


Abstract
The relationship between supplemental vitamins and various types of cancer has been the focus of recent investigation, and supplemental vitamins have been reported to modulate cancer rates. A significant association has been demonstrated between cancer and low levels of thiamine in the serum. Genetic studies have helped identify a number of factors that link thiamine to cancer, including the solute carrier transporter (SLC19) gene, transketolase, transcription factor p53, poly(ADP-ribose) polymerase-1 gene, and the reduced form of nicotinamide adenine dinucleotide phosphate. Thiamine supplementation may contribute to a high rate of tumor cell survival, proliferation and chemotherapy resistance. Thiamine has also been implicated in cancer through its effects on matrix metalloproteinases, prostaglandins, cyclooxygenase-2, reactive oxygen species, and nitric oxide synthase. However, some studies have suggested that thiamine may exhibit some antitumor effects. The role of thiamine in cancer is controversial. However, thiamine deficiency may occur in patients with cancer and cause serious disorders, including Wernicke’s encephalopathy, that require parenteral thiamine supplementation. A very high dose of thiamine produces a growth-inhibitory effect in cancer. Therefore, further investigations of thiamine in cancer are needed to clarify this relationship.
15-PGDH: 15-Hydroxyprostaglandin dehydrogenase; ATTP: adenosine thiamine triphosphate; CNS: central nervous system; DKA: diabetic ketoacidosis; ECM: extracellular matrix; COX: cyclooxygenase; DM: diabetes mellitus; HCC: hepatocellular carcinoma; KTS: 3-ethoxy-2-oxobutyraldehyde bis(thiosemicarbazone); iNOS: inducible NOS; MMP: matrix metalloproteinase; NADPH: the reduced form of nicotinamide adenine dinucleotide phosphate; NO: nitric oxide; NOS: nitric oxide synthase; NOX: the NADPH oxidase; LPS: lipopolysaccharide; PARP-1: Poly(ADP-ribose) polymerase-1; PDH: pyruvate dehydrogenase; PG: prostaglandins; RDA: recommended dietary allowance; ROS: reactive oxygen species; SLC: solute carrier transporter; SOD: superoxide dismutase; TD: thiamine-deficient; TDP: thiamine diphosphate; ThTk: thiamine transporter; TKTL-1: transketolase-like 1; TPP: thiamine pyrophosphate; TIMP-1: tissue inhibitor of metalloproteinase-1.
The relationship of supplemental vitamins and various types of cancer has been the focus of recent investigations, and supplemental vitamins have been reported to modulate cancer rates. In a New Hampshire study, higher total intakes of carotenoids, vitamin D, thiamine, niacin, and vitamin E were inversely related to bladder cancer risk among older individuals (1). Higher intake of vitamin C, beta-carotene, thiamine, and nicotinic acid was also associated with a decrease in gastric cancer risk in a case control study (2). In Hawaii, riboflavin and thiamine levels from food sources exhibited an inverse relationship with incidence of high-grade squamous intraepithelial lesions of the cervix (3). In Poland, higher consumption of alcohol, when combined with a low intake of retinol, thiamine or antioxidant micronutrients, increased the risk of colorectal cancer and established a cancer-preventive role for retinol and thiamine (4). High dietary intake of vitamin C and thiamine has been reported to reduce the risk of prostate cancer in China (5). In another study, Hargreaves et al. (6) suggested that low intake of thiamine, riboflavin, vitamins A and C, and iron may contribute to cancer risk in African-Americans. When monitoring micronutrient levels in gastrointestinal cancer, plasma vitamin C and red blood cell thiamine levels were significantly reduced in weight-losing patients with cancer compared with their weight-stable counterparts, and weight-losing patients exhibited lower vitamin C and thiamine intake (7). Furthermore, patients with acute leukemia exhibited low thiamine content in leukocytes and blood plasma (8). The thiamine pyrophosphate (TPP) stimulating effect and urinary excretion of thiamine were high in patients with breast and bronchial carcinomas, suggesting that these patients may be at risk for thiamine deficiency (9). Transketolase reactions play a crucial role in the use of glucose carbons in tumor cell synthesis of nucleic acid ribose. Experimentally, a dramatic decrease in tumor cell proliferation was observed in several in vitro and in vivo tumor models after administering the transketolase inhibitor oxythiamine (10, 11). Supplementation with excess thiamine and pantothenic acid in tumor-bearing rats reduced growth restriction and enhanced the antitumor activity of 3-ethoxy-2-oxobutyraldehyde bis(thiosemicarbazone) (KTS). Similarly, the addition of thiamine protected the KTS-treated rats from weight loss in a dose-related manner (12). These findings suggest that thiamine may be associated with cancer. Therefore, we reviewed the role of thiamine in cancer.
