


Abstract
NME1 is a well-documented metastasis suppressor gene, with suppressor activity demonstrated across a wide spectrum of human cancers including melanoma and carcinomas of the breast, stomach and thyroid. A primary aim of the current study was to identify profiles of genes whose expression is regulated by NME1 in cell lines of melanoma and thyroid carcinoma origin. Impact of NME1 was determined by forcing its expression transiently in cell lines using a novel Ad5-based adenoviral vector (Ad5-NME1), followed 48 h later by analysis of RNA expression profiles using the U133A microarray chip. Robust NME1 expression was achieved following infection with the Ad5-NME1 adenovirus in the human metastasis-derived cell lines WM1158 (melanoma) and WRO82 (follicular thyroid carcinoma), resulting in wide-ranging effects on gene expression in both settings. A substantial proportion of the NME1-regulated genes identified in the analyses were of clear potential relevance to metastasis, such as matrix metalloproteinase-1 (MMP1), angiopoietin-2 (ANGPT2), SERPINB9 and colony stimulating factor receptor-2B (CSFR2B). Nine genes were identified (false discovery rate <0.1) that were regulated by NME1 in both the WM1158 and WRO82 cell lines, each possessing one or more such metastasis-relevant activities as stress fiber formation and focal adhesion (PPM1E, ZYX, PFN1), chemotaxis (CCR1) epithelial-mesenchymal signaling (WNT6), differentiation and morphogenesis (TBX4, ZFP36L2), and G protein modulation (GPR52 and PFN1). In addition, a number of the NME1-regulated genes were shown to be of prognostic value for distant disease-free survival and overall survival in melanoma and breast cancer. The combined expression of three NME1-regulated genes CSFR2B, MSF4A1 and SERPINB9 provided a strongly synergistic correlation with distant disease-free survival in the basal subtype of breast cancer (p<3.5e-5, hazard ratio=0.33). Our study demonstrates that analysis of NME1-dependent gene expression is a powerful approach for identifying potential modulators of metastatic potential in multiple cancer types, which in turn may represent useful therapeutic targets. The study also highlights NME1-dependent genes as potential prognostic/diagnostic indices, which are profoundly lacking at present in melanoma.
Metastasis accounts for most of the morbidity and mortality associated with cancer, yet the underlying mechanisms are poorly understood (1). A better understanding of those processes could provide new therapeutic approaches to block cancer dissemination, as well as targeting metastatic lesions that have already been established. Metastasis represents a complex cascade of events that requires a coordinated expression of gene networks to overcome barriers to their dissemination and growth in heterotopic environments. A growing number of metastasis-suppressor genes and their encoded proteins are being identified that inhibit metastasis selectively, with minimal impact on growth of the primary tumor. Their metastasis-specific functions are providing critical insights into events driving the metastatic cascade in many cancers (2, 3). We recently verified metastasis suppressor activity of the NME1 and NME2 genes in melanoma for the first time in vivo, using a transgenic mouse model of UV-induced melanoma (4). Our site-directed mutagenesis analysis of the NME1 molecule has identified amino acid residues critical for its nucleoside diphosphate kinase and 3′-5′ exonuclease activities (5) and metastasis suppressor function (6).
Considerable evidence suggests NME proteins regulate gene expression, although the underlying mechanisms involved have yet to be fully elucidated. Early studies demonstrated an affinity of NME1 and NME2 for single-stranded motifs in the CMYC (7) and PDGFA (5, 8) promoter regions, as well as their abilities to regulate transcription from those promoters. Chromatin immunoprecipitation analyses have further demonstrated physical associations of NME1 with non-B form DNA elements in the promoter regions of these and the TP53 genes (9-11). In addition, NME1 has been shown to regulate global gene expression profiles in the breast carcinoma cell line, MDA-MB-435, with the lysophosphatidic acid receptor EDG2 identified as a motility-driving target of NME1-mediated suppression (12). A primary intent of the current study was to analyze NME1-dependent gene expression profiles in the specific setting of melanoma, which had yet to be explored. In addition, we included thyroid carcinoma, for which a number of studies have indicated reduced NME1 expression in metastases and advanced forms of the disease (13-15). An important rationale for studying NME1-dependent gene expression in both melanoma and thyroid carcinoma cell lines was the potential for discovery of genes regulated across these diverse cancer types, which in turn could potentially enrich for genes with fundamental roles in metastasis suppressor function. This study has used a novel adenoviral vector for efficient forced expression of NME1, and microarray analysis of resulting gene expression profiles to reveal broad effects of NME1 on gene expression in metastatic cell lines of melanoma and thyroid carcinoma origins. A number of the NME1-dependent genes identified in these analyses are also shown to have prognostic value for survival in melanoma and breast cancer patients.
Materials and Methods
Cell lines and culture conditions. The WM1158 (metastasis-derived) melanoma cell line was a gift of Dr. Meenhard Herlyn (Wistar Institute, Philadelphia, PA, USA) and WRO82 cells (derived from a follicular thyroid carcinoma metastasis) were obtained from Dr. G. Juillard (University of California-Los Angeles, Los Angeles, CA, USA (16)). HEK-293 cells were obtained from the American Type Culture Collection. WM1158 cells were cultured at 5% CO2 in Tu2% media, composed of the following: MCDB:Leibovitz-15 medium (4:1, v/v; Sigma-Aldrich, St. Louis, MO, USA) supplemented with 2 mM CaCl2, 2.5 μg/ml insulin and 2% fetal bovine serum (Life Technologies, Grand Island, NY, USA). The other cell lines were maintained at 10% CO2 in Dulbecco’s Modified Eagle Medium supplemented with 10% fetal bovine serum, 1 mM non-essential amino acids, 100 U/ml penicillin and 100 μg/ml streptomycin.
