


Abstract
The advent of various technologies for the generation of bi- and multispecific recombinant antibody-based molecules brought with it a multitude of formats for selecting target combinations. Some of the format options are outlined from a technical point of view. We focus on the achievements and prospects of the underlying technologies for generating bi- and multispecific antibodies to i) target immune effector cells and/or cytokines to tumors, ii) engage death receptors on tumor cells simultaneously, iii) improve antiangiogenic intervention by blocking complementary pathways of angiogenesis and iv) achieve more efficient targeting of human epidermal growth factor-related and other receptor tyrosine kinase-related pathways. Many of the outlined approaches, in addition to potential improvement of therapeutic efficacy in comparison to single agent intervention, also offer the potential to counteract therapy resistance.
- Antibody formats
- cytokine delivery
- death receptors
- HER signaling
- recruitment of immune effector cells
- inhibition of angiogenesis
- review
Bevacizumab, rituximab and trastuzumab are approved monoclonal antibodies (mAbs) for treatment of cancer and are among the ten top-selling protein therapeutics (1). Specifically, their mode of action involves blockade of ligand binding and signal perturbation, targeting tumor cells and the tumor microenvironment, resulting in the inhibition of angiogenesis and the enhancement of antitumor responses by recruiting immune effector cells (2). Antibody-mediated cellular cytotoxicity (ADCC), antibody-dependent cellular phagocytosis (ADCP), complement-dependent cellular cytotoxicity (CDC) and induction of T-cell immunity through cross-presentation are also possible mediators of their mode of action (3-5). The antitumor efficacy of mAbs can be enhanced by combination with other immunomodulatory approaches such as chemotherapy, radiotherapy and target-specific small-molecule therapeutics. However, according to the hallmarks of human cancer, which include deregulation of several pathways (6, 7) and also cross-talk between pathways and their role in resistance to therapeutic agents (8, 9), simultaneous modulation of several targets with bi- or multispecific antibodies might result in an increased therapeutic benefit. Dual engagement of death receptors, concurrent dual blockade of receptor tyrosine kinases, as well as recruitment of immune effector cells and cytokines to tumors by bi- or multispecific antibodies, are themes discussed in this review.
Formats
More than 20 years ago, Shery Morrison and colleagues provided the first description of the concept of recombinant bispecific antibodies (10). Since then, academia and corporate research have devised a multitude of formats suitable for generation of bispecifics. The formats of these molecules can be assigned to two general classes: those entities with and those without immunoglobulin G (IgG). Molecules of these classes differ in size, are frequently produced by different technologies, and have different in vivo distribution, tissue penetration and pharmakokinetic (PK) properties. And within each format class, another level of format variability exists with regard to binding stoichiometry and geometry, steric considerations, crosslinking capability and avidity.
The format class of ‘IgG-derived molecules’ characterized by containing Fc regions (Figure 1A) are frequently produced in mammalian cells. The presence of Fc regions enables binding to protein A. This makes their production compatible with upstream and downstream processes that are well established for production of ‘normal’ IgGs. Bispecific antibody derivatives of this class have a relatively large size (>100.000 Da) and frequently have a long serum half-life due to Fc-mediated neonatal Fc receptor (FcRn) binding and recycling.
Building upon the Morrison concept, a variety of formats were generated which contain additional binding entities attached to N- or C-termini of antibodies. These fusions with single-chain or disulfide stabilized fragment, variable region (Fvs) or fragment, antigen binding (Fabs) result in the generation of tetravalent molecules with bivalent binding specificity for each antigen (11-13). In a similar manner, combinations of single chain (sc) Fvs and scFabs with IgGs enable the production of molecules which can recognize three (or more) different antigens (14). A related format is that of dual-variable domain immunoglobulins (DVDs). These are composed of variable region heavy chain (VH) and variable region light chain (VL) domains of a second specificity placed upon the N-termini of the V domains by short linker sequences (15, 16) (Figure 1A).
Finally, bispecific Fc-containing entities (mini-antibodies) can be generated by fusing scFv to the C-termini of constant region heavy chain domain 3 (CH3-scFv) and/or to the hinge region (scFv-hinge-Fc) of an antibody with a different specificity. Trivalent entities have also been described which harbor disulfide-stabilized variable domains (without peptide linker) fused to the C-terminus of CH3 domains of IgGs. These bispecifics have a bivalent binding mode for one antigen and bind the second antigen in a monovalent manner (17) (Figure 1A). Monovalent binding to the second antigen can be further modulated by generating ‘cloaked’ molecules which become fully binding competent only upon proteolytic processing (17).
Alternatively to bispecifics composed of additional binding entities added to an IgG scaffold, bispecificity can also be directly introduced into the bivalent (Y-shaped) IgG structure. Such molecules possess two binding arms which recognize different antigens. One means of achieving that is via display technologies to evolve variable domains capable of recognizing two unrelated antigens (18, 19) (Figure 1A and 2A). IgGs that contain such variable regions bind two antigens, with each V-domain being occupied once. Thus, each of the cognate antigens can be either bound bivalently or monovalently.
IgG-derived formats that enable defined monovalent (and simultaneous) antigen binding are generated by forced heavy (H)-chain heterodimerization, combined with technologies that minimize light (L)-chain mispairing. Generation of antibody H-chain heterodimers is in many instances achieved via application of the knobs-into-holes principle (20), replacing a small amino acid with a larger one in one of the CH3 domains (‘knob’) and simultaneously introducing smaller amino acids (‘hole’) into the CH3 domain of the second H-chain. H-Chain heterodimerization can also be modulated via strand-exchange engineered domains (SEED) (21, 22) (Figure 1A). This principle is based upon sequence exchanges between IgG1 and IgA to create non-identical chains which heterodimerize preferentially. In addition to assurance of H-chain heterodimerization, L-chain mispairing must be avoided to generate homogenous preparations of bispecific IgGs. One way to achieve this is through the use of the ‘common light chain’ principle, i.e. combining two binders that share one light chain but still have separate specificities (23). Another option is CrossMab technology (24, 25), which avoids non-specific L chain mispairing by exchanging CH1 and CL domains in the Fab of one half of the bispecific antibody (Figures 1A and 2B). Such crossover variants retain binding specificity and affinity, but make the two arms so different that L-chain mispairing is prevented.
The ‘Fc-less bispecifics’ format class (Figure 1B) is characterized by having a generally smaller size than Fc-containing entities and these are frequently produced in bacterial expression systems. Bispecific antibody derivatives (Figures 1B and 2C) of this class often have a short serum half-life due to their small size (renal clearance) and due to lack of interaction with the neo-natal Fc receptor (26). The addition of an Fc domain to an antibody fragment (see above), or of albumin binding entities (27, 28) is a clear alternative for increasing the serum half-life. Alternatively, improvement of PK properties of small bispecific antibody fragments has been achieved by conjugation with polymers such as polyethylene glycol (29). Molecules of this class are composed of individual binding modules such as scFvs or Fabs which are connected to each other via flexible linker peptides. Bispecificity is either achieved by appropriate choice of fusion partners, or by inclusion of heterodimerization (and/or trimerization) functionalities.
One format that is devoid of all antibody constant domains are bispecific T-cell engagers (BiTEs) (30) (Figures 1B and 2C). These bispecific binders are created by fusing two scFvs via a flexible linker peptide. These will be discussed in more detail below. Another format for bispecific binders lacking constant region sequences is based on diabodies (19) (Figure 1B and 2D). These consist of two VH and two VL domains from two different antibodies. Interaction only with complementary domains on another chain is achieved by attaching domains with short linker peptides which permits pairing only with VH and VL domains. VH of the first binder linked to the VL of the second binder is coexpressed with the VH of the second antibody linked to VL of the first antibody. Based on the diabody technology, tetravalent tandem antibodies (TandAbs) (Figure 1B and 2E) were designed to increase the number of valences, stability and therapeutic potential (31, 32). These molecules are generated by functional dimerization of a protein consisting of four antibody variable H- and L-chains in an orientation that prevents intramolecular pairing. A further modular bispecific antibody platform applies dual affinity retargeting molecules (DART), entities that are stabilized by disulfide bonds which apply a similar design concept to that of diabodies (33) (Figure 1B).