The Genomic Factors Associated with Thiamine in Cancer
The solute carrier (SLC) transport protein. The SLC19 gene is a family of three transporter proteins with significant structural similarities, but each protein binds substrates with different structures and ionic charges. The SLC19 gene family plays an important role in the transport and homeostasis of folate and thiamine in the body (13). SLC19A2 and SLC19A3 mediate the transport of thiamine (14, 15). SLC19A3 RNA levels have been reported to be down-regulated in breast and lung cancer compared with adjacent normal tissues (16, 17). The SLC19A3 gene is regulated by exogenous thiamine in a manner similar to the effect of SLC19A3 overexpression (17). SLC19A3 expression is down-regulated in gastric cancer cell lines (71%) and restored after pharmaceutical demethylation. Exogenous SLC19A3 expression causes growth inhibition of gastric cancer cells (17). SLC19A3 gene expression is down-regulated in breast cancer and associated with resistance to apoptosis in these tumors (18). Its mRNA expression is also down-regulated by DNA methylation, and by histone deacetylation in colon cancer cells (19). However, hypoxia induces the up-regulation and function of the SLC19A3 gene in a breast cancer cell line (20). Aberrant SLC19A3 has been suggested as a novel biomarker for breast and gastric cancer diagnosis (21). SLC19A2 has also been shown to transport mono- and pyrophosphate derivatives of thiamine (22, 23). Polymorphisms of SLC19A1 are reported in many types of cancers, such as osteosarcoma (24), colorectal cancer (25), non-small cell lung cancer (26), bladder cancer (27), prostate cancer (28), diffuse large B-cell lymphoma (29), pediatric acute lymphoblastic leukemia (30), head and neck cancer (31), central nervous system (CNS) tumors in Thai children (32), and human ovarian carcinoma cells (33).
Transketolase. Thiamine acts as a co-enzyme for transketolase, pyruvate dehydrogenase (PDH), and α-ketoglutarate dehydrogenase complexes, which contain enzymes with fundamental roles in intracellular glucose metabolism. Thiamine also regulates the expression of genes that code for enzymes using thiamine as a co-factor. Thiamine deficiency reduces the mRNA levels of transketolase and PDH (34). In the blood of patients with cancer, the concentration of thiamine and thiamine diphosphate (TDP) are lowered by 20%; transketolase activity is decreased by 20%, and the TDP effect by 5-42% (35). Moreover, aberrant levels of transketolase and transketolase-like-1 (TKTL-1) participated in glucose metabolism in malignant cells in pleural effusion (36). TKTL-1 has been reported to play a pivotal role in carcinogenesis and has an important role in controlling the non-oxidative pentose-phosphate pathway, which is significantly up-regulated in cancer. Increased TKTL-1 expression has been shown to be correlated with poor patient outcome and increased tumor progression (37). Additionally, increased TKTL 1 has been observed in many types of cancer, including thyroid cancer (38, 39), nasopharyngeal carcinoma (40, 41), non-small cell lung cancer (42, 43), gastric cancer (44), human hepatoma cell (45), colorectal cancer (46), urothelial cancer (47), anaplastic nephroblastoma (48), breast cancer (49), endometrial cancer (50), ovarian cancer (51, 52), cancer of the uterine cervix (53), head and neck cancer (54), and glioblastoma (55). However, the related proteins TKTL-1 and TKTL-2 lack numerous invariant residues involved in co-factor and substrate binding of TPP and are therefore not expected to possess transketolase activity (56).