Plasmid and viral vectors. A DNA fragment harboring full-length NME1 coding sequence and Kozak box with no 5′- or 3′-untranslated sequence was previously cloned into an expression plasmid (pCI-IRES-GFP) and designated pCI-H1(6). The key features of this plasmid are its CMV promoter, which is broadly expressed in most cell types, and an IRES (internal ribosome entry sequence) upstream of the marker gene GFP, which allows co-expression of NME1 and GFP from the same promoter. Using the restriction enzymes Bgl2 which cuts just upstream of the CMV promoter, and ClaI which cuts downstream of the GFP, the entire expression cassette was transferred into the corresponding site of pAdLink as described (17) to create pAdLink-NME1. For construction of adenovirus via recombination, HEK-293 cells (2×106 cells) expressing the adenoviral E1 gene were first seeded in 60-mm dishes. The cells were transfected 24 h later with 3.2 μg of ClaI-digested DNA isolated from wild type Ad5 (18), plus 4.8 μg of NheI-digested pAdLink-NME1. Following recombination, viral plaques expressing GFP were isolated and expanded in HEK-293 cells, and respective DNAs screened for the presence of NME1 by dot-blot analysis using a radiolabeled DNA probe generated by PCR from the NME1 cDNA sequence. Correct recombination of DNA fragments was verified by Southern blot using the same radiolabeled probe, followed by another round of plaque purification and Southern blotting. Potential contamination of the resulting virus, Ad5-NME1, by wild-type Ad5 was measured using PCR. A previously described Ad5-based vector also containing the IRES-GFP cassette plus a LacZ coding sequence (18, 19) was used as a negative control and is denoted herein as Ad5-clig. The Ad5-NME1 and Ad5-clig adenoviruses were purified by CsCl density gradient centrifugation (20) and eluted in 50 mM Hepes (pH 7.2), 0.9% NaCl, 0.5 mM MgCl2, 0.5 mM CaCl2 and 0.5% bovine serum albumin. Purified recombinant viruses were again screened for wild-type Ad5 adenovirus by PCR. Adenoviral titers were measured with the Adeno-X Rapid Titer Kit (Clontech; Mountain View, CA, USA) and were as follows: Ad5-clig, 4.6×1010 i.f.u/ml; Ad5-NME1, 2.9×1010 i.f.u/ml. Adenovirus stocks were stored in 5% glycerol in Dulbecco’s phosphate buffered saline at -80°C.
RNA isolation and microarray analysis. WM1158 cells were seeded at a density of 1.5×105 per 100-mm plastic dish and grown for 3 days to 75-80% confluence in Tu2%. Total cellular RNA was harvested (RNeasy Extraction Kit, Qiagen, Valencia, CA, USA) from five independent dishes for each cell line and virus treatment group. Purified RNA samples were diluted to ∼1 mg/ml with RNAse-free deionized water and transferred to the University of Kentucky Microarray Core Facility for cDNA synthesis, hybridization and scanning using their established protocols (http://www.research.uky.edu/microarray/AffymetrixGeneChip.html). Genome-wide expression profiling was conducted using U133A human genome arrays (Affymetrix, Santa Clara, CA, USA).
Gene expression values were calculated using Affymetrix® Expression Console™ Software version 1.1.2. Expression signals were subjected to background correction and scaled across all chips using quantile normalization. Resulting intensities were summarized and log2 transformed using the Robust Multi-Array Average (RMA) algorithm (21). Primary data are available via the NCBI Gene Expression Omnibus (GEO) (http://www.ncbi.nlm.nih.gov/geo/).
Kaplan-Meier analysis. Gene expression profiles of 31 primary and 52 metastatic melanoma biopsies published by Xu et al. (22) were obtained from Gene Expression Omnibus (GEO) under series accession number GSE8401. The expression profiles were imported into Expression Console v1.1.2 software (Affymetrix) and RMA was used to normalize the expression data across the samples. SigmaPlot 12 (Systat Software, San Jose, CA, USA) was used to generate Kaplan-Meier survival curves and perform log-rank tests to assess statistical significance between “high” and “low” expression of individual genes in the metastatic melanoma samples. Samples were categorized as “high” or “low” expressers if the log2 expression of gene was above or below, respectively, the median of all samples for that gene. Statistically significant differences in normalized log2 expression of individual genes in GSE8401 and a second human melanoma GEOdataset by Kabbarah et al (accession number GSE46517) (23) were analyzed by one-way ANOVA followed by pairwise multiple comparison testing (Holm-Sidak method, SigmaPlot 12). The GSE46517 dataset consisted of 9 nevi, 31 primary melanoma, and 73 metastatic melanoma biopsies.
Kaplan-Meier survival analyses in breast cancer were conducted using BreastMark, a newly-described breast cancer database and algorithm (24). The system contains integrated gene expression and survival data from 26 datasets on 12 distinct microarray platforms from a total of 4,738 patient samples, distributed across the four major breast cancer subtypes of luminal A, luminal B, Her2-positive and basal. Briefly, the BreastMark tool utilizes log-rank tests to measure differences in survival curves and Cox regression analyses to calculate hazard ratios via a web-based server (http://glados.ucd.ie/BreastMark/). The BreastMark tool conducts all calculations in the R statistical environment.
Results
Construction of an adenoviral vector for forced NME1 expression in metastatic cell lines of melanoma and thyroid carcinoma origin. To identify genes dysregulated by loss of NME1 in metastatic cancers, gene expression profiling was conducted in the absence or presence of forced NME1 expression. The study was conducted in two human metastatic cell lines obtained from distinct cancers, melanoma and thyroid carcinoma, to identify NME1-responsive genes unique to the respective cancers, as well genes regulated across both. It was postulated that genes regulated by NME1 across these two diverse metastatic cancer types would be enriched for relevance to metastasis-suppressor function. The melanoma cell line, WM1158, was derived from a lymph node metastasis in a patient with a Stage II, superficial spreading melanoma lesion (http://ccr.coriell.org/Sections/Collections/WISTAR/CellLines.aspx?PgId=572&coll=WC). This metastatic line was suitable for measuring effects of forced NME1 expression, as we showed previously that it exhibits markedly reduced NME1 and NME2 expression (6). WRO82 was originally obtained from a metastatic follicular thyroid carcinoma lesion, and expresses almost undetectable levels of NME1 and NME2.
