping analysis of viral protein 1 (VP1) of both viruses were assessed. Results: Data showed a preferential viral re-activation in the urinary compartment and a statistically significant prevalence of JC viruria and of BKV in PC tissues. A BKV DDP-like NCCR sequence was isolated in two patients, whereas JCV NCCR was consistently of an archetypal structural organization. A prevalence of the European genotypes was observed for both viruses. Conclusion: Our data demonstrated the presence of JCV DNA in 14/24 (58.3%) cancerous prostatic tissue specimens, confirming the results obtained in a previous study, in which JCV has been defined as common inhabitant of the prostate, and opening the discussion about its potential role in PC.
With an estimated 220,800 new cases and 27,540 deaths in the United States during 2015, prostate cancer (PC) represents the second leading cause of cancer-related death and the most frequently diagnosed cancer in men aside from skin cancer (1). Along with benign prostatic hyperplasia (BPH), the most frequent urological diagnosis in elderly males, PC is a significant health concern that may become increasingly prevalent in the coming years in relation to the gradual aging of the population (2). In fact, increasing age is one of the well-established risk factors that contribute to the development of PC (1).
Currently, there is both experimental and clinical evidence validating the hypothesis that inflammation may contribute to the pathogenesis of PC (3). Inflammation has been associated with about 20% of all human cancer in adults by causing genomic damage (2, 4-7). Infection is a known major cause of inflammation, and can lead to chronic inflammation of the prostate. In fact, different infectious agents (for instance sexually transmitted bacteria and viruses) have been shown to affect the prostate, which is a sterile organ, inducing prostatic inflammation (2, 5, 8-11) and eventually cancerous transformation of the prostate gland, mainly when this phenomenon is prolonged (3).
Several non-sexually transmitted infectious agents have also been investigated in relation to prostate carcinogenesis, including those of the genitourinary tract, such as the human polyomavirus BK (BKV) (10). This urotheliotrophic virus exhibits oncogenic properties via the large T antigen, which can bind and block the p53 tumor-suppressor and the retinoblastoma protein (pRb) (12). Because of the long-term latency, genitourinary tropism and potential oncogenicity that are associated with BKV, several articles have investigated its possible link to prostate cancer, with controversial results (13-25). However, the possibility of ‘hit-and-run’ carcinogenic mechanisms induced by BKV cannot be excluded (18).
BKV genome is functionally divided into two coding regions: an early region, which codes for the T antigens (large and small), and a late region, which codes for the late proteins (agno and capsid proteins). It also presents a non-coding control region (NCCR), which contains several cis-acting sequences important for viral DNA replication [promoter/enhancer elements, binding sites for T antigens, origin of replication (O-block, 142 bp)], and sequences containing the binding sites for cellular transcriptional factors involved in transcriptional regulation of both the early and the late genes (26). The NCCR of the proposed archetypal BK strain (WW) has been arbitrarily divided into four transcriptional factor binding sequence blocks, called P (68 bp), Q (39 bp), R (63 bp), and S (63 bp) (27). The different BKV strains display a marked heterogeneity in their NCCRs due to point mutations, deletions, duplications, and re-arrangements in this region. These re-arrangements may play an important role in virus replication, allowing the virus to adapt to ongoing changes within the host cell environment, by increasing or decreasing the number or the affinities of host transcription factor binding sites (28). Genetically re-arranged BKV variants (e.g. URO1) might exist in the urinary tract and be responsible for neoplastic transformation of prostate cells (29). Therefore, BKV has been indicated as a potential cofactor in the earliest stages of PC (18). In fact, our previous studies also demonstrated that the presence of BKV and its variants in transformed cells does not exclude viral pressure in cell immortalization (19, 25).
Regarding the human polyomavirus JC (JCV), despite its high homology with BKV, the presence of the oncogenic protein large T antigen and its latency site in the genitourinary tract, only a few studies have investigated the possible role of this virus in relation to PC, obtaining conflicting results (13, 16, 21, 24). The JCV NCCR is a highly variable region and contains many neurotropic and neurovirulence determinants. Based on the structure of this region, the JCV NCCR forms have been classified into four types (30). Type II-S has a sequence made up of boxes A (25 bp), B (23 bp), C (55 bp), D (66 bp), E (18 bp) and F (69 bp), as seen in the archetype CY (31). Type I-S is composed of boxes A (25 bp), C (55 bp) and E (18 bp), forming a single 98-bp unit, and box F (69 bp). Conversely, type I-R has repeats of this 98 bp unit with various deletions, as seen in JCV Mad-1 and Mad-4 strains (32-33), respectively. Finally, type II-R has repeats of this 98 bp unit and inserts with various mutations, also called rearranged NCCRs. In types I-R and II-R is always present the box F (33). The NCCR of the JCV Mad-1 strain has been detected in the prostate gland associated with the urinary tract together with different NCCR sequences, suggesting that the prostate may be inhabited by more than one JCV strain or NCCR variant (13).