Transcription factor p53. The p53 gene and protein play a critical role in the regulation of the normal cell cycle, cell cycle arrest, and the apoptotic response. p53 is a transcription factor with a major role in determining cell fate in response to DNA damage. p53 reduces the incidence of cancer by promoting apoptosis in cells that have activated oncogenic pathways. p53 polymorphisms have been demonstrated in breast cancer (57), mesothelioma (58), esophageal squamous cell carcinoma (59), and pancreatic cancer (60). In many meta-analyses, p53 polymorphisms have been reported to be associated with cervical (61), gastric (62), bladder (63), colorectal (64), and endometrial (65) cancer, but not with prostate cancer (66). Increased thiamine transporter levels have been observed in murine cells that overexpress mouse SLC19A2 or in cells exposed to conditions that induce DNA damage or p53 activation (67). TDP has been shown to inhibit p53 binding and thiamine has been shown to inhibit intracellular p53 activity (68). The expression of p53 decreased significantly in cultured retinal neurons of diabetic rats treated with thiamine (69). These observations suggested that the transcription factor p53 is activated in cancer with increasing apoptotic response from cellular damage and thiamine ameliorated these effects on cells.
Poly(ADP-ribose) polymerase-1 (PARP-1). PARP 1 is a nuclear protein that contributes to both cell death and survival under stressful conditions. PARP-1 catalytic activity is stimulated by DNA strand breaks. In mice, Parp-1-deficient cells exhibited enhanced sensitivity to lethal effects of ionizing radiation and alkylating agents (70), and Parp-1 knockout mice were found to develop spontaneous mammary and liver tumors (71, 72). Overexpression of PARP-1 has been reported in Ewing’s sarcoma (73), malignant lymphoma (74), colorectal cancer (75), hepatocellular carcinoma (HCC) (76), breast cancer (77), pediatric CNS tumors (78), and ovarian cancer (79). In a meta-analysis, PARP-1 mRNA expression was correlated with high-grade, medullary histological type, tumor size, and worse metastasis-free survival and overall survival in human breast cancer (80). PARP-1 polymorphisms have been found to be associated with gastric cancer in Han Chinese (81), prostate cancer (82), esophageal squamous cell carcinoma (83), and lung cancer (84), and with a reduced risk of non-Hodgkin lymphoma in Korean males (85). PARP 1 polymorphisms reduce PARP-1 catalytic activity by 30-40% (86). In a meta-analysis, no significant association was observed between the PARP-1 V762 polymorphism and cancer risk; however, the variant A allele of the PARP-1 V762 polymorphism was associated with an increased risk of cancer within Asian populations, but with a decreased risk of cancer among Caucasians, particularly for glioma (87). However, thiamine has a cytoprotective effect on cultured neonatal rat cardiomyocytes under hypoxic insult; thiamine also inhibits PARP cleavage and DNA fragmentation (88). Benfotiamine, a fat-soluble thiamine analog, prevents bacterial endotoxin-induced inflammation and PARP cleavage in mouse macrophage cell lines (89). Adenosine thiamine triphosphate (ATTP) has been identified in small amounts in the mouse brain, heart, skeletal muscle, liver and kidneys (90) and has been shown to inhibit PARP-1 activity (91). Taken together, the results indicate that thiamine may play a role in cancer by modulating PARP-1 activity.