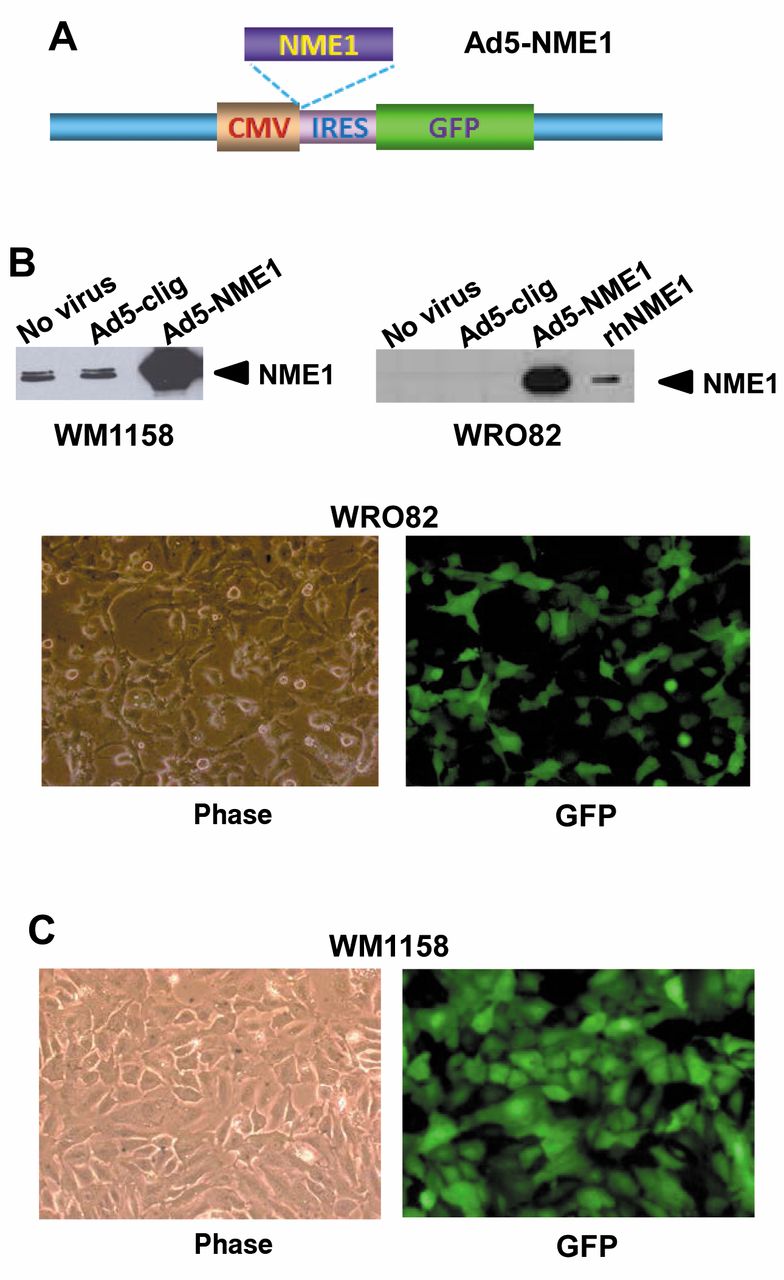
Prior efforts at forced NME1 expression via stable transfection in metastatic melanoma cell lines have provided only modest (2- to 3-fold) increases at best in our laboratory, probably due to proteolytic degradation of NME1 in the lysosome, as we recently described (25). To maximize effects of NME1 expression on overall gene expression profiles in these studies, we investigated the utility of an adenoviral (Ad5-based) expression vector, constructed with an NME1 cDNA under control of the cytomegalovirus promoter/enhancer (Ad5-NME1) (Figure 1A). An internal ribosome entry sequence (IRES) and green fluorescent protein cDNA generates a bicistronic mRNA for co-translation of NME1 and GFP proteins. Infection with Ad5-NME1 provided robust expression of NME1 protein in both WM1158 and WRO82 cells, as indicated by immunoblot analysis (Figure 1B). GFP fluorescence was observed microscopically in >90% of infected cells in both lines, consistent with attainment of NME1 overexpression in a similar proportion of the cells (Figure 1C).
Profiling of NME1-regulated gene expression in WM1158 and WRO82 cells. For RNA expression analysis, WM1158 and WRO82 cells were infected with the control adenovirus Ad5-clig, Ad-NME1, or were left uninfected, followed 48h later by harvesting of total cellular RNA (see Materials and Methods). RNA samples were analyzed using the U133A human microarray chip (Affymetrix), which analyzes expression for approximately 18,400 transcripts and variants. Genes were identified as NME1-regulated based on the following criteria. Signal intensities for the three experimental groups were compared pairwise using Welch two sample t-tests. Resulting p-values were adjusted for multiple comparisons using the correction method of Benjamini and Hochberg (26) and are listed as FDR p-values herein. Probe sets showing significant differences (unadjusted p<0.05) between the uninfected and Ad5-clig control groups were filtered out as non-specific, adenovirus infection-induced effects. NME1-induced expression changes were then identified as probe sets showing significant and consistent alterations as compared to both uninfected and Ad5-clig-infected cells at the adjusted p-values indicated.
Forced expression of NME1 in WM1158 cells resulted in up-regulation of 390 genes and down-regulation of 338 genes, as determined using a false discovery rate (FDR) of 0.1 (Figure 2). Ontology analysis using Ingenuity® pathway analysis software identified eight functional categories altered significantly by NME1 expression (Table I). The most pronounced effect was on the “cell death and survival” category which contained 276 genes related to apoptosis, cell death and viability functions. Other strongly represented categories included “small molecule biochemistry” (104 genes), “DNA replication, recombination and repair” (49 genes), “cell signaling” (49 genes), and “nucleic acid metabolism” (37 genes). NME1-mediated regulation of nucleotide metabolism genes is noteworthy in light of the nucleoside diphosphate kinase activity of NME1, whose activity would be expected to impact regulation of other genes in the category. Furthermore, a significant NME1-induced enrichment for members of the CDC42 signaling cascade (ARPC5L, CDC42, CDC42EP2, FNBP1L, HLA-DOA, HLA-F, HLA-G, ITGB1, JUN, MYL7, MYL10, MYL12A and RASA1) was identified (Fisher’s exact test p<0.014). Using a more stringent FDR (≤0.05), 13 down-regulated and 19 up-regulated genes were identified (Table II). Of particular note in this list were strong inductions of such metastasis-relevant genes as matrix metallopeptidase 1 (MMP1), angiopoietin 2 (ANGPT2), interleukins 8 and 11 (IL8 and IL11), and induction of two genes associated with cellular apoptosis, GADD45A and programmed cell death 11 (PDCD11).

The adenoviral expression vector Ad5-NME1 provides strong NME1 expression in WM1158 and WRO82 cell lines. A: Salient features of Ad5-NME1 are shown: CMV, cytomegalovirus promoter/enhancer; NME1, NME1 cDNA; IRES, internal ribosome entry site/sequence; GFP, green fluorescence protein; LacZ, β-galactosidase. B: Immunoblot analysis of NME expression at 48h post-infection with Ad5-NME1. rhNME1, recombinant human NME1 (1 μg) expressed in E. coli and purified as described (5). C: Representative phase and fluorescence microscopic images from WM1158 and WRO82 cells infected with pAd5-NME1.

Supervised hierarchical clustering of genes differentially expressed in WM1158 and WRO82 cells in response to forced NME1 expression. Expression differences were identified by comparing cells with forced overexpression of NME1 using the Ad5-NME1 adenovirus to two control treatments (Ad5-clig control virus and untransduced) using t-test within the cell lines. A: Gene expression in WM1158 cells. B: Gene expression in WRO82 cells. False discovery rates were used for multiple test correction, with the most significant probesets (FDR p-value<0.1) displayed. The heatmaps depict relative expression levels for each significant probe set (rows) among each of triplicate samples (columns), based on the number of standard deviations from the mean. Lower relative expression levels are indicated in blue and higher levels in red. For WM1158, 787 probe sets corresponding to 733 genes were identified, while 267 probe sets (253 genes) were found in WRO82 cells. C: Venn diagram summary of NME1-regulated genes (FDR≤0.1) in WM1158 and WRO82 cells. The total number of NME1-regulated genes is given for each cell line, the number of up- and down-regulated genes (represented by up and down arrows) within them, and the intersection of twelve genes up-regulated in both.
Enrichment of gene groups regulated by forced NME1 expression in metastatic WM1158 melanoma cells.
Genes regulated by forced NME1 expression in the metastatic melanoma cell line, WM1158 (FDR < 0.05).