One example for generation of bispecifics via added entities that support hetero-dimerization is the ‘dock-and-lock’ principle for the construction of multispecific antibodies. This technology is based on the natural binding by the dimerization and docking domain (DDD) of cAMP-dependent protein kinase A to the anchoring domain (AD) of A-kinase anchoring protein (34). DDDs are linked to a Fab fragment resulting in dimerization and thus the generation of a divalent Fab fragment. This creates a docking site for binding to the AD sequence incorporated into a second protein, resulting in the formation of a trivalent complex. The trivalent complex is stabilized by disulfide bridges between inserted cysteine residues.
Initial Efforts to Recruit Immune Effector Cells Using Bispecific Antibodies
Tumor-associated antigens, such as cluster of differentiation 19 (CD19), CD20, human epidermal growth factor receptor 2 (HER2), carcinoembryonic antigen (CEA), and epithelial cell adhesion molecule (EpCAM), were initial targets for redirecting immune effectors to tumor cells. For the recruitment of immune effector cells, CD3 on T-cells, CD16 (FcγRIII) on natural killer (NK) cells and CD64 (FcγRI) on monocytes and neutrophils, were the preferred targets. Chemically cross-linked antibodies represented the first generation of bispecific antibodies. No impact on disease improvement was noticed in clinical studies. Immunogenicity of the murine antibody fragments and the difficulties associated with the production of homogeneous batches were partly responsible for this disappointing outcome (35). A further issue is that the mere engagement of the activating receptors by bispecific antibodies as outlined is not sufficient to induce target cell killing due to the requirement for co-stimulatory signals (36). In addition, high mAb doses and a high effector-to-tumor (E/T) cell ratio is necessary for killing, conditions which are not easily achieved in vivo (25). As discussed later, new formats have significantly improved these issues.
Targeting CD28 for Recruitment of Immune Effector Cells
One of the alternative strategies involved targeting CD28 based on the finding that supra-agonistic CD28 antibodies induce potent T-cell activation without a primary T-cell receptor/CD3 (TCR/CD3) stimulus (37). A bispecific antibody directed against CD20 and CD28 was shown to mediate target cell-restricted, supra-agonistic CD28 stimulation and killing of lymphoma cells in the presence of autologous normal or malignant B-cells (38). Two scFv fragments were connected by a flexible linker as a basic design of this bispecific antibody. Analogously constructed antibody r28M directed against melanoma-associated proteoglycan neural glial antigen 2 (NGA-2) and CD28 resulted in killing of NGA-2-positive tumor cells and did not require a primary signal through the TCR/CD3 complex (37). The striking supra-agonistic activity of this recombinant bispecific antibody is most likely due to homodimerization as suggested by gel filtration analysis. It was shown that the lytic activity measured after three days of supra-agonistic CD28 stimulation of peripheral blood mononuclear cells (PBMCs) is partly mediated by non T-cells (39). Subsequently this bispecific antibody was expressed in transgenic cows and isolated from their serum (40, 41). In vitro, T-cells and NK cells contributed to tumor cell killing after r28M-mediated activation of PBMCs. In vivo, r28M inhibited growth of orthotopically implanted U87M glioblastoma cells together with PBMCs after intralesional application. The lytic activity of the non-T-cell population is largely mediated by CD56-positive NK cells. If they are separated from T-cells, they are not active, indicating that NK cells seem to be activated by T-cell-derived cytokines such as interleukin 2 (IL2), interferons (IFNs), tumor necrosis factor (TNF) and interleukin 4 (IL4). A half-life of 6 h was noted after i.v. injection of this bispecific antibody. On the other hand, a cytokine storm induced by the monoclonal antibody to CD28, TGN 1412, was observed in a phase I trial, resulting in life-threatening syndromes (42). In all six volunteers, 90 min after administration of the antibody, systemic inflammatory response and rapid induction of proinflammatory cytokines accompanied by headache, myalgia, nausea, diarrhea, erythema, vasodilatation and hypotension was observed.
Trifunctional Bispecific Antibodies
Rat/mouse quadromas circumvent the problem of promiscuous chain associations observed in mouse/mouse and rat/rat quadromas. In these hybrid quadromas, species-specific H/L chain pairing is observed (43). The resulting bispecific antibodies are composed of mouse IgG2a and rat IgG2b and can be easily purified by protein affinity chromatography. Their trifunctionality is based on targeting a tumor antigen, the recruitment of T-effector cells by interaction with CD3, and the binding and activation of Fcγ receptor-positive accessory cells such as monocytes/macrophages, NK cells and dendritic cells via its Fc region (44). After interaction of these bispecific antibodies with tumor cells in the presence of PBMCs, up-regulation of co-stimulatory molecules such as CD40, CD80 and CD86 and the production of cytokines such as IL2 and IL6 was observed (44).
Specifically, these antibodies were directed to EpCAM, HER2, CD20 or gangliosides GD2 and GD3 (45-48). For example, Catumaxomab (anti-EpCAM/anti-CD3) killed tumor cells very efficiently at low concentrations without any stimulation of effector cells (45). Bi20 (anti-CD20/anti-CD3) at antibody concentrations as low as 0.5 ng/ml in vitro, eliminated 70% of Raji tumor B-cells within 24 h and killed tumor cells completely after three days (47). One hundred percent (n=12) of mice pre-administered with 4 μg of a trifunctional antibody survived after i.p. injection of a lethal dose of melanoma cells (47). Another important feature is the capability of trifunctional antibodies to kill cells expressing low levels of target antigens such as HER2 or CD20, as well as trastuzumab- or rituximab-resistent tumor cells (46, 47). Induction of long-lasting, protective antitumor immunity by bifunctional antibody Bi20 was demonstrated for syngeneic B16 melanoma and A-20 B-cell lymphoma mouse tumors (49). Survival correlated with the induction of a humoral immune response (anti-idiotypic antibodies) and T-cell depletion experiments demonstrated the contribution of a T-cell-based antitumor response. The role of the Fc region in this context was demonstrated by the significantly diminished survival time when two antigen-binding fragments [F(ab)2] of the corresponding bispecific antibody were used.
Catumaxomab prevented the accumulation of ascites and efficiently eliminated tumor cells in patients with ovarian carcinoma with an acceptable safety profile and was approved in 2009 for treatment of malignant ascites in patients with EpCAM-positive ovarian carcinoma for whom standard therapy was no longer feasible (50). Extremely small doses of antibodies (5-20 μg) with four to five i.p. infusions within 9 to 13 days were administered with moderate anti-mouse or anti-rat immunoglobulin responses in the majority of the patients, which did not affect the efficacy of the treatment. Finally, Ertumaxomab (anti-HER2/anti-CD3) and Bi20 are currently being evaluated in clinical studies in patients with metastatic mammary carcinoma and recurrent B-cell lymphoma, respectively (51).
Bipecific T-Cell Engagers (BiTEs)
BiTEs (Figure 1B and 2C) are composed of two scFvs connected by a flexible linker (52-54) and can exert potent antitumor cell cytotoxicity (55). In this approach, one of the scFvs is directed against a tumor-associated antigen, while the second scFv is directed against the CD3έ subunit of the TCR complex. In the following examples the affinity of the antibody directed against the tumor-associated antigen is two orders of magnitude higher than that directed against CD3.