The reduced form of nicotinamide adenine dinucleotide phosphate (NADPH). The NADPH oxidase (NOX) enzyme complex mediates critical physiological and pathological processes, including cell signaling, inflammation and mitogenesis, through the generation of reactive oxygen species (ROS) from molecular oxygen. NOX1 is required for RAS oncogene-induced cell transformation (92). The NOX subunit p22phox has been reported to inhibit the function of the tumor suppressor protein tuberin in renal carcinoma cells (93). The activation of NOXs has been demonstrated in the development of cancer, including melanoma (94), leukemia (95), esophageal adenocarcinoma (96), HCC (97), prostate cancer (98), colon cancer (99), glioblastoma multiforme (100), and multiple myeloma (101). NADPH polymorphisms have been reported to be associated with myelodysplastic syndrome and de novo acute myeloid leukemia (102, 103), esophagus cancer (104), lung cancer (105, 106), non-Hodgkin’s lymphoma (107, 108), childhood acute leukemia (109), postmenopausal breast cancer (110), and gastric cancer (111). The levels of NADPH cytochrome c reductase are increased in thiamine-deficient animals (112). The expression of NOX4 is significantly down-regulated by benfotiamine treatment under both normo- and hyperglycemic conditions (113). In addition, animals fed a high thiamine diet exhibited NADPH-cytochrome c reductase activity of approximately 57% that of those fed a thiamine-deficient (TD) diet (114). Altogether, these results suggest that thiamine may produce a cancer-protective effect by regulating NADPH-cytochrome c activity.
The genetic factors related to thiamine and cancer are summarized in Table I.
The Role of Thiamine in Cancer
Diabetes mellitus. Alterations in glucose metabolism have been linked to a significantly increased risk for many forms of cancer. Gestational diabetes mellitus (DM), a state of glucose intolerance associated with pregnancy, is associated with an increased risk of pancreatic cancer and hematological malignancies (115). DM and impaired glucose metabolism contribute to the risk of postmenopausal breast and endometrial cancer (116). The metabolic syndrome has been reported to be associated with the risk of prostate cancer in Italian populations (117). Type 1 DM is associated with an increase in the risk of cancer, particularly gastric, uterine cervix, endometrial, and ovarian cancer (118, 119). Several meta-analyses have demonstrated an association between DM and increased cancer risk, including esophageal (120), gastric (121), colorectal (122), HCC (123), cholangiocarcinoma (124), and breast cancer (125). The incidence of DM in renal cell carcinoma is higher in females than males and higher in Hispanic populations compared to Caucasian populations and other ethnicities (126). However, a relationship between thiamine and DM has also been reported in the literature. A significant proportion of healthy individuals (36-47%) have been reported to have thiamine deficiency while in a hyperglycemic state (such as on a diet high in carbohydrates or if diabetic or pregnant) (127). Low plasma thiamine levels have been noted in patients with DM type 1 (128). Thiamine reserves are reduced in litters of untreated diabetic rats (129). In children, acute thiamine deficiency can be manifested as diabetic ketoacidosis (DKA), lactic acidosis and hyperglycemia (130, 131). In another study, low blood thiamine levels in the blood and erythrocyte transketolase activity, and high erythrocyte TPP activity were documented in patients with diabetes (132, 133). The activity of transketolase has been used to assess thiamine activity in mammalian tissues. The low thiamine values in patients with diabetes might be a reflection of reduced apo-enzyme levels from the disease itself rather than from a thiamine deficiency (134). In addition, plasma thiamine levels are decreased by 76% in type 1 and 75% in type 2 DM; decreased plasma thiamine levels have also been associated with increased renal clearance and fractional excretion of thiamine (134). Furthermore, thiamine transporter protein concentrations are increased in erythrocyte membranes in type 1 and type 2 DM. Therefore, changes in thiamine levels may be masked by an increase in thiamine transporter expression (134).
Genetic factors related to thiamine and cancer.