A substantial number of genes were also regulated by NME1 in WRO82 cells (FDR <0.1), with 185 up-regulated and 68 down-regulated transcripts identified (Figure 2). NME1 expression also exerted an impact on multiple functional gene categories in WRO82 cells (Table III), with the strongest effects seen on “cell-mediated immune response” (5 genes), “cellular development” (32 genes), “cellular function and maintenance” (23 genes), hematological system development and function” (43 genes), and “hematopoiesis” (26 genes). At the more stringent level (FDR <0.05), a total of 6 down-regulated and 14 up-regulated genes were identified (Table IV). Effects of potential relevance to metastasis were the down-regulation of BCL2L1 and PALB2, and the up-regulation of CYP3A43 (3.0-fold), SERPINB9 (2.9-fold), KIF5A, PPM1E, and CSF2RB. KIF5A and PPM1E were of particular interest for their roles in microtubule-based movement (27) and regulation of the nonreceptor tyrosine kinase PAK1 (28), respectively.
Having identified numerous NME1-regulated genes in two cell lines representing diverse forms of metastatic cancer, an effort was also made to identify genes that were similarly regulated across both lines. Using an FDR<0.1, a total of nine such genes were identified, six of which were up-regulated and three down-regulated (Table V). Interestingly, three (PPM1E, ZYX, PFN1) of those genes are participants in the highly metastasis-relevant processes of stress fiber formation and focal adhesion. On the whole, each of the eight genes possessed metastasis-relevant activities, such as chemotaxis (CCR1), epithelial-mesenchymal signaling (WNT6), differentiation and morphogenesis (TBX4, ZFP36L2), and G protein modulation (GPR52 and PFN1). Taken together, the genes coregulated by NME1 across metastatic cell lines from these distinct cancers appear to be enriched for functions of probable relevance to metastatic potential.
Enrichment of gene groups regulated by forced NME1 expression in metastatic WRO82 thyroid carcinoma cells.
Genes regulated by forced NME1 expression in the metastatic thyroid carcinoma cell line, WRO82 (FDR < 0.05).
Genes regulated similarly by forced NME1 expression in both WM1158 and WRO82 cells (FDR<0.1).
Summary of genes regulated by NME1 in both WM1158 and WRO82 cells that predict survival similarly in melanoma and at least one subtype of breast cancer. Blue checks represent genes associated with longer patient survival, and those in red are negatively associated with survival.
NME1-regulated genes are strong predictors of survival in melanoma. As a metastasis suppressor in cell culture and in vivo settings, NME1 may regulate the expression of genes that determine metastatic potential and overall survival in human cancer patients. To determine whether the NME1-regulated genes identified in the microarray analyses were of predictive value, we conducted correlational analysis between expression of these genes and indices of patient metastasis and survival in existing RNA expression databases from melanoma patients. Expression of NME1 RNA in human melanoma samples was not well-correlated with many of the NME1-regulated genes identified in the WM1158 and WRO82 cell lines. This was expected, as NME1 down-regulation is exerted at the level of protein expression via lysosomal degradation (25) rather than RNA expression in the majority of cancer cell lines we have analyzed to date. Nevertheless, it was considered of potential value to analyze the predictive power of the NME1-regulated genes, with the premise that many are likely under control of NME1 protein expression in melanoma tumors. Analyses could not be conducted in thyroid carcinoma patients due to a lack of robust, publicly available RNA expression datasets.