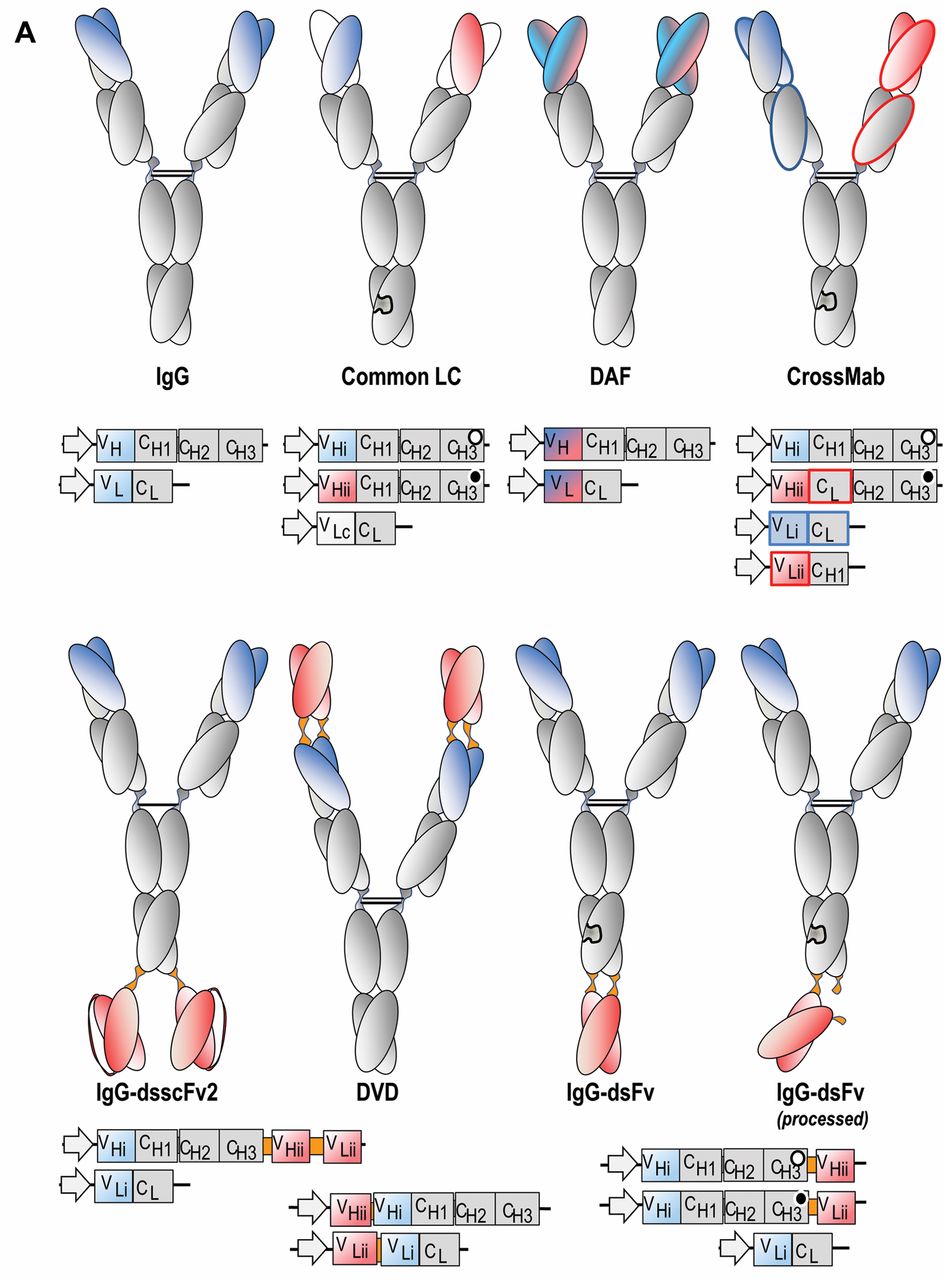
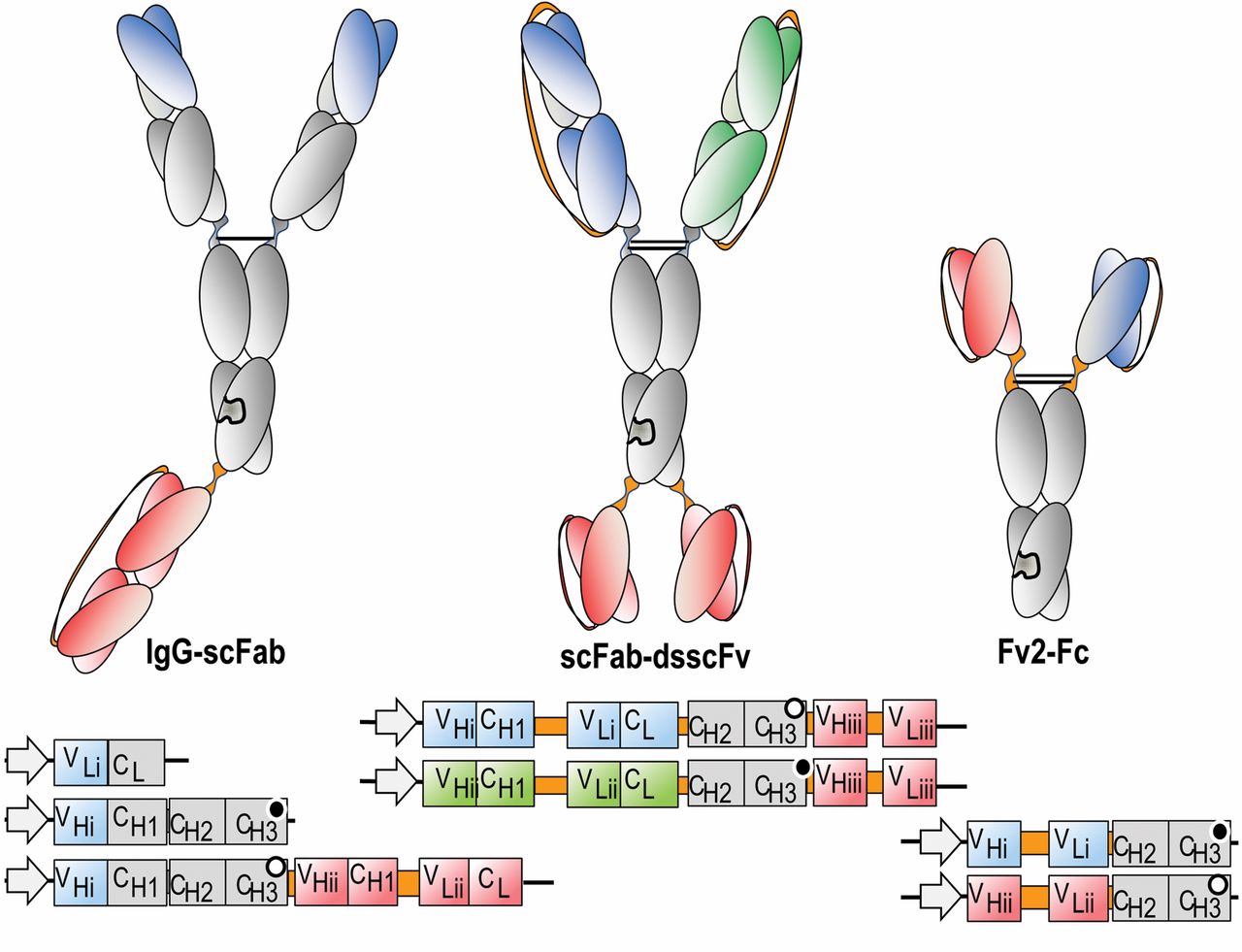

Schematic composition of bispecific antibody derivatives. The domain compositions of various bispecific antibody formats (upper panels), as well as the composition of the mRNAs (lower panels) that encode the individual building blocks for these molecules, are shown. A: IgG-derived molecules containing Fc regions. Heterdimerization of different H-chains may be forced by knobs-into-holes technologies, including in common LC (light chain), CrossMab, IgG-dsF, IgG-scFab and Fv2-Fc approaches. Knobs/holes are indicated as complementary black/white circles in the CH3 encoding regions. The dual acting Fab (DAF) which contains evolved Fvs that bind with dual specificity, as well as the DVD (dual variable domain) formats do not require H-chain heterodimerization and hence contain unmodified H-chains. B: Bispecifics without Fc regions. These are generally smaller than Fc-containing entities and are composed of individual scFvs or Fabs of different binding specificities which are fused to each other via linker peptides. Fusion of VH-CH1 and/or L chains to scFvs generate bi- or trivalent Fab-Fv or Fab-Fv2 entities. Fusions of scFvs of different specificities generate rather small scFv-scFv molecules (such as BiTEs). Diabodies, dual-affinity retargeting molecules (DARTs), and TandAbs are generated by modulating the length of the linker peptides to force correct pairing of VH and VL domains (which are not scFvs in a strict sense). These molecules can be additionally stabilized by interchain disulfide bonds (e.g. in DART). Variable regions of different specificities are indicated by different colors (red, blue and green), invariable constant (CH1 and CL) regions are colored grey, and linker peptides that are introduced to connect added entities are colored orange colour. Bispecifics that contain scFvs modules have them frequently additionally stabilized by interchain disulfide bonds between VH and VL. Stars indicate the location of a disulfide bridge. BiTE=Bispecific T-cell engagers; CH1, CH2, CH3=constant region heavy chain domains; CL=constant region light chain; DAF=dual-acting Fab; ds=disulfide; Fab=fragment, antigen binding; Fc=fragment, contant region; Fv=fragment, variable region; HAS=human serum albumin; IgG=immunogobulin G; TandAb=tandem antibody; VH=variable region heavy chain; VL=variable region light chain.
