


Abstract
Background: When the functions of a protein serve a useful survival and unique purpose, the selective pressures of evolutionary laws of nature conserve the DNA sequences encoding such proteins. In many instances, the conservation of these sequences has occurred since the inception of life on earth to the present in phylogenetically related species. The unique function(s) of metallopanstimulin (MPS-1/RPS27) ribosomal protein (RP) and a limited number of other RPs, in growth regulation, and viral infection is further documented here. Based on the correlation of information concerning Genome Context Analysis, and new information presented here, the author proposes that neutralization or elimination of ribosomal MPS-1/S27 DNA, mRNA or translated protein in eukaryote cells, initiated in the process of chemical, viral or radiation carcinogenesis can result in control of most carcinogenic processes by selective elimination of transformed cells which display overexpression of RPMPS-1/S27, and/or non-lethal pathogenic mutations of RPMPS-1/S27 gene. Recently, critical interactions were reported between RPMPS-1/S27 and p53 induced by DNA damage such as ionizing radiation, or chemotherapy drugs, that result in the activation of p53 which in turn represses RPMPS-1/S27 actions. Thus, p53, RPS27L, and RPS27/MPS-1) regulate growth and survival. Materials and Methods: Antivirals were tested in virus-infected cells using: cell culture, cytotoxicity assays, apoptosis, defined virus strains, cloned cells, and RT-PCR. Purity of antivirals was validated by mass spectroscopy (MS). Disruption of zinc finger peptides (ZFPs), by these agents was determined by NMR. Results: The data presented here indicates that anti-ZFP agents can potentially be used to prevent and control viral infections by disrupting viral ZFP motifs. Different DNA/RNA virus-infected cells exposed to the antivirals resulted in distruption of both RPMPS-1/S27 and essential viral ZFPs. Picolinic acid (PA) and fusaric acid (FU) were tested and have been shown to have both antiviral and preventive antiviral activities which have been consistently shown to be mediated, at least in part, via interacting with RPMPS-1/S27. The same antiviral agents simultaneously disrupt essential viral ZFPs. Both antiviral events on ZFPs render the pathogenic virus inactive. Conclusion: It is demonstrated here that PA and FU exhibit antiviral activity towards several DNA and RNA viruses of human and animal importance. Illustrative evidence of the mechanism of action was obtained via MS, NMR, and molecular modeling that PA and, more potently, FU, bind to a particular site of the viral ZFPs. Similarly, it was previously shown by MS, NMR and molecular modeling with RPMPS-1/S27 that PA and FU disrupts the function of this RP protein, preventing viral replication by formation of ternary complexes. This work is consistent with a critical role of RPMPS-1/S27 in the life cycle of various viruses and shows that disruption of viral ZFPs is potentially important to control and prevent deathly viral diseases.
RPMPS-1, also known as metallopanstimulin-1 (MPS-1/RPSP27) is a ribosomal protein that is over expressed in many cancer cell lines, in most of the cancer tissues obtained directly from the patient tumors, and is increased in the blood of patients with cancer at early stages of carcinogenesis (1-10). Previous work reported that this 9.8 kDa protein plays a role in DNA repair, contains one zinc finger in its monomeric state and is also involved in carcinogenesis (1-10). Initially, its expression was found to be increased in over forty different types of malignancies (11). Furthermore, Fernandez-Pol et al. demonstrated the potential of MPS-1 as a tumor marker in the blood for numerous cancers and carcinogenic processes (8, 9, 11).
MPS-1 is a ribosomal protein with extra-ribosomal functions such as DNA repair and transcription (1, 10). The results obtained with Archaea and the functions of MPS-1 and other ribosomal proteins indicate a previously unrecognized regulatory network coupling DNA replication, DNA repair, DNA transcription, translation and biogenesis of ribosomes that exist in both Archaea and Eukarya (1, 6, 7).
MPS-1 was first identified, cloned, sequence, and associated with carcinogenesis by Fernandez-Pol (6), as a key ribosomal protein involved in signal transduction (1, 3, 7). This 9.8 kDa protein was reported by Fernandez-Pol et al. to be a DNA repair protein (12), containing one zinc finger in its monomeric state, and also involved in carcinogenesis (1).