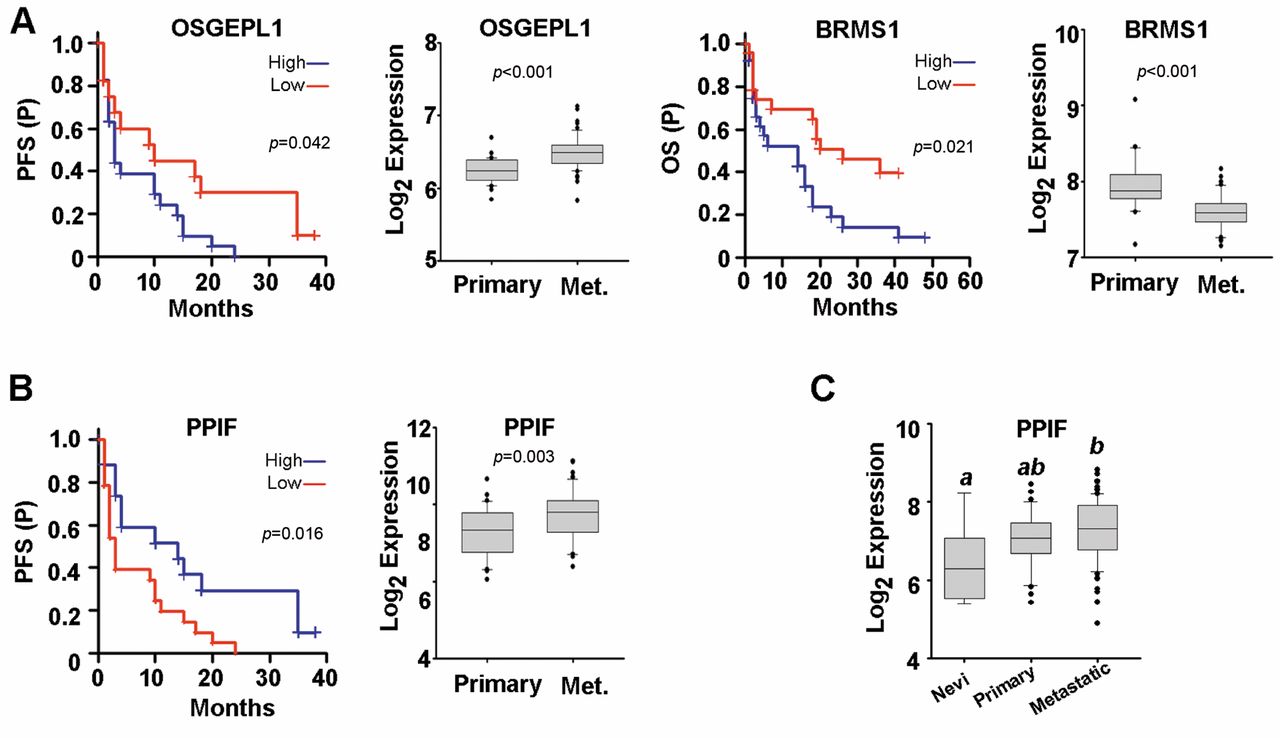
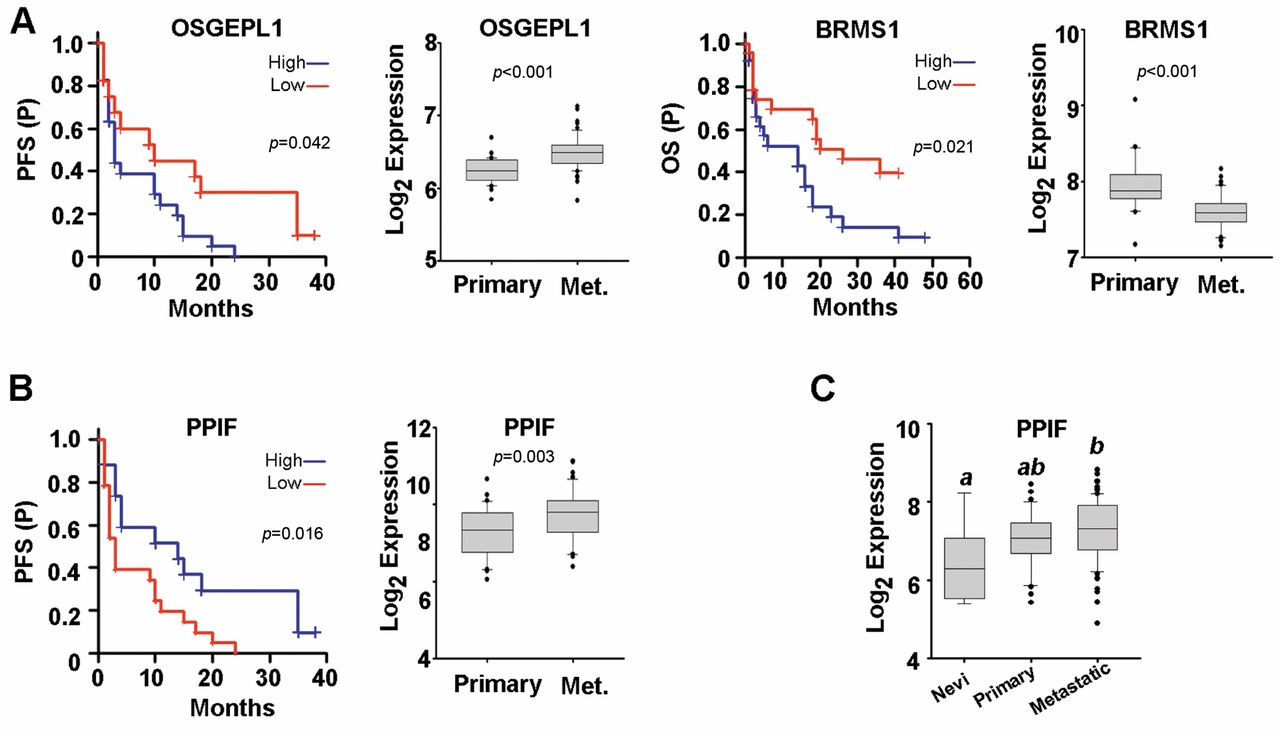
Genes regulated by NME1 in WM1158 cells are predictive of survival and RNA expression in metastatic melanoma patients. A: Shown are Kaplan-Meier plots for NME1-regulated genes that displayed significant effects on both patient survival (left panels: progression-free, PFS; overall, OS) and differences in RNA expression in primary versus metastatic melanoma biopsies (right panels). Both analyses employed the GEO dataset GSE8401 (22). B: Kaplan-Meier survival plots for the PPIF gene in metastatic melanoma samples (left) and RNA expression in primary metastatic versus primary melanoma biopsies from GSE8401 (right). C: Expression of the PPIF RNA in the human melanoma progression GEO dataset GSE46517. Groups not sharing a common superscript are statistically significantly different (p≤0.05 by ANOVA, Holm-Sidak post-hoc testing).
Of the top twenty genes regulated by NME1 in the melanoma cell line WM1158, three were found to have a significant predictive value in either progression-free survival (PFS) or overall survival (OS) of metastatic melanoma patients and also display significantly different RNA expression in primary versus metastatic melanoma samples in the GEO dataset, GSE8401 (Figure 3). Consistent with being repressed by the metastasis suppressor NME1 (Table II), expression of OSGEPL1 and BRMS1 were both associated with shorter PFS and OS, respectively (Figure 3A). Expression of the NME1-induced gene, PPIF, was correlated with longer progression-free survival of metastatic melanoma patients (Figure 3B, upper left). Interestingly, PPIF expression was significantly higher in metastatic samples versus primary melanomas or benign nevi in both the GSE8401 dataset (Figure 3B) and another (GSE46517) (Figure 3C), suggesting complexity in its regulation by NME1 as well as potential survival value in metastatic melanoma.
Interestingly, a number of genes regulated by NME1 in the metastatic thyroid carcinoma cell line WRO82 also exhibited significant predictive value for survival in metastatic melanomas (Figure 4A-C). Three genes, SERPINB9, CSF2RB and SLC4A4, were positively correlated with both PFS (Figure 4B) and OS (Figure 4C). In order to identify NME1-regulated genes with the most potential for clinical application, we filtered for genes that displayed both 1) significant predictive value for melanoma survival and 2) differential expression in metastatic melanoma relative to primary lesions or benign nevi in two independent melanoma datasets. Two genes, SPARCL1 and TEAD4, met those selection criteria. Consistent with metastasis suppressor function and its induction by NME1, SPARCL1 was a strong predictor of increased OS and its expression was significantly decreased in more advanced melanoma samples (Figures 4D and 4E). Conversely, TEAD4, which was repressed by NME1 (Table IV), was inversely associated with OS and also significantly increased in metastatic melanomas compared to primary melanomas in dataset GSE8401 (Figure 4D) and to benign nevi in dataset GSE46517 (Figure 4E).