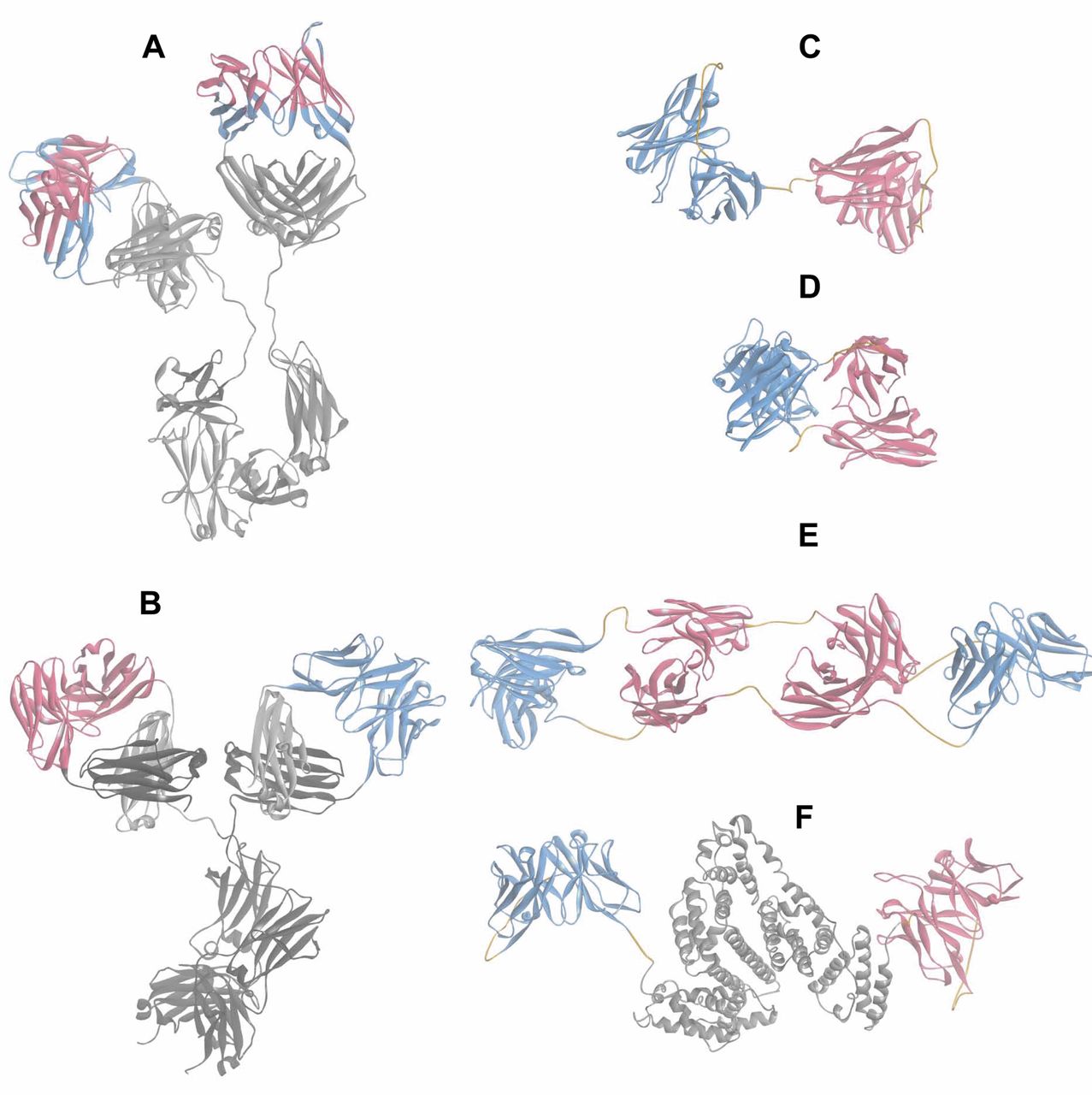
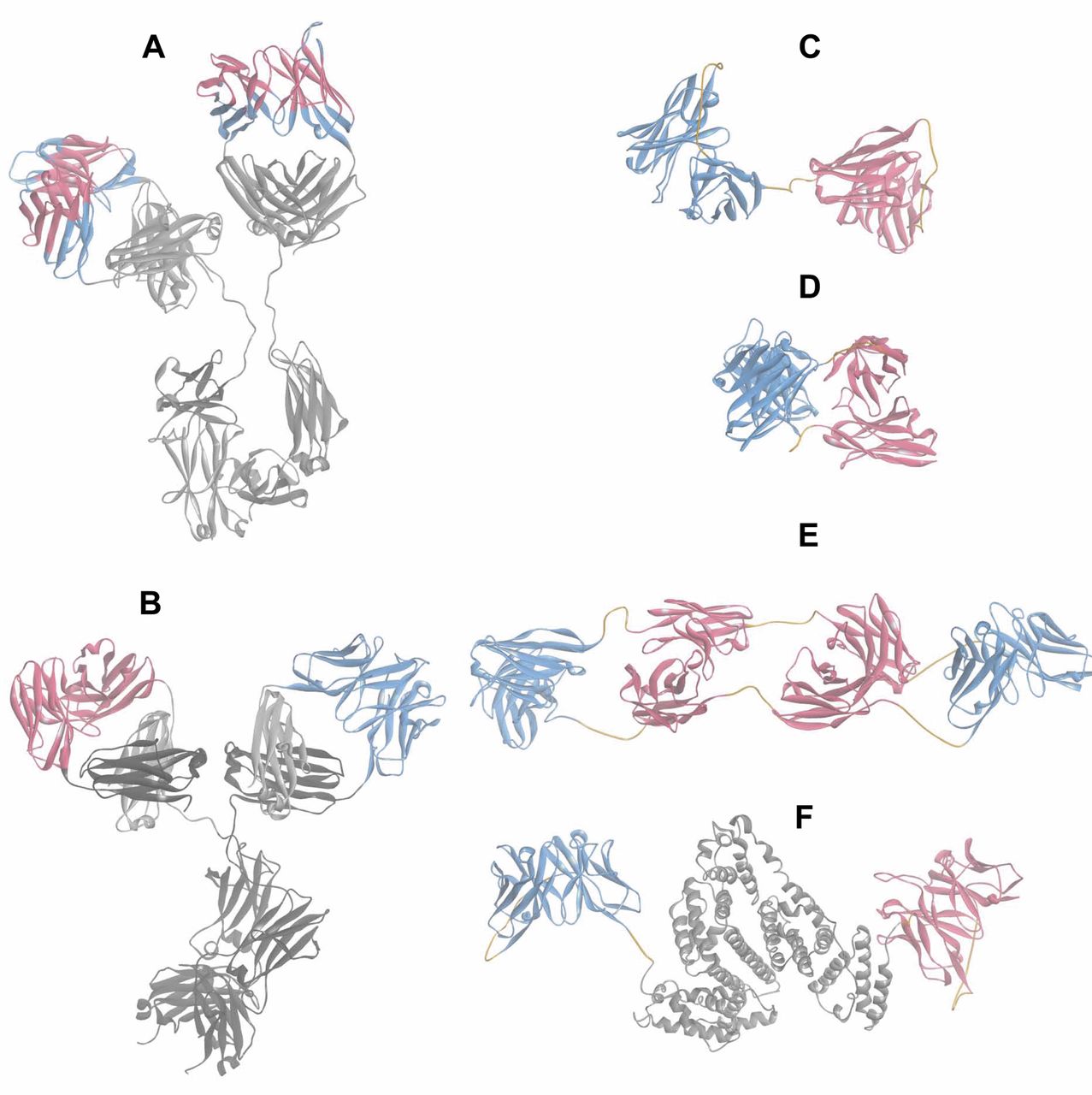
Structure models of mutlispecific proteins. The structure models were generated based on available structure data of domains or entities. Structural data from the Protein DataBank (PDB, Nov. 2011) (135) were assembled and minimized using DiscoveryStudio 31 (136). Ribbon representations are shown to facilitate the domain identification. The color scheme of the models follows that of Figure 1. A: Model of the of the ‘two-in-one’ antibody DAF (HER-2/VEGF) composed of the two Fab structures (pdbcode: 3BE1 and 3BDY, same Fab in complex with their respective antigen) and the Fc portion (pdbcode: 1HZH). B: CrossMab model is composed of the Avastin Fab, blue arm (pdbcode: 1BJ1) and an anti-angiopoietin 2 Fab model fused to the Fc portion (pdbcode: 1HZH); the CH1-CL domain exchange is depicted on the right hand Fab for which the dark grey CH1 domain is now part of the L-chain and the CL (light grey) is inserted in the H-chain. C: The Blinatumomab BiTE model is based on the crystal structure of the anti-CD3 Fv (pdbcode: 1SY6), shown in blue, and a model of the anti-CD19 Fv, shown in red; the linkers within the scFv and between them are displayed in orange. D: A structure of a diabody (pdbcode: 1MOE). E: A TandAb structure of two anti-CD16A Fvs and two anti-CD30 Fvs. Variable domains are connected by (G2S)3 linkers highlighted in orange. F: Structural model of MM-111 composed of scFv HER2 (red ribbon), an AAS linker, human serum albumin (pdbcode 3 JQZ), grey, an AAAL linker and scFv HER3 shown as a blue ribbon. AAAL=Alanine and leucine-based linker; AAS=alanine and serine-based linker; CD=cluster of differentiation; CL=constant region light chain; DAF=dual-acting Fab; Fab=fragment, antigen binding; Fv=fragment, variable region; HER2=human epidermal growth factor 2 receptor; VEGF=vascular endothelial growth factor.