Taking into account these assumptions, the aims of this molecular study were: (i) to investigate whether the presence of BKV might be linked to PC progression, (ii) to assess a possible association between JCV and PC, considering its high homology with BKV, and (iii) to determine whether a specific NCCR configuration and a particular viral genotype were associated with a normal or cancerous prostate cellular environment. In particular, the presence of both human polyomaviruses was evaluated in fresh prostatic adenocarcinoma and in benign tissue specimens, in urine, plasma and peripheral blood mononuclear cells (PBMCs) collected from each patient enrolled in this study.
Materials and Methods
Study population. Fresh prostate tissue specimens, blood and urine samples were obtained from 26 patients (mean age=66 years; range=51-75 years), without a history of neoplastic diseases. These subjects underwent radical retropubic prostatectomy for prostate cancer treatment at the Department of Obstetrics, Gynecology and Urological Sciences of the Umberto I General Hospital (Rome, Italy) between December 2011 to April 2013 (Table I). Enrollment was achieved according to the following criteria: (i) no previous hormonal or radiation therapy, (ii) no previous surgery of the prostate gland, and (iii) histologically-proven PC by biopsy and confirmed by radical retropubic prostatectomy.
Prostate specimens, obtained at surgery, were analyzed for areas of relatively pure prostatic adenocarcinoma, or benign tissue, which served as benign controls, and processed independently. All tissue samples were immediately frozen and stored at −80°C until DNA extraction.
All patients enrolled in this study provided written informed consent with guarantees of confidentiality.
Clinical specimens collection and processing. Standard laboratory procedures for sterile DNA extraction and polymerase chain reaction (PCR) were practiced for all specimens, including the use of ultra-pure, UV irradiated and autoclaved water, autoclaving of all tubes and equipment, tissue processing under a laminar flow, UV irradiated hood, the use of filter-barrier pipette tips, etc.
Fresh urine specimens, collected without preservatives, were centrifuged at 1.376 × g for 10 min at 4°C. DNA for molecular analysis was extracted from the pellet of each specimen using the QIAamp® DNA Mini Kit (QIAGEN S.p.A, Milan, Italy) according to the manufacturer's instructions.
Blood samples, collected in 7-ml Vacutainer® tubes containing EDTA (BD Becton Dickinson S.p.A, Milan, Italy), were centrifuged at 1.376×g for 10 min and DNA was extracted from 200 μl of plasma using the QIAamp® DNA Mini Kit (QIAGEN S.p.A). PBMCs were isolated from whole blood using the standard Ficoll Hypaque density gradient centrifugation technique (34) and the number of viable leukocytes was determined by trypan blue exclusion. PBMCs DNA extraction was performed on 106 cells by the QIAmp® DNA Mini Kit (QIAGEN S.p.A) following the manufacturer's instructions.
Small pieces of normal and cancerous prostate tissue specimens were thawed and processed for DNA extraction using the QIAamp® DNA Mini Kit (QIAGEN S.p.A) and the recommended tissue protocol. DNA yield of all samples was determined by measuring its concentration in the eluate by absorbance at 260 nm and DNA was stored at −20°C until use.
Clinical characteristics of patients with prostate cancer and clinical samples provided.
BKV/JCV real-time TaqMan PCR (Q-PCR). DNA extracted from each sample was tested for BKV and JCV genome and quantification by two commercial kits: the BKV Q-PCR Alert Kit and the JCV Q-PCR Alert Kit (ELITechGroup S.p.A, Trezzano S/N, Milan, Italy). These quantitative assays, based on TaqMan-MGB® (Minor Groove Binder) technology, were performed using a 7300 real-time PCR system and SDS 1.4 software to analyze the amplification data (AB Applied Biosystems, Foster City, CA, USA). The amplification reaction was carried out both for a region of the gene that encodes BKV and JCV large T antigen and for the promoter region and 5’UTR of the human beta-globin (β-globin) gene. β-Globin gene was amplified simultaneously with the target sequence to verify successful DNA isolation and exclude false-negative results. All samples were tested in triplicate and the number of viral copies in each sample was calculated from the standard curve. The standard curve for these quantitative amplification assays was obtained using four stabilized serial dilutions at known titer (range: 102-105 plasmid copies) of a plasmid containing part of the BKV and JCV large T antigen. For urine and plasma specimens, BKV and JCV DNA load was expressed as genome equivalents of viral DNA (gEq)/ml of sample, as genome equivalents (gEq)/106 cells of sample for PBMCs and as genome equivalents (gEq)/mg for tissues. Standard precautions designed to prevent contamination during Q-PCR were followed and a no template control (NTC) lane was included in each run. Both assays were able to quantify between 10 and 106 DNA copies of the target gene per amplification reaction (gEq).