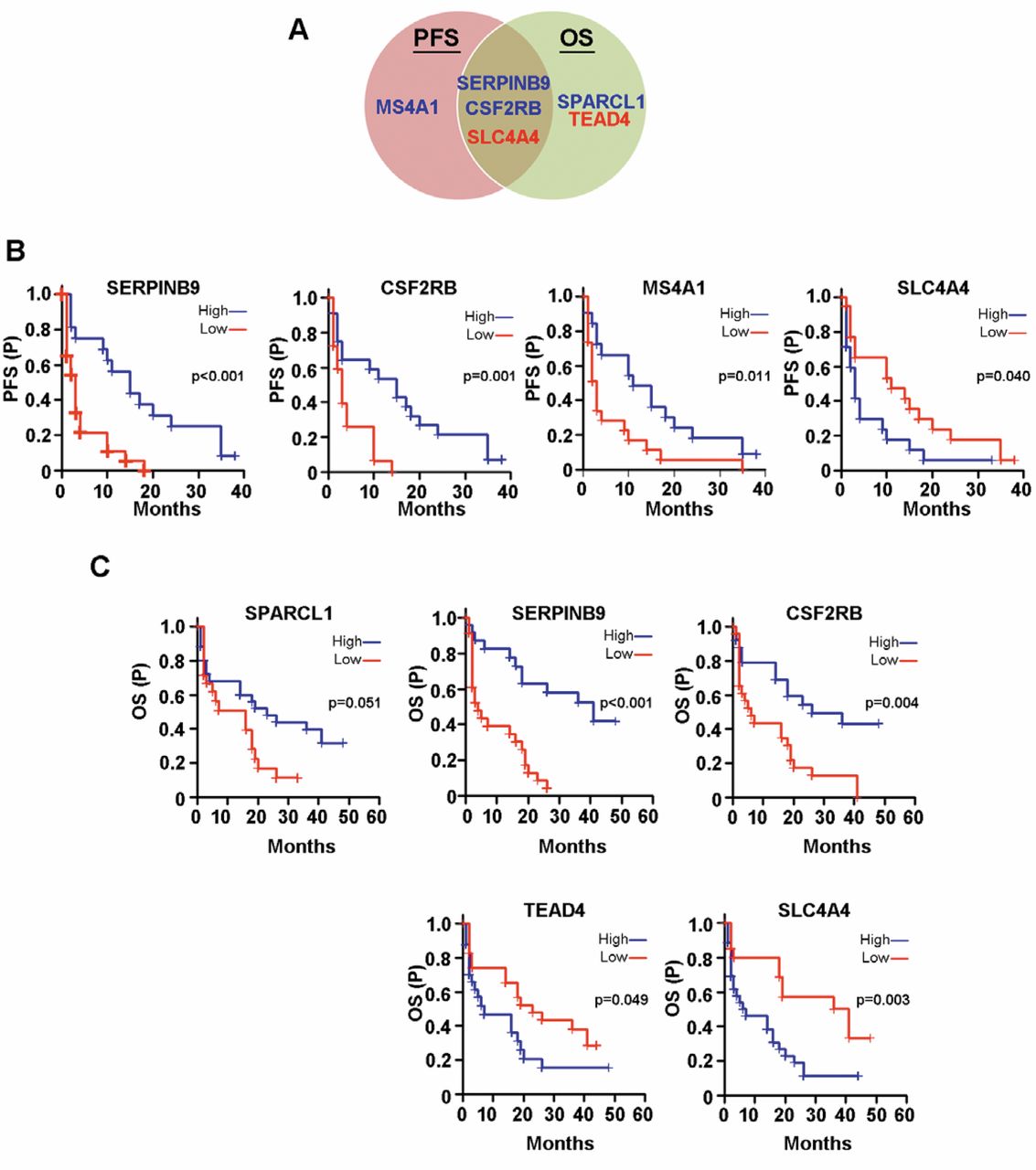

Summary of Kaplan-Meier survival plots in metastatic melanoma for genes regulated by NME1 in WR082 cells. A: Venn diagram summarizing NME-regulated genes predictive of progression-free survival (PFS) and/or overall survival (OS) in melanoma patients. B: NME1-regulated genes that were predictive of PFS and C: OS in metastatic melanoma patients from the GSE8401 GEOdataset. D: Genes with significantly different RNA expression in melanoma dataset GSE8401. E: Genes with significantly different RNA expression in melanoma dataset GSE46517. Groups not sharing a common superscript are statistically significantly different (p≤0.05 by ANOVA, Holm-Sidak post-hoc testing).
The list of genes coregulated by NME1 across both the metastatic WM1158 and WRO82 cell were next tested for their potential association with survival in melanoma patients. Of the nine genes found to be regulated by NME1, two genes, CCR1 and ZFP36L2, displayed significant predictive value in patient survival times. Expression of each gene was associated with prolonged PFS (Figure 5A), while CCR1 was also predictive of improved OS (Figure 5B). This commonality is intriguing as CCR1 was induced by NME1 while ZFP36L2 was repressed by NME1 (Table V). In addition, CCR1 was significantly increased in more advanced lesions while ZFP36L2 expression was decreased in advanced melanomas (Figure 5C). By combining all of the individual lists of NME1-regulated genes, we have identified seven genes with potential diagnostic value in terms of predicting melanoma patient survival as well as differential expression in more advanced samples.
A number of genes regulated by NME1 in melanoma and thyroid carcinoma cell lines are predictors of survival in breast cancer. In light of the well-documented metastasis suppressor function of NME1 in human breast cancer (45), we also examined the utility of NME1-regulated genes identified in WM1158 and WRO82 cells as prognostic indicators of breast cancer survival. Analyses were conducted using the breast cancer database and algorithm, BreastMark (24), which contains integrated gene expression and survival data from 26 datasets obtained with 12 distinct microarray platforms on a total of 4,738 patient samples, distributed across the four major breast cancer subtypes of luminal A, luminal B, Her2-positive and basal (http://glados.ucd.ie/BreastMark/). A number of the NME1-regulated genes identified in WM1158 melanoma cells provided statistically significant predictive value in the luminal A subtype (Figure 6A). All expressions were inversely correlated with survival, including MMP1 (distant disease-free survival, or DDFS, p<0.048), FKBP1 (DDFS; p<0.026), HSP90AA1 (DDFS, 0.013; overall survival or OS, p<0.006), and BRMS1 (DDFS, p<0.021; OS, p<0.018). Moreover, the combination of HSP90AA1, MMP1 and FKBP1 expression exhibited a highly cooperative association with poor DDFS (p<0.004, hazard ratio or HR of 1.9) and OS (p<0.006, HR of 1.9). In the setting of the luminal B subtype of breast cancer, expression of six NME1-regulated genes identified in WM1158 cells was associated with patient survival (Figure 6B), with two showing a correlation with improved DDFS (PPIF and PDCD11) and four inversely correlated with OS (PQBP1, CNPY2, ANGPT2 and IL8). Expression of the genes AKAP12 (positive) as well as the previously cited genes ANGPT2 and MMP1 (both inverse) were associated with OS in the Her2 subtype (Figure 6C), while expression of FKBP3 (inverse), PPIF and POLR2G (both positive) were correlated with DDFS in the basal subtype (Figure 6D).