Cancer. Thiamine levels are decreased in many cancer subjects. Hepatoma induced by butter-yellow exhibited a decrease in thiamine and co-enzyme A concentrations by more than 50% and 92%, respectively, compared to normal livers (135). Thiamine deficiencies have also been noted in patients with uterine cancer (136), B-chronic lymphocytic leukemia (137), and colorectal cancer (138). During the growth of Ehrlich’s ascites tumor, the concentration of TDP in the liver of tumor-bearing animals continuously drops for 10 days after inoculation, and the TDP level in the tumor itself decreases more abruptly by the terminal period of tumor growth (10th day) (139). Some authors have suggested that thiamine supplementation may contribute to the high rate of tumor cell survival, proliferation and chemotherapy resistance (140). A significantly higher incidence of urinary bladder carcinomas was induced in rats fed bracken fern, which contains a carcinogen agent, and additionally supplemented with thiamine than observed in rats fed only bracken fern (141). In melanoma cells, overexpression of the melanoma tumor antigen p97 resulted in decreased thiamine triphosphatase levels that may lead to greater TPP and increased cellular proliferation (142). TPP inhibits cytotoxicity of methotrexate, an antimetabolite used for the treatment of cancer (143). In vitro, thiamine blocks the antitumor effect of mechlorethamine in murine leukemia cells (144) and blocks the antitumor effect of cyclophosphan in Ehrlich’s ascites tumor (145). The administration of cyclophosphan further increased thiamine deficiency in tumor-bearing mice. Simultaneous administration of thiamine and cyclophosphan abolished the cytostatic toxic effect but did not affect their antitumoral properties (145). Furthermore, low thiamine levels were of benefit against some types of cancers. Thiaminase I digests thiamine and has been reported to have a growth-inhibitory activity against breast cancer cell lines (146) and lymphoid leukemia cell lines (147). In addition, MDA231 breast cancer xenografts exhibited growth delay in mice fed a thiamine-free diet (146). Injections of oxythiamine increased the rate of pathological mitoses in Walker 256 carcinoma cells, reduced the tumor weight by 45%, and inhibited the synthesis of TDP and activity of transketolase in tissues (148). However, marginal dietary thiamine deficiency induces the formation of aberrant colonic crypt foci in rats (149). In Syrian hamsters, which are susceptible to oral tumor induction by chemical agents, a dietary regime low in thiamine resulted in the development of malignant neoplasms arising in the pouch epithelium in a significantly shorter period of time than in hamsters fed a diet containing adequate amounts of this vitamin (150). Moreover, thiamine did not induce death or increase the proliferation rate of Hepa or HepG2 HCC cell lines, either in vitro or in vivo, and thiamine was reported to attenuate Wilson’s disease-induced HCC (151). In another report, thiamine did not stimulate tumor growth in patients with gastric tumors (145). In a patient with osteosarcoma, subcutaneous perfusion of thiamine reduced the tumor circumference from 30 to 20 cm, equivalent to a reduction of 50-75% in volume, within two days (152). A metabolic control analysis demonstrated a high stimulatory effect on tumor growth of 164% compared with controls with a thiamine dose of 25-fold the recommended dietary allowance (RDA); however, at very high doses of thiamine, ∼2500-fold the RDA, the opposite effect was observed, producing an inhibitory effect on tumor growth of 36% compared with control animals (153). The Authors suggested that when thiamine supplementation is necessary for patients with cancer, it should be administered at high doses to avoid the tumor-promoting effect of low doses. Furthermore, thiamine analog benfotiamine significantly reduced genomic damage of peripheral lymphocytes in hemodialysis patients, who exhibited enhanced DNA damage in end-stage renal disease, independent of changes in the plasma levels of advanced glycation end product (154).
Matrix metalloproteinases (MMPs). MMPs are proteolytic enzymes responsible for both extracellular matrix (ECM) remodeling and the regulation of leukocyte migration through the ECM, which is an important step in the inflammatory processes. MMP overexpression has been reported in many types of cancer, including malignant peripheral nerve sheath tumors (155), non-small cell lung cancer (156), glioblastoma multiforme (157), sarcoma (158), colorectal cancer (159), prostate cancer (160), and ovarian carcinoma (161). MMP polymorphisms are associated with breast cancer (162), colorectal cancer (163), renal cell carcinoma (164), adult astrocytoma (165), endometrial carcinoma (166), bladder cancer (167), epithelial ovarian cancer (168), oral cancer (169), cervical cancer (170), esophageal squamous cell carcinoma (171), gastric cancer (172), malignant head and neck tumors (173), hypophyseal adenoma (174), prostate cancer (175), glioblastoma multiforme (176), and HCC (177). In meta-analyses, MMP-2 polymorphisms have been linked to the risk of lung (-735 C/T) (178) and breast cancer (-1306 C/T) (179), but MMP-2 -1306 C/T allele may be a protective factor for digestive cancer risk (180). In other studies, polymorphisms of the MMP-1, -2, 3, and -7 may have played allele-specific roles in cancer development (181, 182). However, MMP-9 has also been shown to be up-regulated in the TD mouse brain (183, 184). Thiamine reduces the MMP-2 activity in the heart of diabetic rats and prevents diabetes-induced cardiac fibrosis (185). Moreover, thiamine and benfotiamine correct the increase in MMP-2 activity that results from high glucose levels in human retinal pericytes, while increasing tissue inhibitor of metalloproteinase-1 (TIMP-1) (186). Altogether, these studies suggest that thiamine may play an important role in the pathological processes of cancer by modulating the levels of MMPs and TIMPs.