PCR for BKV/JCV NCCR and VP1 gene typing region. All samples positive for viral DNA were subsequently screened for the NCCR of both viruses by nested-PCR using specific sets of primers (35-36) in a 9700 GeneAmp® PCR System (AB Applied Biosystems). To characterize the viral genotype in all BKV/JCV-positive samples, a PCR amplifying the VP1 typing region of both viruses was carried out with specific primers for these regions (37-39). All assays included positive (previously amplifiable BKV or JCV DNA) and negative (pure water as template) controls to exclude false-positive and false-negative results. PCR products were detected by electrophoresis on a GelRed™ (Biotium)-stained 2% agarose gel and visualized under UV light.
Sequencing of BKV/JCV NCCR and VP1 regions. PCR products corresponding to the NCCR and VP1 regions were purified before sequencing to remove excess primers with QIAquick® PCR purification kit, according to the QIAGEN protocol (40). DNA sequencing was performed using a dedicated facility (Bio-Fab research s.r.l., Rome, Italy). All sequences obtained from the NCCR amplifications were compared to the BKV prototypic NCCR sequence (41) and to the JCV NCCR of the archetype strain CY (31) in order to investigate the presence of possible rearrangements within these regions. Sequences obtained from the amplification of the VP1 gene typing region of both viruses were classified into known genotypes/subtypes examining the single nucleotide polymorphisms (SNPs) within the amplified VP1 coding sequence. To detect the SNPs by which we have classified our isolates, many complete VP1 gene sequences belonging to all known BKV subtypes/subgroups and JCV genotypes were aligned in order to generate a consensus sequence for each BKV subtype and JCV genotype (28, 42). Subsequently, we aligned our isolates with the consensus sequences generated for each BKV subtypes by Anzivino and colleagues in 2011 (28) and with those reported by Jobes and colleagues in 2001 for JCV (42). Sequence alignments were performed with ClustalW2 at the EMBL-EBI website using default parameters (43).
Summary of polyomavirus detection and characteristics in all clinical specimens.
Statistical analysis. Data were summarized as medians and ranges or as the mean, as appropriate. If the Z-test indicated a non-normal distribution, we used non-parametric test such as Mann-Whitney U-tests and Kruskal–Wallis tests. Categorical data were analyzed by using χ2 test and Student's t-test. p-Values of less than 0.05 were considered statistically significant.
Results
Evaluation of JC and BK viral load in biological specimens. This study was conducted on 26 patients with PC from whom we collected at surgery 52 fresh prostate specimens (26 benign and 26 cancerous tissue samples), 24 urine and 24 whole blood samples. Regarding human polyomavirus DNA detection by Q-PCR in urine, BK viruria was detected in seven out of 24 specimens (29.2%) with a mean load of 2.97 log10 gEq/ml (range=2.34-4.85 gEq/ml), whereas JC viruria was found in 15/24 samples (62.5%) with a mean value of 5.16 log10 gEq/ml (range=3.43-6.51 gEq/ml) (Table II). Concerning blood samples, BKV DNA was detected in 6/24 plasma samples (25%), with a mean viral load of 3.25 log10 gEq/ml (range=2.44-4.25 gEq/ml), and in two PBMCs specimens (8.3%) with a load of 1.15 log10 gEq/106 cells (range=1.11-1.18 gEq/106 cells). JC viremia was found in two out of 24 samples (8.3%), with a mean load of 3.81 log10 gEq/ml (range=3.76-3.86 gEq/ml), whereas 14/24 specimens (58.3%) were positive for JCV DNA in PBMCs samples, with a mean value of 1.29 log10 gEq/106 cells (range=1.11-1.45 gEq/106 cells) (Table II). Finally, in prostate specimens, BKV was found in 6/26 cancerous tissue samples (23.1%), with a value of 3.35 log10 gEq/mg (range=2.26-3.90 gEq/mg), and in one benign tissue sample, with a value of 1.54 log10 gEq/mg, whereas JCV DNA was detected both in 14 cancerous and in 14 benign tissue samples (58.3%), with a mean load of 2.04 log10 gEq/mg (range=1.6-2.79 gEq/mg) and of 1.99 log10 gEq/mg (range: 1.56-3.13gEq/mg), respectively (Table II).