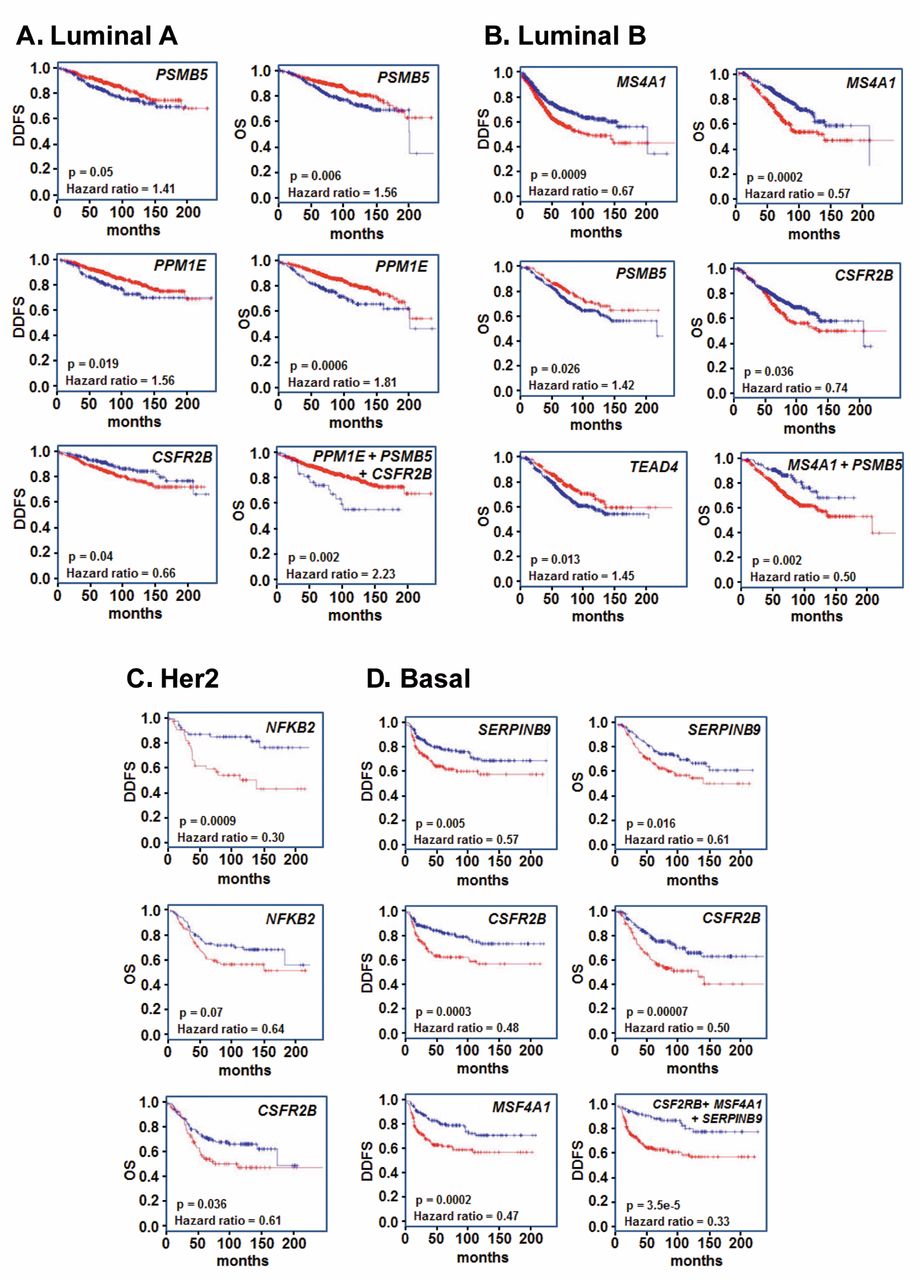
A number of the genes identified as NME1-regulated in the metastatic thyroid carcinoma cell line WRO82 also had significant predictive value for survival across the four breast cancer subtypes. Expression of two genes, PSMB5 and PPM1E, was inversely correlated with both DDFS and OS in the luminal A subtype (Figure 7A). In addition, expression of the colony-stimulating factor receptor isoform CSFR2B was positively correlated with DDFS (p<0.04) in luminal A patients. Moreover, the combination of PPM1E, PSMB5 and CSFR2B exhibited significant synergy in predicting OS for this subtype (p<0.002, HR=2.23). In luminal B patients (Figure 7B), expression of MS4A1 exhibited a strong inverse correlation with both DDFS (p<0.0009) and OS (p<0.0004). As also seen in luminal A patients, expression of PSMB5 (p<0.026, positive correlation) and CSFR2B (p<0.036, inverse correlation) had predictive value for OS in the luminal B subtype. Expression of another gene, TEAD4, was also positively correlated with OS (p<0.013). In Her2-positive patients (Figure 7C), NFKB2 expression was correlated strongly with DDFS (p<0.0009) and approached significance for OS (p<0.07), and CSFR2B was again correlated with DDFS (p<0.036). In the basal subtype (Figure 7D), expression of SERPINB9 was a strong predictor for both DDFS (p<0.005) and OS (p<0.016), as again were CSFR2B (p<0.0003 and 0.00007 for DDFS and OS, respectively) and MSF4A1 (p<0.0002, for DDFS only). In the strongest example of cooperativity observed in the overall survival analyses, expression levels of CSFR2B, MSF4A1 and SERPINB9 together were highly predictive of DDFS in the basal subtype (p<3.5e-5, HR=0.33).
A total of seven genes identified in WM1158 and WRO82 cells as NME1-regulated were shown to have predictive value in melanoma and at least one subtype of breast cancer (Table VI). As might be expected, each gene had the same positive or negative effect on patient survival across the different cancers (i.e. genes associated with longer/shorter survival in melanoma were also associated with longer/shorter survival in one or more breast cancer subtypes). The most robust of these genes was CSF2RB, expression of which was associated with longer PFS in melanoma and the breast cancer subtypes LumA and basal, as well as longer OS in melanoma and LumB, Her2 and basal breast cancers. The other NME1-regulated genes exhibiting associations with survival to varying degrees across these cancers consisted of MS4A1, PPIF, SERPINB9, ZFP36L2, BRMS1 and TEAD4. Taken together, these analyses of melanoma and breast cancer datasets demonstrate that NME1-regulated genes identified in the two metastatic cell lines of melanoma (WM1158) and thyroid carcinoma (WRO82) origin were enriched in predictive value for survival in melanoma and breast cancer patients, possibly due to their impact on the metastatic phenotype.
Discussion
While considerable evidence suggests that the metastasis suppressor function of NME1 is mediated directly via inhibitory physical interactions with motility-driving proteins, other studies indicate it regulates expression of motility and metastasis-relevant genes as well (12, 29). This study was conducted to address a current gap in knowledge regarding the impact of NME1 on global profiles of gene expression in two cancers, melanoma and thyroid carcinoma. To this end, a novel adenovirus expression vector was constructed that successfully provided robust and efficient expression of NME1 in both melanoma (WM1158) and thyroid carcinoma (WRO82) cell lines. Forced NME1 expression resulted in broad impacts on gene expression profiles in both cell lines, with ontology analysis revealing enrichment of a number of gene groups (Tables I and III). One gene group of interest identified in the WM1158 melanoma cell line was DNA replication, recombination and repair (49 genes), which may well relate to our recent observations that NME1 contributes to the repair of UV-induced DNA damage in yeast and mammalian cells (4, 30). Other gene groups identified in the ontology analysis, such as cell death and survival, cellular development/differentiation, and cell signaling strongly suggest NME1 regulates expression of genes capable of impacting metastatic potential in both cancers. Of interest was the significant regulation of CDC42 and a number of its downstream effectors in WM1158 melanoma cells. While NME1 has been shown to induce motility suppression in MDA-MB-435 cells through a physical interaction with CDC42 to modulate downstream signaling (30), our data suggest an alternative pathway of regulating expression of CDC42 pathway genes in the context of melanoma.