BiTEs have been generated for several tumor-associated target molecules such as CD19, EpCAM, HER2, CEA, ephrin A2 (EphA2), CD33 and melanoma-associated chondroitin sulfate proteoglycan (MCSP) (52). They induce target cell lysis in cell culture assays at concentrations ranging between 10 pg/ml and 10 ng/ml (56), without the need for T-cell co-stimulatory signals. As was shown by laser scanning confocal microscopy, the polyclonal T-cell engagement is associated with the induction of cytolytic T-cell synapses also on MHC-class I negative tumor cells (57). Video-assisted microscopy has shown that BiTEs can elicit multiple rounds of target cell lysis by T-cells at very low E/T cell ratios, and that activation of T-cells occurs only in the presence of target cells (58). T-Cells kill cancer cells by membrane perforation and subsequent induction of apoptosis using granzyme B and other granzymes (59) resulting in activation of caspases 3 and 12. BiTEs exhibit 100-to 10,000-fold higher efficacy in tumor cell lysis compared to other CD3-bispecific formats (60). They are stable and homogenous at the protein level, not glycosylated and can easily be produced as soluble proteins.
In vivo, treatment of non-obese (NOD) severe combined immune-deficient (SCID) mice with submicrogram doses of anti-CD19/anti-CD3 BiTE (Blinitumomab) per mouse was able to prevent growth of subcutaneous (s.c.) human B lymphoma xenografts and to cure animals when administered at an early stage of disease in the presence of human lymphocyte effector cells (61). Efficacy was dependent on the dose, the E/T ratio and the time between tumor inoculation and administration of the BiTE. The human T-lymphocytes did not express T-cell activation markers. In vivo evaluation of the anti-EpCAM/anti-CD3 BiTE was performed by s.c. injection of SW480 colon carcinoma cells mixed at a 1:1 ratio with unstimulated human PBMCs into NOD/SCID mice and treatment with five daily injections of 100 ng per mouse (62). Tumor growth was prevented when treatment was started on the day of tumor inoculation. Established tumors were eradicated when BiTEs were administered up to eight days after tumor cell injection. In a different model, tumor tissue from patients with ovarian cancer was implanted into NOD/SCID mice. Treatment of established tumors with 5 μg doses daily led to significant reduction and in some cases, eradication of tumors (62). The relatively short half-life of BiTEs, in the order of several hours, requires an appropriate dosing schedule and formulation to ensure continuous activation of T-cells against target cells.
For Blinatumomab (anti-CD19/anti-CD3) (53), T-cell activation and B-cell depletion was shown in chimpanzees (63). Polyclonal T-cell activation was transient and repeatable. In the clinc, Blinatumomab depleted chronic lymphocytic leukemia (CLL) B-cells in presence of autologous T-cells in 22 out of 25 cases, also in patients pretreated with different chemotherapy regimens (64). Blinatumomab mediated tumor regression in patients with non-Hodgkin’s lymphoma (NHL) (65). Doses as low as 0.005 mg/m2 led to elimination of tumor cells in the blood, partial and complete tumor regressions were observed at a dose of 0.015 mg/m2, and tumor regression was seen in all seven patients treated with 0.06 mg/m2. In addition, clearance of tumor cells from bone marrow and liver was observed. Another BiTE (anti-EpCAM/anti-CD3) is currently under clinical investigation in patients with solid tumors.
Further Formats for Recruitment of Immune Effector Cells
TandAbs (Figures 1B and 2E) were developed for the recruitment of immune effector cells to tumors (66). TandAbs are tetravalent, bispecific fusion proteins, generated by connecting four antibody variable sequences (VHA-L1-VLB-L2-VHB-L3-VLA) with 12 amino acid linkers, which do not allow pairing of the individual domains of the same chain. As a consequence, the V regions are forced to pair with the complementary domains of another identical chain, thus forming homodimeric proteins. Despite the absence of a covalent bond connecting the monomers, the homodimers are stable and do not revert back to the monomeric form. TandAbs possess four binding sites, extended PK properties compared with other, smaller bispecific formats and are, with a molecular weight of 105-110 kDa, much larger than the cut-off size for glomerular filtration.
TandAbs do not possess an Fc moiety. AFM11 (CD19xCD3), a T-cell recruiting molecule is in preclinical development for indications such as NHL, B-cell-CLL and other hematological malignancies. AFM11 exhibits potent cytotoxic activity in vitro, with (EC50s) in the low- and subpicomolar range (66). AFM13 (CD30xCD16A) has entered clinical studies for the treatment of patients with recurrent Hodgkin’s lymphoma. Preclinical studies have shown that AFM13 efficiently can recruit NK cells and macrophages (67).
Yet another format for recruitment of immune effector cells is that of bispecific hexavalent Abs [HexAbs: IgG(Fab)4]. The dock-and-lock method was applied to generate multivalent, multispecific structures that include monospecific and bispecific HexAbs, each comprising a pair of stabilized dimers of Fabs linked to a full IgG at the carboxy-termini of both H-chain subunits, resulting in a hexavalent antibody constructed around a shared Fc moiety targeting CD20/CD22 or CD20/CD74 (68, 69). Potent cytotoxicity induced by the CD20/CD74 HexAb was demonstrated for mantle cell lymphoma (MCL) cells, with loss of mitochondrial membrane potential, production of oxygen species, actin reorganization, and down-regulation of phosphorylated AKT (pAKT) and B-cell leukemia 2-associated X protein (Bcl-xL). A further format to generate molecules redirecting immune effector cells to tumors make use of human IgG1 hinge-CH2-CH3 as a scaffold fused to scFv for tumor targeting and recruitment of immune effector cells (70). Specifically, anti-leukemia stem cell antigen CD123 scFv was fused to the N-terminus, and anti-CD3 scFv to the C-terminus of such molecules (70). The resulting fusion protein is a homodimer which binds to CD123 leukemia cells with a Kd of 10–11 M, one to two orders of magnitude stronger than that of most conventionel antibodies. BLf can kill CD123 positive cells in a T-cell-mediated manner at low picomolar concentrations and E/T cell ratios as low as 2. Inclusion of human IgG-Fc region should result in conferring a half-life in serum similar to antibodies, and the molecular weight of 140 kDa should prevent clearance via glomerular filtration. Placement of the anti-CD3 scFv at the C-terminus reduced binding affinity to cell surface CD3 by two orders of magnitude as compared to an N-terminally attached anti-CD3 scFv, with a Kd in the range of 10–8 M. However, this affinity is still 10- to 40-fold higher than that of the monomeric scFv of BiTEs, with the consequence of stimulating peripheral blood lymphocytes (PBL) to secrete IFNγ in the absence of target cells. Mutations will probaly have to be introduced into the anti-CD3 scFv to select constructs with reduced binding affinity.
Bivalent binding of antibodies to CD3 can induce a massive cytokine release (71) and low affinity Fcγ receptor (FcγRIIIa) and CD32a (FcγRIIa) on immune effector cells mediate killing by monovalently binding to the Fc region. Upon engagement with the target cell, receptors are cross-linked and clustered on the cell surface of the T-cells, leading to activation (71). Therefore an antibody format was generated which enables bivalent binding to the tumor antigens and monovalent binding to antigens on effector cells (72). Monovalent binding to a second antigen is achieved in a full-length antibody context. A variant pair of H-chains to promote heterodimerization of the Fc part was genetically engineered allowing fusion of VH and VL domains to the C-terminus of the two H-chains. Fusion proteins with anti-CD16 or anti-CD3 scFv based on trastuzumab and antibody anti-HM1,24 (73) were evaluated. Potent and selective cytotoxic effects were demonstrated for both anti-CD16 and anti-CD3 scFvs. The simultaneous engagement of antigens with different valencies in the context of a bispecific antibody with a favorable half-life can probably be extrapolated to many other experimental systems.