Human polyomaviruses characteristics in all clinical specimens provided according to Gleason score.
Statistical analysis of these data highlighted that the average number of genome equivalents of BKV detected in cancerous prostate tissues was significantly higher than that observed in JCV-positive ones (p=0.002) (Table II). A statistically significant difference between BKV and JCV loads in malignant tissues was observed even when the results were categorized according to the Gleason score 6 and 7. In fact, the mean BKV load reported in the Gleason 7 class was significantly higher than that stated for JCV (p=0.013) (Table III). However, the difference among the mean JC and BK loads was not statistically significant in the Gleason 6 class, although the number of BKV genome equivalents in cancerous prostatic tissue specimens was higher than that of JCV (p=0.053) (Table III).
Conversely, the average number of BKV genome equivalents detected in urine was significantly lower than that observed in JCV-positive urine (p=0.001) (Table II). Even when the data were partitioned into the two Gleason score classes, the BKV load in urine specimens was significantly lower than that of JCV in both classes (Table III). However, the comparison between the mean values of JCV and BKV loads in urine by Gleason score was not statistically significant, although the number of genome equivalents decreased from those with Gleason score 7 to those with Gleason score 6 (data not shown).
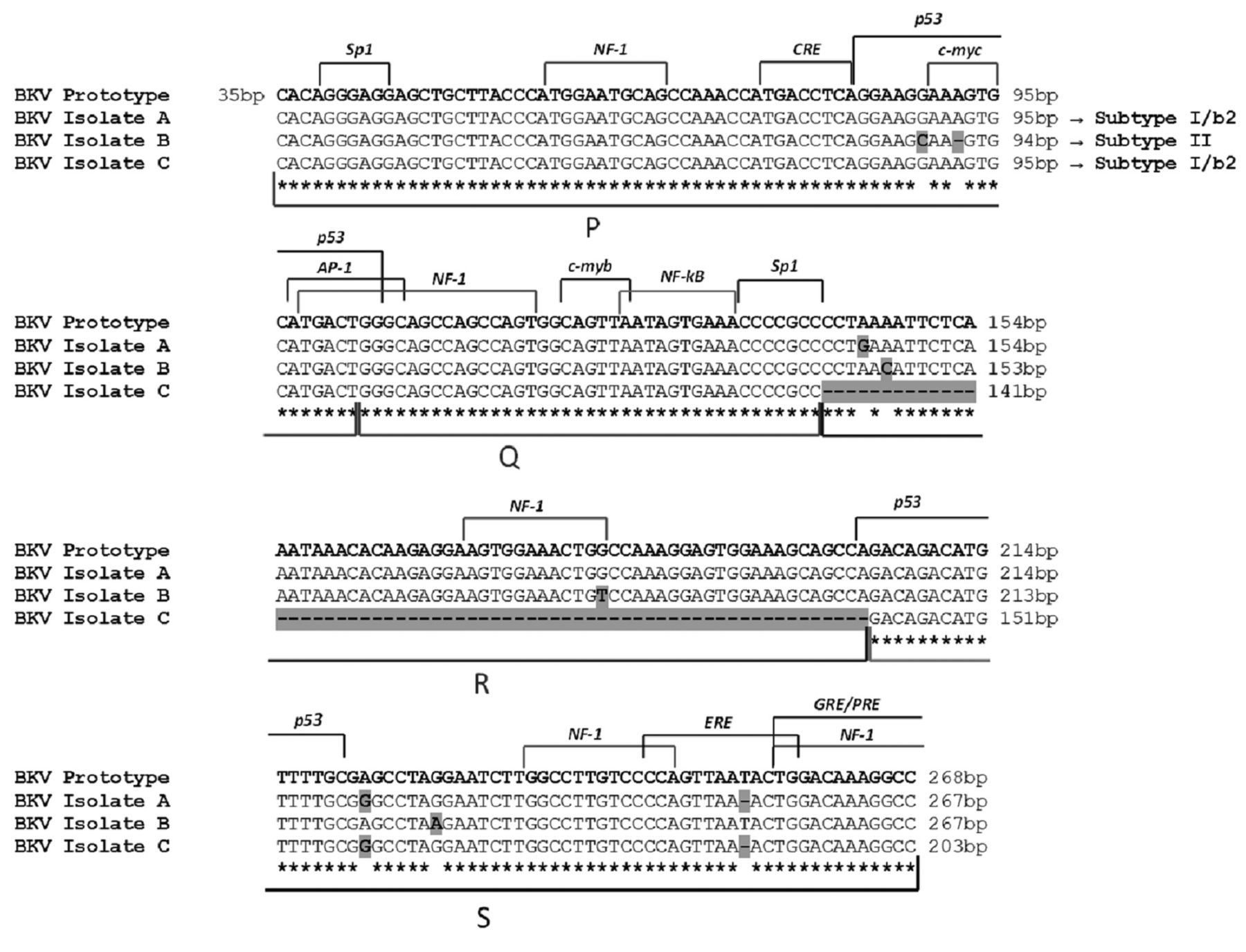
Alignment of BKV NCCR isolates with the prototypic NCCR sequence. The prototypic NCCR defined by Yogo and colleagues (41) is shown at the top of the figure, while nucleotide sequences of three variants, determined directly from prostatic tissue, urine and blood samples, are shown below in relation to the prototypic NCCR, with nucleotide substitutions indicated by bold highlighted letters and deletions identified by bold highlighted hyphens. The genotype/subtype corresponding to each isolate is also indicated. Blocks (P, Q, R and S) commonly used to denote archetypal NCCRs (27) are indicated under the NCCR sequences, while proven and putative binding sites for transcriptional factors are illustrated above the prototypic NCCR sequence. The O-block, containing the origin of replication, was omitted.