Genes regulated by NME1 in both WM1158 and WR082 cells are predictive of survival in metastatic melanoma. Shown are NME1-regulated genes that displayed significant effects on A: progression-free survival (PFS) or B: overall survival (OS) of metastatic melanoma patients from the GSE8401 dataset. C: RNA expression of CCR1 and ZFP36L2 in the human melanoma progression GEOdataset GSE46517. Groups not sharing a common superscript are statistically significantly different (p≤0.05 by ANOVA, Holm-Sidak post-hoc testing).
Interestingly, the majority of NME1-down-regulated genes identified in WM1158 cells with known interactions with cancer (4/5) had the expected anti-motility/metastatic functions. For example, the NME1-down-regulated gene CNPY2 potentiates FGF-induced cell motility in glioma and neurite outgrowth in neuroblastoma cells (31). In fact, we and others observe an association of neurite-like outgrowth with more aggressive phenotypes in melanoma and melanoma cell lines (32). Additional examples were PPP1CA, a carcinoma oncogene (33) and risk factor for recurrence of bladder carcinoma (34), and ELL3/SERINC4 which has been documented to induce the cancer stem cell properties of breast carcinoma cells. While our observation of NME1-down-regulation of the well-characterized melanoma metastasis suppressor BRMS1 (35) is at first glance counter-intuitive, additional complexity is indicated by recent observations that its subcellular localization may be as important as expression level (36). In contrast, a majority of NME1-up-regulated genes (5/6), such as MMP1 (37), ANGPT2 (38), IL8 (39) and IL8 (40), and HSP90AA1 (11, 41) were associated with functions that could be metastasis-driving. One potential factor may have been the robust NME1 overexpression achieved in our studies with the Ad5clig-based adenoviral vector, which was much higher than achieved with plasmid-based stable transfection in our laboratory. While we have observed the expected low NME expression pattern in many metastatic cell lines of melanoma and other cancer origins, we have also observed marked NME overexpression in other melanoma cell lines and patient samples, suggesting that strong NME overexpression may have deleterious impacts on gene expression and cancer phenotypes as well. Alternatively, the roles played by these genes in metastasis are not fully understood, and theoretically could be suppressive in certain cellular and physiological conditions.
Three genes regulated by NME1 in WRO82 cells possess activities in cancer consistent with being effectors of suppressor activity. In fact, the NME1-up-regulated gene SPARCL1 has been reported to exhibit bona fide metastasis suppressor activity in carcinomas of the prostate (42), breast (43) and colon (44). Another NME1-up-regulated gene PPM1E, plays a key role in PAK1-driven breakdown of cytoskeletal stress fibers (28) and is found within a susceptibility locus for testicular germ cell tumor (45), the latter suggesting a suppressor function. Conversely, the NME1-down-regulated gene TEAD4 is reported to be a tumor driver and mediator of chemoresistance in hepatocellular carcinoma (46), and is associated with poor prognosis in ovarian and gastric carcinoma (47).

Genes regulated by NME1 in WM1158 cells are predictive of survival across multiple breast cancer subtypes, as identified using the BreastMark RNA expression database and algorithm (24). Kaplan-Meier plots shown in red represent survival data for patients in whom expression of the queried gene is significantly below the median, while those in blue represent expression above the median. DDFS, distant disease-free survival; OS, overall survival.

Genes regulated by NME1 in WRO82 cells are predictive of survival across multiple breast cancer subtypes, as identified using the BreastMark RNA expression database and algorithm (24). Kaplan-Meier plots shown in red represent survival data for patients in whom expression of the queried gene is significantly below the median, while those in blue represent expression above the median. DDFS, distant disease-free survival; OS, overall survival.
A substantial number of the genes identified as NME1-regulated in WM1158 and WRO82 cells were of predictive value for survival in melanoma and the major subtypes of breast cancer, another NME1-dependent cancer. Moreover, many of those possessed significant concordance between the directionality of regulation by NME1, their canonical functions in cancer, and their impact on survival. Perhaps the strongest concordance was observed with the NME1-up-regulated metastasis suppressor SPARCL1, whose expression was positively correlated with survival in both melanoma datasets (GSE8401 and GSE46517), as well as with overall survival (OS) in LumA (also disease-free survival, DFS), LumB and Her2 subtypes of breast cancer. As predicted, low expression for many NME1-down-regulated genes in WM1158 cells (i.e. CNPY2, OSGEPL1, PQBP1, FKBP3, POLR2G, BRMS1, ATP5D and ELL3SERINC4) was associated with better survival in melanoma and/or breast cancer. For example, low CNPY2 expression was associated with improved DFS and OS in LumB breast cancers, while low expression of FKBP3 was associated with better survival across all four breast cancer subtypes (LumA, DDFS; LumB, DFS; Her2, DFS and OS; basal, DDFS and OS). Interestingly, expression of the NME1-down-regulated gene BRMS1 was reduced in metastatic melanoma, consistent with its metastasis suppressor function in melanoma but, surprisingly, was inversely correlated with OS in melanoma, as well as survival in the LumA (DFS, DDFS and OS) and OS in LumB subtypes of breast cancer. Thus, the down-regulatory effect of NME1 on BRMS1 expression, while superficially discordant with prediction, was actually concordant with the association between BRMS1 expression and poorer survival in melanoma and breast cancer patients. As the metastasis suppressor function and expression profiles associated with BRMS1 have been established convincingly (35), our data suggest further complexity, possibly due to heterogeneity within these cancer populations.
A recent paper has described NME1 as a factor whose function is in “binding up the bad guys” in cancer (48). Our study as well as previous analyses of gene expression in breast cancer cell lines, indicates NME1 also regulates expression of RNAs that encode metastasis drivers and suppressors in melanoma. Clearly much remains to be done to further elucidate the mechanisms underlying this regulation, which may reside not only in transcriptional regulation but all other events in the regulatory cascade including RNA splicing, export and stability. The NME1-regulated genes uncovered in this analysis highlights potential candidates for diagnosis and prognosis in melanoma and other NME1-dependent cancers. Moreover, they represent candidate effectors of its metastasis suppressor function that could be exploited as targets in therapy of such cancers.
Acknowledgments
This work was supported by the United States National Institutes of Health, National Cancer Institute grants CA83237 and CA159871 (D.M. Kaetzel), and training grant T32CA15474 from the National Cancer Institute (M.K. Leonard).
Footnotes
-
↵† Deceased.
- Received April 28, 2014.
- Revision received June 18, 2014.
- Accepted June 23, 2014.
- Copyright© 2014, International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- A noncanonical microRNA derived from the snaR-A noncoding RNA targets a metastasis inhibitor
- Structure Learning for Hierarchical Regulatory Networks
- A non-canonical microRNA derived from the snaR-A non-coding RNA targets a metastasis inhibitor
- NME1 Drives Expansion of Melanoma Cells with Enhanced Tumor Growth and Metastatic Properties