For the visualization of antibody-induced T-cell responses, an elegant method has been described (74). Green fluorescent protein (GFP) flanked by two novel helical linker elements was inserted into scFv anti-CD33/anti-CD3 and anti-prostate stem cell antigen (PSCA)/anti-CD3 bispecific tandem scFvs. It was shown that both GFP-fusion antibodies were concentrated in the artificial immunological synapses formed between T-cells and tumor cells.
Single-chain Triple Antibodies
Single-chain triple antibodies (sctbs) consist of three scFvs which are connected in tandem by linkers (75). The objectives for the design of sctbs were to increase the avidity of the molecule for one of the targets as well as generating a molecular mass beyond the kidney exclusion limit of 65 kDa. One such sctb consisting of two distal scFvs specific for CD19 and a central scFv specific for CD16 (FcγRIII) was evaluated in vivo. (75). The disulfide bridge-containing scFvs had a serum half-life in mice of 4 h. Thus, the plasma-half life was improved two-fold, while the avidity for CD19 was increased three-fold (13 versus 42 nM), with equal affinity for CD16 (58 nM). In an ADCC assay,10- to 40-fold lower concentrations of the sctb were required for lysis of leukemia cells by human PBMCs as compared to a dual tandemly repeated fusion protein. The reasons for the improved ADCC are unclear. More effective formation of an immunological synapse and/or improved binding to CD19 due to the second CD19 binding moiety are possible explanations. A similar fusion protein designed for the treatment of acute myeloid leukemia (AML) was evaluated (76). The corresponding sctb simultaneously targets CD123 (IL3 receptor α chain), CD33, and CD16 (FcγR III), only the scFv targeting CD16 contains a disulfide bridge. This sctb was compared to a bispecific construct lacking an scFv targeting CD123. An improvement of Kd from 25 nM to 6 nM was noted, probably due to an avidity effect caused by the addition of the scFv directed against CD123. Both the bi- and the trispecific fusion proteins bound to CD123/CD33-expressing AML-derived cell lines MOLM-13 and THP-1. However, a significantly higher target cell lysis was observed for the sctb as compared to the bispecific molecule, which was likely due to the higher combined antigen density. For targeting of mixed lineage leukemia cells, an sctb targeting CD19, CD33 and CD16 was constructed (77). The CD16 and CD19-specific scFvs were stabilized by a disulfide bridge. The sctb 33-ds16-ds19 mediated lysis of target cells more effectively than 33-ds16 and ds16-ds19. Unfortunately, in vivo data are not yet available. However, formation of aggregates might be a critical issue with this type of molecules and remains to be resolved. The hypothesis that dual specificity might confer the ability for selective binding to double-positive as compared to single-positive cells was tested (78). To this end, sctb HLA-ds16-hu19 was produced and characterized. The molecule consists of three scFv antibody fragments in a single polypeptide chain, two peripheral ones specific for the human histocompatibility antigen HLA-DR and human B-cell antigen CD19, and a central one directed against human low-affinity Fc-receptor (FcγRIII, CD16). The CD16-binding modules are disulfide-stabilized. This sctb was compared with corresponding bispecific constructs and shown to target preferentially double-positive Raji cells (HLA-DR and CD19). Preferential targeting of leukemia stem cells might be a potential application of this technology.
Cytotoxic Bispecific Antibodies
TNFα is a multipotent cytokine produced mainly by activated macrophages. TNFα has potent antiproliferative, antitumoral and antiangiogenic activity (79, 80). It induces disruption of the vasculature by involvement of integrin αvβ3 (81). Clinical studies have revealed a significantly lower maximal tolerated dose in humans (about 10 μg/kg) as compared to mice (400 μg/kg) (82). Targeting of TNFα to the tumor might therefore improve the therapeutic index. Antibody Fab fragments specific for CEA and TNFα were connected via stable thioether linkage. Targeting of such molecules to human colon carcinoma xenografts was shown with 125-I-labeled bispecific antibodies (82). In vitro clonogenic assays suggested an additive or supra-additive interaction between TNFα and ionizing radiation. In addition, oxidative damage produced by TNFα may enhance the cellular damage induced by ionizing radiation (83). In a BxPC3 pancreatic xenograft model, bispecific anti-CEA/anti-TNFα molecules augmented tumor sensitivity to radiation as reflected by an increase of medium survival times (84); and in a syngeneic CEA-transfected colon carcinoma model (MC38), the bispecific antibody mediated radiation curability (85).
Another strategy is the combination of tumor-cell targeting and activation of a death receptor by bispecific antibodies. One well-known death receptor is CD95 (APO-1, FasR). The starting point for targeting of activating CD95 on tumors with bispecific antibodies was the finding of liver failure in mice treated with an agonistic CD95 antibody (86). A bispecific antibody directed against CD20 and CD95 was shown to induce cell in death in lymphoma cells by cross-linking of the CD20 target antigen and CD95 through bicellular binding (87). Subsequently, it was shown that recombinant bispecific tandem scFv connected by a flexible linker were as effective as a chemically hybridized F(ab)2 fragment with identical specificities (88). In addition, it was demonstrated that a bispecific antibody directed against CD95 and NGA-2 induced CD95-mediated apoptosis of glioblastoma cells, whereas a bispecific antibody directed against CD95 and EGFR failed to induce apoptosis of these cells (88). In the case of bicellular binding, effective cross-linking of CD95 is mediated by this bispecific antibody and correlates with induction of apoptosis; in the case of unicellular binding (cis binding), apoptosis is not induced. In order to facilitate bicellular binding in a trans configuration, a bispecific antibody directed against CD95 and extracellular matrix protein tenascin was designed and shown to be the most effective of all bispecific antibodies, simultaneously targeting CD95 and a tumor-associated antigen. The efficacy of these bispecific antibodies was enhanced by various cytotoxic agents.
Another strategy to induce cell death using bispecific antibodies is the simultaneous engagement of two death receptors such as TNF-related apoptosis-inducing ligand receptor-2 (TRAIL-R) and lymphotoxin-beta receptor (LTβR), which are often coexpressed on the cell surface of epithelial tumors (89-91). A tetravalent, stably engineered antibody was generated by genetically fusing an anti-LTβR scFv to either the N- or C-terminus of the H-chain of a full-length monoclonal antibody to TRAIL-R2 IgG1. These tetravalent, bispecific antibodies exhibited enhanced antitumor efficacy as compared to the mere combination of the corresponding parental antibodies in a subset of cell lines in vitro. This may be due to cross-linking of the receptors, an event which does not occur by combining individual antibodies. No inhibition of proliferation of human umbilical vein endothelial cells (HUVECs) was observed, despite the presence of both receptors on these cells. Finally, established tumors were treated in in vivo models (WiDr, colon cancer and MDA-MB 231, breast cancer). While individual antibodies, and their combination did not result in efficacy, 60% tumor growth inhibition was observed in WiDr xenografts treated with the N- and C-terminal bispecific antibodies, whereas 50% tumor growth inhibition was observed for the C-terminal antibody treatment of MBA-MB 231 xenografts. No tumor growth inhibition was seen using the N-terminal antibody fusion. Half-life times of 248 and 362 h were observed in mice for the N- and C-terminal bispecific antibodies, respectively.