Finally, categorizing the results according to the number of patients and type of virus, JCV was detected in 22 out of 26 (84.6%) of the individuals, whereas BKV in 11 out of 26 (42.3%). Two patients were negative for both viruses. Therefore, JCV was more common than BKV in the cohort of patients enrolled in this study.
Analysis of JCV and BKV NCCR sequences. All the specimens that were positive for viral DNA in the initial screening were further analyzed for the presence of re-arrangements within the NCCR region of both viruses. The NCCR sequences amplified in each BKV-positive clinical sample were compared to the prototypic NCCR defined by Yogo et al. (41), which spans from the midpoint of the origin of replication and the start site of the agno protein gene. Specifically, the 268 nucleotides of the prototypic NCCR sequence were arbitrarily divided into five blocks (O, P, Q, R, and S) with lengths of 35, 68, 39, 63, and 63 nucleotides, respectively, to describe the major rearrangements within this region more easily.
Homology sequence analysis evidenced that the BKV NCCR sequence isolated from the urine of four patients was identical to the prototypic NCCR, consisting of a single complete copy of the five blocks arranged in the correct order, with the occurrence of a few point-mutations at nucleotide positions 4 within R-block and 18 within S-block, and a thymidine nucleotide deletion at nucleotide position 50 within S-block. These point mutations did not involve any cellular transcriptional factors binding site, except for the single-nucleotide deletion at position 50 within S-block involving the estrogen response element (Figure 1, isolate A). A prototypic NCCR structure was also isolated from three urine and four plasma specimens belonging to five patients. All sequences were characterized by various nucleotide substitutions at nucleotide positions 55 within P-block, 7 and 41 within R-block, 24 within S-block and by the deletion of a single adenine nucleotide at nucleotide position 58 within P-block. The nucleotide substitutions at position 55 within P-block and 41 within R-block involved the binding sites for the cellular factors p53 and NF-1, respectively, whereas the adenine deletion at position 58 within P block interested the binding site for the transcriptional factor c-myc (Figure 1, isolate B). Finally, two patients showed a rearranged NCCR structure: the former in benign prostatic tissue samples, whereas the latter in plasma and PBMCs specimens. This re-arranged NCCR consisted of a whole R-block deletion and of a perfectly conserved single copy of the remaining P, Q and S blocks arranged in the correct order. The R-block deletion results in the loss of the NF-1 binding site. This re-arranged sequence was also characterized by the same two nucleotide changes within the S-block found in isolate A: the nucleotide transition at position 18, which did not involve any cellular transcriptional factors binding site, and the thymidine deletion at nucleotide position 50, involving the estrogen response element binding site (Figure 1, isolate C). This rearranged sequence was similar to the DDP strain of BK virus isolated from PBMCs by Degener and colleagues (44).

Examples of nucleotide sequence diversity in JCV NCCR isolates from different clinical specimens of patients with PC. The NCCR sequence of the archetypal JCV strain CY, as proposed by Yogo et al. (31), is shown at the top of the figure and the consensus sequences of the NCCR isolated from cancerous or normal prostatic tissues (CPT and NPT, respectively), blood and urine specimens of three patients with PC, with related genotype, are shown below. The four nucleotide changes (G108A, A133C, G213C, G217A), identified by comparing with CY isolate, are indicated by bold highlighted letters. The JCV NCCR isolated from specimens of the other patients always included the nucleotide substitution G217A (data not shown). The six boxes commonly used to divide the archetypal NCCR (33) are indicated under the NCCR sequences, while proven and putative binding sites for transcriptional factors are illustrated above the CY NCCR sequence.