Prostaglandins (PGs) and Cyclooxygenase-2 (COX2). PGS play a role in inflammatory processes. COX participates in the conversion of arachidonic acid into PGs. The expression of COX2 mRNA and PGE2 were selectively increased in vulnerable regions in the symptomatic stages of TD encephalopathy animal models (187). Up-regulation of 15-hydroxyprostaglandin dehydrogenase (15-PGDH) expression was observed in breast cancer cell lines transfected with SLC19A3 gene and down-regulation was observed after suppression of SLC19A3 with siRNA vectors (188). Overexpression of 15-PGDH inhibited interleukin (IL-1β)-induced COX2 expression (189). In murine macrophages, benfotiamine also blocked the expression of COX2 and its product PGE2 by lipopolysaccharide (LPS)-induced cytotoxicity (190). In addition, benfotiamine significantly prevented LPS-induced macrophage death and monocyte adhesion to endothelial cells. These anti-inflammatory effects of benfotiamine are mediated through the regulation of the arachidonic acid pathway in macrophages (191). Tumor inflammation is now recognized as one of the hallmarks of cancer. Over-expression of COX2 has been associated with resistance to apoptosis, increased angiogenesis and increased tumor invasiveness in various types of cancer. Increased COX2 expression has been reported in endometrial adenocarcinoma (192), breast cancer (193), reno-medullary interstitial cell tumor (194), colorectal carcinomas (195), gastric cancer (196), carcinoma of the cervix (197), and familial adenomatous polyposis (198). Deletion of Cox2 in mouse mammary epithelial cells delayed breast cancer onset (199). COX-2 inhibitors also reduced the growth and induced regression of human esophageal adenocarcinoma xenografts in nude mice (200), and retarded murine mammary tumor progression by reducing tumor cell migration, invasiveness, and angiogenesis (201). Genetic variability in enzymes could impact on the risk of the disease. COX2 polymorphisms have been associated with bladder (202), biliary tract (203), lung (204), non-melanoma skin cancer after organ transplantation (205), esophageal squamous cell carcinoma (206), nasopharyngeal carcinoma (207), pancreatic (208), invasive ovarian carcinoma (209), breast (210), gastric carcinoma (211), acute myeloid leukemia (212), prostate (213), head and neck (214), colorectal adenoma (215), and HCC (216). In meta-analyses, the COX2 1195G>A polymorphism was significantly associated with an increased risk of digestive system cancer, especially in Asian populations (217), and the -765G>C variant may confer an increased risk of colorectal carcinoma and esophageal cancer in Asians. The 8473T>C polymorphism may confer a decreased risk of breast and lung cancer (218). In addition, PGE2 has been reported to be associated with colorectal adenoma (219), pancreatic tumor (220), and childhood neuroblastoma (221). The suppression of PGE2 receptor inhibited human lung carcinoma cell growth (222). These findings suggest that thiamine may play a role in modulating the inflammatory process of cancer.