Bispecific Antibodies Directed against Receptor-tyrosine Kinases
In humans, four membrane tyrosine kinases belonging to the HER receptor family are known: HER1/EGFR, HER2, HER3 and HER4. The ligands are EGF-related growth factors which induce intrinsic receptor kinase activity leading to stimulation of intracellular signaling cascades after binding (92). In this system, HER2 acts as a co-receptor and is overexpressed in up to 30% of breast carcinomas due to gene amplification, and overexpression correlates with shorter time to relapse and shorter overall survival (93). Preferential phosphorylation of HER3, but not EGFR, was observed in HER2-amplified breast cancer tissues (94). HER2 requires heterodimerization with HER3 to drive breast cancer cell proliferation and is essential for the transformation process (95). Trastuzumab, lapatinib and pertuzumab are poor inhibitors of ligand-induced HER3 activation, as shown by inhibition of phospho-HER3 generation in a panel of cancer cell lines (96). However it was demonstrated that targeting HER2 and HER3 with a bispecific scFv was able to induce cell-cycle arrest in cancer cell lines with amplified HER2 (97). In vivo proof-of-concept experiments were not shown in this study and might be difficult to perform due to the serum half-life inherent to the antibody design. In order to improve PK properties, MM-111, a bispecific single polypeptide fusion protein of two antagonistic scFv directed against HER2 and HER3 and linked to modified human serum albumin (HSA) was developed (96). A modified version of HSA was placed between the scFvs of MM-111 with short peptide linkers connecting the scFvs to the amino- and carboxy-termini of HSA, respectively (Figures 1B and 2E). MM-111 can be produced on a large scale, forms a trimeric complex with HER2 and HER3, and potently inhibits the phosphoinosotol-3 kinase (PI3K) pathway and proliferation of HER2 overexpressing tumor cells. In vivo efficacy of MM-111 was investigated in nine xenograft models and its efficacy was found to depend on HER2 expression. In the BT 474-M3 in vivo model, improved efficacy was noted by combination of MM-111 with lapatinib or with trastuzumab. Half-life times of 17 to 23 h were observed in nude mice, which compares favorably with typical half-life times of ca. 5 h for scFv tandem bispecific molecules (98).
Extensive cross-talk between HER family members has been observed. Therefore, addressing two HER family members simultaneously might increase therapeutic efficacy (99-100). In this context, a bispecific HER2/EGFR affibody was generated (101). Affibodies are affinity proteins that have been developed by engineering of a 58 amino acid residue Z-domain scaffold derived from Staphylococcus aureus protein A (102). Simultaneous targeting of HER2 and EGFR was demonstrated.
Based on the observation that complete inhibition of mitogen-activated protein kinase (MAPK) and AKT signaling was achieved when EGFR and HER3 were blocked simultaneously, a so-called two-in-one antibody, MEHD7945A (Figure 1A), which can bind both to EGFR and HER3 (affinities of 1.9 and 0.4 nM, respectively) was generated (18). MEHD7945A (Figure 1A) is based on a high-affinity antagonistic EGFR antibody and was derived from an antibody library with diversity restricted to the H-chain complementarity determining regions (CDRs) and a subsequent library of variants with mutations in the L-chain CDRs resulting in molecules binding to HER3 while maintaining their specificity for EGFR. As a human IgG1, MEHD7945A inhibited EGFR and HER3 signaling and was able to exert immune effector functions. MEHD7945A was as efficacious in vivo as the corresponding parental EGFR antibody in the non-small cell lung carcinoma (NSCLC) model NCI-H292 in which tumor growth is dependent on EGFR expression and signaling. When compared to the corresponding HER3-monospecific antibody, MEHD7945A was also similarly efficacious in a model depending on HER3 activation (BxPC3). Importantly, in models in which both receptors contribute to tumor growth, MEHD7945A possessed superior efficacy over the combination of the monospecific antibodies, e.g. in the Calu3 (NSCLC) and FaDu [head and neck squamous cell carcinoma (HNSCC)] xenograft models. Synergy with gemcitabine was shown in vitro in several NSCLC cell lines and in vivo in the NSCLC xenograft model NCI-1975. In cynomolgus monkeys, MEHD7945A induced less severe skin toxicity when compared with cetuximab at comparable drug exposures.
EGFR and insulin-like growth factor receptor 1 (IGF1-R) signaling is deregulated in many types of cancer, suggesting that the simultaneous inhibition of these receptors might be a valid therapeutic strategy. A12scFv (anti-IGF1-R) or 11F8scFv (anti-EGFR) in one version were fused to the N-termini of CL and CH1 domains, or vice versa, of an IgG1 scaffold resulting in a tetravalent, bispecific antibody (103). Improved inhibition of signaling as compared to that of the combination of the individual antibodies was shown in BxPC3 cells in vitro, but the combination of the antibodies was more effective in vivo.
An IgG-like tetravalent bispecific di-diabody targeting IGF1-R and EGFR was constructed (104). Functionally, blocking of the signaling pathways as well as IGF1-R internalization and degradation, was observed in vitro. In in vivo nude mouse xenograft models, similar (HT29, colorectal cancer), or slightly lower (BxPC3, pancreatic cancer) efficacy was observed treating 200-300 mm3 tumors as compared to that of the combination of the parental antibodies at the same dose. Another tetravalent IgG1 format with dual targeting capacity for IGF1-R and EGFR was also explored (13). Here, disulfide bridge-stabilized scFv domains of the EGFR antibody GA201 (105) were fused via Gly-Ser connectors to the C-termini of the H-chain (XGFR2) or L-chain (XGFR4), or to the N-termini of the L-chain (XGFR5) or H-chain (XGFR3), of the parental IGF1-R antibody R1507 (106, 107). In vitro, XGFR2, XGFR3 and XGFR4 effectively inhibited IGF-1R transphosphorylation as well as 3D proliferation of H322M and H460M2 tumor cells, and induced down-modulation of EGFR and IGF-1R. ADCC was maintained; however, a format dependency of ADCC and receptor down-modulation was noted. In vivo evaluation of XGFR2 and XGFR4 in BxPC3 and H460M2 xenograft models showed potent antitumor efficacy comparable to that of the combination of the parental, monospecific IGF-1R and EGFR antibodies. Interestingly, while IGF1-R down-modulation was maintained, EGFR down-modulation was enhanced as compared to use of the monospecific GA201 antibody. At present, this phenomenon is being investigated in more detail.
Targeting non-overlapping epitopes on hIGF-II using a bispecific antibody (m660) was described as a novel type of candidate cancer therapeutic (108). M660 blocked the interaction of hIGF-II with IGF-1R on MCF-7 breast cancer cells and modulated hIGF-II mediated IGF-1R and insulin receptor phosphorylation and cell growth. In addition, the formation of oligomeric antibody ligand complexes that bound to cells expressing Fc gamma receptors (FcγRs) with high avidity, as well as phagocytosis by macrophage-like U937 cells of such complexes, was shown. This happened more efficiently than with the monospecific antibody hIGF-II complexes. A combination of the monospecific antibodies displayed similar properties to the bispecific one. Of concern is the possibility of side-effects due to immune complex formation because this is a common problem in several autoimmune diseases (109). In vivo studies will have to reveal whether m660, or the combination of the corresponding monospecific antibodies can be potential candidates for IGF-related cancer therapeutics.
Bispecific Antibodies Interfering with Tumor Angiogenesis
Formation of blood vessels is controlled by several angiogenic factors and their respective receptors within a complex and highly interconnected network. However, inhibition of one particular pathway can lead to the activation of alternative pathways, resulting in resistance to antiangiogenic treatment (110, 111).
Vascular endothelial growth factor (VEGF) promotes angiogenesis mainly through interaction with VEGF receptor 1 (VEGFR1) and VEGFR2. Thus, blocking both receptors simultaneously with a bispecific antibody might result in improved efficacy regarding inhibition of tumor angiogenesis. Indeed, a corresponding diabody blocked the interaction of VEGF-A with both receptors and also inhibited binding of placental growth factor to VEGFR1. The diabody inhibited VEGF-A-induced mitogenesis of human endothelial cells, as well as VEGF-A-induced migration of human leukemia cells (112); however, no in vivo efficacy data were reported.
Osteopontin, an acidic, phosphorylated glycoprotein can bind to some CD44 splice variants and several integrins (113) and can promote angiogenesis by inducing PI3K/AKT and extracellular signal-regulated kinase 1/2 (ERK1/2) signaling in endothelial cells (114). A bispecific antibody based on the antibody to VEGF-A, bevacizumab, and antibody to osteopontin, hu1A12, was produced (115). The VH and VL domains of bevacizumab were fused to the N- and C-termini of the H- and L-chains of hu1A12, respectively. Compared to the combination of parental antibodies, the bispecific antibody was significantly more efficacious in suppressing tumor growth and angiogenesis in the liver cancer xenograft model HCCLM3 in nude mice at equal doses.