Regarding JCV, the archetypal variant type II-S, with a few single-nucleotide substitutions, has been found in all samples. In particular, four nucleotide changes (G108A, A133C, G213C and G217A), identified in the NCCR from 10 isolates of three patients by comparing with the archetypal JCV CY strain, have been described previously by Agostini et al. (13) and Zambrano et al. (45) and correspond to sequence polymorphisms (Figure 2). The nucleotide changes G108A and A133C did not involve any cellular transcriptional factors binding site, whereas the nucleotide substitutions G213C and G217A involved nuclear factor-1(NF-1) binding site. The NCCR sequences isolated from specimens of the remaining JCV-positive patients were all characterized by the 217-G to A nucleotide transition inside the box F (data not shown).
PCR and sequence analysis of BKV/JCV VP1 region. An additional specific nested-PCR was undertaken to detect the presence of VP1 typing region in all BKV- and JCV-positive samples. Regarding BKV, the 327 bp VP1 gene sequence was amplified in clinical specimens belonging to 11 patients (Table II). The amplified PCR products were sequenced in order to classify each BKV strain into the corresponding subtype/subgroup, analyzing the SNPs within the amplified VP1 region and aligning our 327 bp typing isolates with the consensus sequences generated for each BKV subtype/subgroup (28). Based on the SNPs and on the consensus sequences, subtype I/subgroup b-2 was detected in six patients, while subtype II was detected in five individuals.
The SNPs and the consensus sequence, generated from the alignment of 32 complete JCV genomes (42), were used to determine the JCV genotype of our positive samples. In particular, among the 26 enrolled patients, a prevalence of the European genotypes 1A, 1B and 4, followed by the African genotype 3A, was observed. In particular, genotypes 1A and 1B were found in six and in 13 individuals, respectively, whereas type 4 was identified in one patient. Finally, two patients carried the African genotype 3A.
Discussion
The human prostate is a organ recognized without normal flora. However, different infectious agents have been shown to affect the prostate and cause both asymptomatic and symptomatic diseases, especially prostatitis. Currently, there is both experimental and clinical evidence validating the hypothesis that infection-induced chronic inflammation of the prostate may be one of the many causes of PC. Moreover, there is an indication that men who have used anti-inflammatory drugs have a reduced risk of PC, and that the use of antibiotics may result in a declined rate of chronic prostate inflammation. This further demonstrates a strong link between infection and chronic prostate inflammation, suggesting that infectious agents may have a link to prostate carcinogenesis (3). However, the limited number of studies conducted over the past few years to define viral infections as a risk factor in PC development has yielded rather inconclusive data (22, 24).
BK virus is among several non-sexually transmitted infectious agents investigated in relation to prostate carcinogenesis. BKV, a member of the Polyomaviridae family, is widely distributed in the human population. Following primary infection, typically acquired during early childhood, the virus disseminates into various organs and establishes a persistent infection, predominantly in the kidney and the genitourinary tract, including the prostate (8, 10, 22). Since BKV possesses potentially oncogenic properties, and it is associated with long-term latency and genitourinary tropism, several studies investigated its possible link to PC with controversial results (13-25). In the literature, there are reports on a positive correlation between BKV DNA and PC (14, 15, 18, 19, 24, 25). Among these, Das et al. (18), Russo et al. (19), Delbue et al. (24) and Mischitelli et al. (25) found that BKV DNA positivity was more common in malignant than in normal prostatic tissue. However, the intriguing hypothesis that BKV might contribute to prostate carcinogenesis has not been supported by the results of other molecular studies: one observed no difference in BKV DNA positivity between malignant and benign tissue specimens (29), whereas others observed minimal or no evidence of BKV DNA in prostate tissue specimens (16, 17, 20-23). In this study, the percentage of BKV DNA-positive cancerous prostate tissues (23%) is lower than those reported in our previous reports (19, 25), in which BKV genome was found in 22/26 (85%) and in 31/71 (44%) fresh PC specimens, respectively. However, this percentage is similar to that reported by Balis and colleagues (15), who revealed the presence of BK virus in 19% of all PC cases. It is not clear why there has been such a variation in the percentage of BKV DNA-positive malignant prostate tissues among our three study populations, as we used the same methodological assay to evaluate BKV load in clinical specimens. This discrepancy may be due to the different number of Gleason score classes in these studies. In fact, in Russo et al. (19) and in Mischitelli et al. (25) the data were scheduled into four Gleason score classes, spanning from Gleason 6 and Gleason 9, whereas in this study they were categorized in only two classes: Gleason 6 and 7. The main BKV DNA positivity in PC biopsies was found in the highest Gleason score classes (8 and 9), whereas the lowest frequencies of viral positivity were reported within the classes with Gleason score 6 and 7, especially in that with Gleason 6. The latter result is in agreement with data reported by this study, in which the lowest BKV positivity and mean load were detected in patients with a Gleason score of 6 (Table III). This led us to speculate that BKV might interfere with the control of cellular proliferation, promoting the acquisition of a complete ‘neoplastic phenotype’ and tumor progression. Moreover, the possibility of a ‘hit and run’ carcinogenic mechanism induced by BKV cannot be excluded (18). Even the prevalence of BKV DNA in plasma and urine specimens from patients with PC belonging to this study group (viremia 25% and viruria 29.2%) was lower with respect to that (viremia 31% and viruria 54%) detected by Russo et al. in 2008 (19). Nevertheless, both studies demonstrated that the urinary compartment was the preferential site of BKV replication.