Reactive oxygen species (ROS). ROS play a major role in various cell-signaling pathways. ROS activates various transcription factors and increases in the expression of proteins that control cellular transformation, tumor cell survival, tumor cell proliferation and invasion, angiogenesis, and metastasis (223). ROS also play an important role in the initiation and progression of many types of cancer (224-229). Single-nucleotide polymorphisms of antioxidant defense genes may significantly modify the functional activity of the encoded proteins. Women with genetic variability in iron-related oxidative stress pathways may be at increased risk for postmenopausal breast cancer (230). The Ala variant of superoxide dismutase (SOD) has been associated with a moderately increased risk of prostate cancer (231). In meta-analyses, manganese SOD polymorphisms may contribute to cancer development (Val-9Ala) (234) and prostate cancer susceptibility (Val-16Ala) (232) but not breast cancer susceptibility (Val-16Ala) (233). However, oxidative stress has been associated with region-specific neuronal death, and lipid peroxidation products accumulated in the remaining thalamic neurons after 11 days of TD animal models (234). In vitro, thiamine inhibits lipid peroxidation in rat liver microsomes and free radical oxidation of oleic acid (235). Benfotiamine promotes a reduction in ROS-induced by advanced glycated albumin in macrophages (236). In primary human peritoneal mesothelial cells of a rat model of peritoneal dialysis, the addition of benfotiamine enhanced transketolase activity and reduced expression of advanced glycation end products and their receptor (237). These data suggest that benfotiamine protects the peritoneal membrane and remnant kidney in such a rat model. Thiamin rescues hepatocytes from iron-catalyzed oxidative stress by reducing lipid peroxidation, mitochondrial and protein damage and DNA oxidation (238). These findings suggest that thiamine modulates oxidative stress in cancer.
Nitric oxide synthase (NOS). NOS is an enzyme involved in the synthesis of nitric oxide (NO), which regulates a variety of important physiological responses, including cell migration, immune response and apoptosis. NO and calcium have been reported to regulate mitochondrial biogenesis in follicular thyroid carcinoma cells (239). There is a link between NO and the induction of apoptotic cell death in head and neck squamous cell carcinoma development (240). Cytokines especially interferon-gamma was found to induce apoptosis in acute leukemia via the NO and caspase-3 pathway (241). The reduction of NO enhanced the radiosensitivity of hypoxic non-small cell lung cancer (242). Increased NO may be a sign of subclinical cardiotoxicity of doxorubicin (243). High NO concentrations at the periphery of a melanoma may contribute to metastasis by stimulating cell proliferation, inhibiting apoptosis, or acting as a lymphangiogenic factor (244). Inducible NOS (iNOS) mRNA expression was considerably higher in glioblastoma than meningioma specimens (245). iNOS expression has been correlated with angiogenesis, lymphangiogenesis, and poor prognosis in gastric cancer (246) and in estrogen receptor-negative breast cancer (247). NOS inhibition enhanced the antitumor effect of radiation in the treatment of squamous carcinoma xenografts (248). NOS polymorphisms have been associated with bladder (249), urothelial carcinoma (250), gastric (251), colorectal cancer (252), and non-Hodgkin’s lymphoma (253). In a meta-analysis study, the endothelial NOS 894G>T polymorphism was associated with breast cancer (254). However, increased brain endothelial NOS expression was demonstrated in TD (255). In murine macrophages, benfotiamine was also found to block the expression of iNOS by LPS-induced cytotoxicity (189).

Summary the role of thiamine in cancer.
The role of thiamine in cancer.