Another strategy for the inhibition of tumor angiogenesis is the simultaneous inhibition of VEGF-A-mediated signaling in endothelial cells and the coverage of newly formed endothelial cells with pericytes (116). Platelet-derived growth factor (PDGF) isoforms and their corresponding receptors play important roles in tumorigenesis through stimulation of tumor cell growth and promotion of angiogenesis by vessel formation and recruitment of pericytes. Signaling-inhibitory antibodies directed against murine PDGF receptor β (PDGFRβ) (1B3) and murine VEGFR2 (DC101) were evaluated in the pancreatic tumor BxPC3 xenograft model (117). Thus, while combination treatment with both antibodies caused tumor regression in 58% of treated mice, only 18% of mice treated with DC101 monotherapy showed tumor regression and no, or only very moderate growth inhibition was observed in mice treated with 1B3 alone at the corresponding doses.
A tetravalent, bispecific antibody was engineered by fusing a scFv directed against mouse PDGFRα to the N-terminus of the L-chain of an antibody directed against mouse VEGFR2. Antigen binding and signaling inhibitory activity of the parental antibodies were retained (118).
Another strategy to target VEGFR2 and PDGFRβ is the combination of a fully human anti-PDGFRβ antibody (IMC-2C5), crossreactive with mouse and human PDGFRβ, with DC101, an antibody directed against murine VEGFR2. In BxPC3, NCI-460 and HCT-116 xenograft models, IMC-2C5 possessed very moderate antitumor activity, however, its combination with DC101 resulted in significantly enhanced antitumor activity, also in comparison to DC101 (119).
In another approach targeting tumor angiogenesis, VEGF-A and PDGFRβ were selected as targets (120). ScFv antibody fragments targeting human PDGFRβ and human VEGF-A were fused to both termini of human Fc to generate bispecific tetravalent molecules. In vitro, the scFv-Fc-scFv fusion protein inhibited VEGF-A-induced proliferation of HUVECs and blocked PDGF-BB isoform-induced proliferation of human brain vascular pericytes. In SCID mice, a half-life of 460 h was observed. Prophylactic and therapeutic treatment of A673 rhabdomyosarcoma xenograft tumors inhibited tumor growth with an efficacy similar to that seen with bevacizumab.
A different approach to inhibit tumor-related angiogenesis is the simultaneous inhibition of VEGF-A and angiopoietin-2 (ANG2), a ligand of tunica internal endothelial kinase 2 (TIE2). It was shown that a chimeric decoy receptor, double anti-angiogenic protein (DAAP), which can bind both to VEGF-A and angiopoietin, was more potent in several in vivo tumor models as compared to treatment with single agents (121). DAAP is composed of the two Ig-like domains and the three EGF-like domains of TIE2, the second Ig-like domain of VEGFR1 and the Fc-portion of IgG1. Another variation of this theme is the targeting of VEGF-A and ANG2 using a bispecific antibody in the CrossMab format (Figure 1A and 2B) as outlined previously (24, 25). In Colo205 xenograft tumors in SCID mice, bevacizumab (10 mg/kg, i.p.), antibody to ANG2 LC06 (10 mg/kg), the combination of the two mAbs (10 mg/each) and ANG2-VEGF CrossMab CH1CL (20 mg/kg) were compared. The bispecific CrossMab CH1CL administered at the equivalent dose in terms of binding sites mediated the best tumor growth inhibition of 92%.
A two-in-one antibody recognizing two clinically validated tumor targets, HER2 and VEGF-A, was produced (Figures 1A and 2A) (19). The HER2 antibody trastuzumab with randomly mutagenized CDRs of the antibody L-chain was expressed as a phage library, and the library was screened for binding to both VEGF-A and HER2. In this way, an antibody with improved affinity for both targets (Kd values of 3 nM and 0.2 nM, respectively) was identified. The binding surface of the antigens overlapped; however, distinct amino acids also contributed to the binding affinities for VEGF-A and HER2. Binding to VEGF-A was primarily determined by the L-chain, while the H-chain mediated binding to HER2. The capability of the L-chain CDRs to rearrange appeared to be necessary for the dual specificity of the selected antibody. In vivo, the dual-specificity antibody was evaluated in the anti-VEGF-responsive Colo205 model, and the trastuzumab-sensitive BT474M model. The dual specificity antibody was as effective as trastuzumab and anti-VEGF treatment administered separately or in combination in these models. In vivo evaluation of this molecule in trastuzumab- and anti-VEGF-responsive models treatment is ongoing.
Bispecific Antibodies for Targeting of Intracellular Antigens
Some of autoimmune disease-related antibodies were found to be localized in the nucleus and to bind to DNA (122). In this context, mAb 3E10, which is directed against double-stranded DNA and localizes to the cell nucleus, was identified (123). A single point mutation resulting in the conversion of aspartic acid to asparagine in residue 31 (VH31Asn) markedly increased its capability to penetrate the plasma membrane of living cells. Complexes between recombinant E310-derived Fab fragments and alkaline phosphatase-conjugated goat antibodies to mouse kappa chains, as well as a fusion protein between the corresponding scFv and GFP, were shown to be able to penetrate the plasma membrane of mammalian cells and subsequently localize to the nucleus (124). Functional restoration of mutated proapoptotic genes by delivery of fusion proteins between 3E10-derived Fv and wild-type proapoptotic gene products in order to induce apoptosis is one possible application for this type of molecule. Delivery of a p53 3E10 Fv fusion protein was shown to inhibit experimental liver metastases after i.v. injection of colon cancer cells and subsequent splenic injection of the fusion proteins (125). Similarly, a fusion protein between 3E10 scFv and nuclear transcription factor forkhead box P3 (FOXP3) induced apoptosis of several types of tumor cells in a dose-dependent manner and reduced the metastasis score in a syngeneic model of colon cancer (126). In addition, bispecific antibodies based on 3E10 scFv have the potential to interfere with oncogenic intracellular protein protein interactions, and thus to restore defective tumor suppressor gene function. For example, the antibody PAb421 was shown to reconstitute transcriptional activity of several p53 mutants by restoring the interaction of the C-terminus of p53 with DNA (127). A bispecific singe-chain construct consisting of 3E10 Fv and PAb421 induced cell death in a dose-dependent manner of SW480 colon carcinoma cells expressing a mutated version of p53 (His-273, Ser-309). In various tumor types, mouse double minute 2 (MDM2), an E3 ubiquitin ligase, is overexpressed and mediates down-regulation of the expression of p53. Inhibition of the interaction between p53 and MDM2 with small molecules was shown to induce tumor cell apoptosis (128). mAb 365 which binds to critical residues at the N-terminus of MDM2 that are required for the interaction with p53, was previously identified (129, 130). A bispecific molecule consisting of 3E10 scFv and 3G5 scFv was shown to retain cell penetrating and MDM2-binding activity, to increase p53 levels in xenografts, and to inhibit growth of MDM2-dependent melanoma cell lines and xenografts (131). Accordingly, molecules such as transcription factors, DNA repair proteins, oncogenes and other tumor-relevant gene products, which are presently considered to be inaccessible for small molecule compounds, could be targeted with such bispecific molecules.
Conclusion
From a technical point of view, depending on the format of the multispecific molecules, optimization of efficacy, PK and tumor penetration, as well as ease of production, are ongoing processes. One of the attractive features of multispecific binders is their capability to target immune effector cells to the tumor. Another approach would be to combine tumor-targeting and abrogation of the inhibition of T-cells, dendritic cells and macrophages (132, 133). Recruitment of immune effector cells making use of a stromal target, such as fibroblast activation protein, is another potential of this technology (134). Co-targeting of stromal cells and tumor cells is another obvious possibility for improvement of efficacy of tumor treatment. The simultaneous engagement of death receptors offers many possibilities for exploitation by multi-specific antibody-derived formats. Simultaneous modulation of plasma membrane-associated receptor signaling offers prospects of more efficient therapeutic intervention and prevention of resistance to therapy. Given the plethora of combinatorial options, appropriate target validation will be instrumental in defining the most promising combinations.
- Received December 21, 2012.
- Revision received January 23, 2013.
- Accepted January 23, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- Formats
- Initial Efforts to Recruit Immune Effector Cells Using Bispecific Antibodies
- Targeting CD28 for Recruitment of Immune Effector Cells
- Trifunctional Bispecific Antibodies
- Bipecific T-Cell Engagers (BiTEs)
- Further Formats for Recruitment of Immune Effector Cells
- Single-chain Triple Antibodies
- Cytotoxic Bispecific Antibodies
- Bispecific Antibodies Directed against Receptor-tyrosine Kinases
- Bispecific Antibodies Interfering with Tumor Angiogenesis
- Bispecific Antibodies for Targeting of Intracellular Antigens
- Conclusion
- References
- Figures & Data
- Info & Metrics