Despite the high structural homology between BKV and the human polyomavirus JC, the latency site in the genitourinary tract and the potential oncogenicity associated with these viruses, only a few studies have investigated JCV in relation to PC obtaining contradictory data (13, 16, 21, 24). Therefore, we extended this study to investigate a possible causative role of JCV in PC development. Our results showed the presence of JCV DNA in 58.3% of cancerous and benign prostatic tissue samples, whereas JC viruria and viremia were found in 62.5% and 8.3% of patients with PC, respectively. Similarly to BKV, major replication in the urinary compartment was observed. However, JC viruria was significantly higher than that of BKV (Table II), even when the results were categorized according to Gleason score (Table III). Therefore, JCV was more common than BKV in the cohort of patients with PC enrolled in this study, even though the average number of BKV genome equivalents in cancerous prostatic tissue specimens was higher than that of JCV (p=0.002).
These results are consistent with the data reported by Zambrano et al., who detected JCV in 50% of 12 patients with PC either in prostatic tissues or in urine specimens (13). Two independent studies, in contrast, did not find significant differences in JCV detection between cancerous and benign prostatic tissue specimens screened (16, 24). Similarly, a Mexican study did not detect JCV sequences in any of the 55 prostate cancer biopsies analyzed (21). Discrepancies among these studies, including ours, may be explained by technical differences in the viral detection methods and differences in sample size. Actually, the difficulty in obtaining fresh surgical tissues limited the statistical considerations of our data due to the low numbers of samples collected for this study and the previous one (19). In fact, only one biopsy specimen of normal and cancerous prostatic tissue from each patient was tested.
The purpose of the histological division of the prostatic tissue specimens was to examine whether a specific BKV and JCV NCCR configuration was associated with a normal or cancerous cellular environment. As mentioned above, the NCCR of both human polyomaviruses BK and JC is a highly variable region, which may undergo sequence rearrangements yielding viral variants that could up-modulate viral expression in a specific anatomical site or acquire new tissue tropism and greater pathogenic potential. Monini and colleagues reported the detection of BKV sequences by PCR and Southern blotting in more than 50% of both normal and tumor tissues obtained from the urinary tract and prostate (29). The sequence analysis of these BKV regulatory regions showed that these tissues were infected by a novel BKV strain, URO1. URO1 strain, which has a high prevalence in the urinary tract of Italian patients, is characterized by NCCR rearrangements that may affect viral DNA replication. However, it remains to be determined whether the mutations in URO1 have an effect on replication or transformation (46). Recently, Mischitelli et al. found two rearranged BKV variants, characterized by enhancement of the oncogenic c-myc and the oncosuppressor p53 binding sites, in cancerous tissues of two patients affected by advanced prostate cancer (25).
In this study, a DDP-like NCCR sequence was isolated from the normal prostatic tissue and the blood (plasma and PBMCs) of two BKV-positive patients. This rearranged NCCR consisted of a whole R-block deletion and of a perfectly conserved single copy of the remaining O, P, Q and S blocks arranged in the correct order (Figure 1). The simultaneous presence of a rearranged NCCR sequence in plasma and in PBMCs of the same patient could likely be explained by the occurrence of an alteration in the BKV NCCR structure during persistent infection in the circulatory compartment. This led us to speculate that in this case, the BK variant found in plasma could derive from the rearranged one recovered in PBMCs. This suspicion was confirmed by the NCCR sequencing of the viral variant isolated from the urine of this patient. In fact, the excreted virus showed a prototypical structural organization of its regulatory region, consisting of a single complete copy of all five blocks (O, P, Q, R and S).