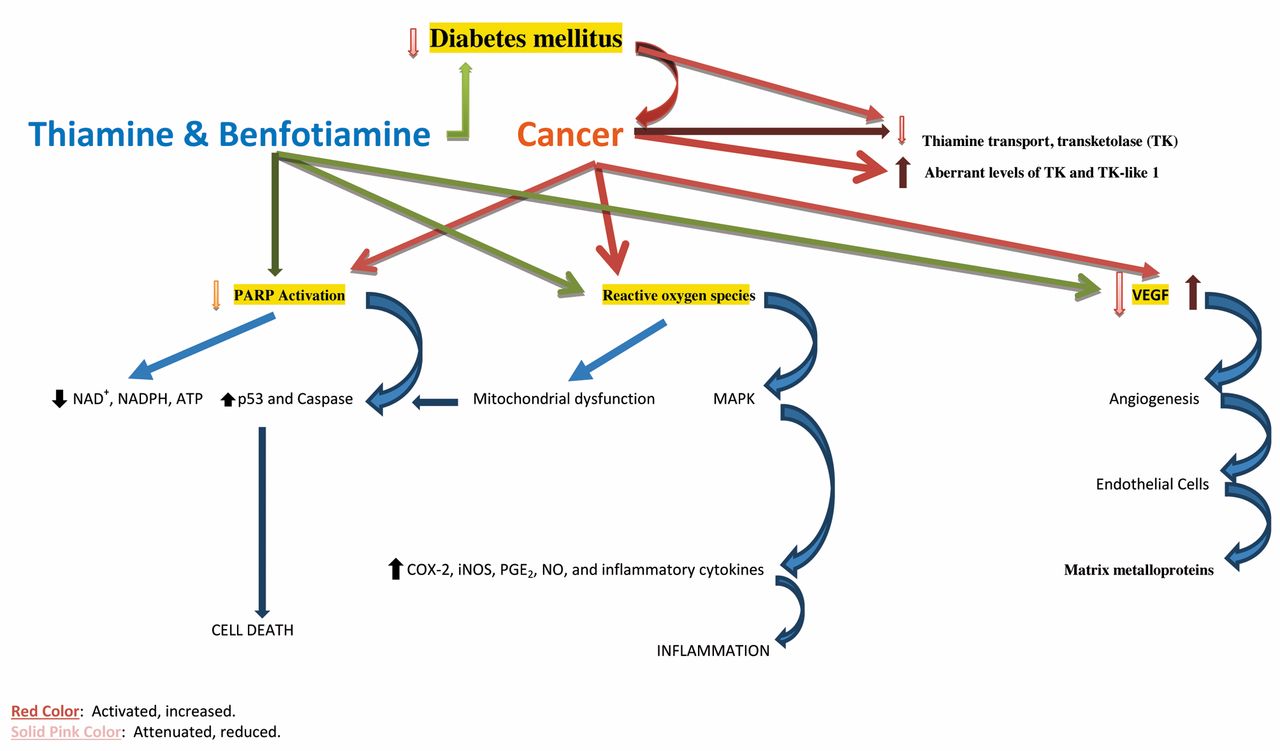
Table II illustrates the role of thiamine in cancer and Figure 1 summarizes the role of thiamine in cancer.
Conclusion
Genetic studies have provided opportunities to determine which proteins may link thiamine to cancer pathology, including SLC19 gene, transketolase, transcription factor p53, PARP, and NOX. Thiamine can also act through a number of non-genomic mechanisms, which include protein expression, oxidative stress, inflammation, and cellular metabolism. Thiamine supplementation may stimulate a high rate of tumor cell survival, proliferation and chemotherapy resistance, but other studies have demonstrated the beneficial role of thiamine in cancer. However, thiamine deficiency may occur in patients with cancer and may produce serious disorders, including Wernicke’s encephalopathy, that require parenteral thiamine supplementation. In the early stages of TD encephalopathy (Wernicke’s encephalopathy), patients were found to respond rapidly to large doses of parenteral thiamine. The initial dose of thiamine is usually 100 mg two to three times daily for one to two weeks. A very high dose of thiamine has been shown to exhibit a growth inhibitory effect in cancer. Thiamine appears to have both cancer-promoting and anticancer properties, which vary in relation to both genomic and non-genomic factors. However, thiamine absorption decreases with advancing age (256). Therefore, further investigations on parenteral thiamine in cancer are needed. Indeed, a more cautious approach would be advisable before recommending the widespread use of thiamine in patients with cancer.
Footnotes
-
Ethical Approval
Not required.
-
Funding
The Authors declare that they received no funding.
-
Conflicts of Interest
Authors declare that they have no competing interests.
- Received April 12, 2013.
- Revision received May 3, 2013.
- Accepted May 9, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵




{kind=link}