The molecular analysis of the BKV VP1 coding sequence between nucleotides 1744 and 1812 (amino acids 61 to 83) allows definition of the BKV genotypes and their different distribution in human populations (37). In particular, subtype I (further divided into four subgroups, each of which has a unique geographical distribution pattern: I/a, I/b-1, I/b-2 and I/c) is widespread throughout the world, subtype IV (further divided into six subgroups with their own geographical distribution pattern: IV/a-1, IV/a-2, IV/b-1, IV/b-2, IV/c-1 and IV/c-2) is prevalent in East Asia and part of Europe, while subtypes II and III are rarely detected throughout the world (47). In this study the European subtype I/b-2 and the subtype II were mainly detected. Interestingly, all BKV subtypes/subgroups are defined by the presence of specific nucleotide substitutions and deletions within the NCCR sequence (41). In fact, alignment of all NCCR sequences, isolated from each BKV-positive clinical sample of our patients with PC, with the prototypic NCCR sequence proposed by Yogo and colleagues (41), revealed that the point mutations detected in these sequence, including the rearranged ones, were typical of the consensus NCCR sequences identified for the BKV subtype I/subgroup b-2 and subtype II.
Concerning JCV, Laghi et al. (48) and Ricciardiello et al. (49) demonstrated that the pathogenic JCV Mad-1 strain was the only JCV variant present in the human colon and that a variant with a single 98-bp sequence within the NCCR is found exclusively in the cancer tissues, suggesting that this Mad-1-like strain might be implicated in the development of chromosomal instability in the colon epithelium. It is noteworthy that JCV Mad-1 was also detected in the prostate gland associated with the urinary tract, where archetypal strains would be expected. Different NCCR sequences were also found in prostatic tissue together with JCV Mad-1, suggesting that prostate may be inhabited by more than one JCV strain or NCCR variant (13). In our study, when JCV was present in histologically different areas, the NCCR sequencing showed that in all cases, the virus was identical to the archetypal variant type II-S in both normal and cancerous areas (Figure 2). The finding of archetypal NCCR structures within our cancerous and benign prostate tissues led us to speculate that the non-pathogenic CY strain of JCV might be selected for replication and persistently infect the prostate gland, mostly the portion associated with the urinary tract. Alterations in the prostatic microenvironment could allow the virus to actively replicate and select for one or more JCV strains or NCCR variants that are capable of transforming the host tissue. Ongoing replication of the virus might also lead to viral and cellular modifications that could promote genomic instability, contributing to tumorigenesis or cancer progression. This hypothesis is supported by Zambrano et al., who observed different NCCR variants in the prostate tissue and in urine of one patient (13). It is possible that JCV could mainly be involved in cancer progression rather than in tumorigenesis, since we detected the virus in an archetypal form in prostate tissues belonging to patients affected by Gleason 7 PC. However, this suspicion must be confirmed by further studies.
Four nucleotide changes (G108A, A133C, G213C and G217A) were detected in the NCCR from 10 isolates of three patients by comparing with the archetypal JCV CY strain (31). All these changes corresponded to known sequence polymorphisms (13, 45), and two of them (G213C and G217A) involved the NF-1 binding site (Figure 2). The 217-G to A nucleotide transition inside box F was also present within the NCCR sequences isolated from specimens of the other JCV-positive patients. This point-mutation seems to be a common feature of the European strains and was also found in European Americans (45). Indeed, in our study group, there was a prevalence of the European genotypes 1A, 1B and 4, followed by the East African genotype 3A.
Although in this study the sample size was small, our data demonstrate the presence of JCV DNA in approximatively 60% of tumor and benign prostatic tissue samples, confirming the conclusions of a previous study in which JCV was defined as a common inhabitant of the prostate. Even though the detection of viral nucleic acids within the tumor tissue is crucial, but not sufficient, in demonstrating any potential involvement of viral infection in human cancer, the finding of a high frequency of JCV DNA in prostate tissue opens the discussion about its potential role in PC. However, more studies are necessary to ascertain how a mainly neurotropic virus such as JCV could operate in PC development and progression, especially through rearrangements of its regulatory region.
Acknowledgments
This work was supported by MIUR grant.
Footnotes
Conflicts of Interest
The Authors declare that they have no competing interests.
- Received April 10, 2015.
- Revision received May 18, 2015.
- Accepted May 20, 2015.
- Copyright© 2015, International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved




{kind=link}
{kind=link}