MPS-1 is being experimentally used as an antiviral target for some agents with low or no toxic effects (1). These novel agents are described in Fernandez-Pol et al., 2004, Patent No.: US 6,803,379. These therapies aim at ejecting the zinc from the zinc finger motif of the MPS-1, thus rendering it inactive or yielding ternary complexes (MPS-1-Zn++–antiviral agent), which result in inactivation of MPS-1 (12).
The observation that highly proliferative and metabolically active cells possess high levels of MPS-1 provided the rationale for targeting MPS-1 to control and/or eliminate cells undergoing uncontrolled proliferation or viral infection (1, 11). Furthermore, extensive work was conducted that demonstrated that agents unrelated to picolinic acid (PA), fusaric acid (FU) or derivatives thereof, that interact with certain metalloproteins and zinc finger peptides (ZFPs) show selective toxicity to virally transformed malignant cells versus untransformed normal cells (1-9; and Fernandez-Pol et al., US patent No. 6,8803,379; 2004).
Materials and Methods
Testing of the cellular and viral actions of compounds to determine various effects on viral-infected cells and control cells, such as cytotoxicity, apoptosis, viral inhibition assays, viral DNA/RNA inhibition assays was done as previously described (1-9, 18, 19). Unless otherwise indicated, standard virus strains and cloned cell lines were used in these studies. Culture conditions for each virus type and cells were carrying out as previously specified (1). Real time-polymerase chain reaction (RT-PCR) was carried out in accordance with protocols and manufacturers specifications. In addition to standard antiviral assays and RT-PCR, antiviral assays and cytotoxicity assays were performed by Advanced Biotechnologies, Columbia, MD, USA.
Evaluation of the assays, validity and test results was performed as previously described (1). DNA repair protocols for eukaryotic systems were carry out following protocols described in (19).
Apoptosis. Apoptosis was evaluated by microscopic examination after the appropriate chemical reactions as previously described (1). The various antiviral compounds were obtained in a highly purified form and further validated in our laboratory by muss spectroscopy (MS). The test compounds were dissolved in various solvents as previously described (1, 18, 19) or in culture media (1).
Virus. Virus strains, cells clones, controls, virus isolation and quantization, viral absorption, detection of virus, viral antigens, and viral nucleic acid were performed following established protocols described elsewhere (18). Viruses and cells were obtained from the ATCC, Bethesda, MD, USA.
Nuclear magnetic resonance (NMR). NMR-based detection of zinc loss from viral and cellular proteins was used. NMR can be used to monitor the loss of zinc from cloned and purified viral and cellular proteins. NMR is a general technique and its many applications are used in this paper to monitor protein–ligand interactions and also the formation of ternary complexes (12). Proteins bound to zinc and those not share a different local environment from proteins which lack zinc. These lead to distinct NMR spectra for both species. For example, by monitoring the proton (1H) spectrum of a sample containing zinc-chelated retroviral Np7 ZFP (4, 5) over time, it is possible to measure if the Np7 retroviral protein loses Zn++ upon exposure to the test antiviral compound that will disrupt the Zn++ binding and eject it from its binding site to the ZFP.
Molecular modeling and docking of experimental antiviral compounds. The aim of the docking protocol is to produce a complex between the antiviral anti-ZFP ligand and the target ZFP which could be a viral or cellular ZFP. This optimizes the environment that resembles physiological conditions, optimizes geometry, and chemical complementarity. The protocols are briefly described in the Results section of this paper. The molecular modeling of ZFP–ligand interactions was performed with a state of the art computer system (Silicon Valley, Laboratory of Molecular Oncology, DVA Medical Center, St. Louis, MO, USA).
Materials and other techniques. Materials and other techniques were previously described in detail by Fernandez-Pol (1-9). Some specific methodological details for certain viruses were as described in Results. Polypeptides were synthesized under contract by the Protein Chemistry Laboratory, Washington University School of Medicine, St. Louis MO, USA.
Antiviral activity and cytotoxicity of PA and FU against herpes simplex virus (HSV)-1 and HSV-2. The efficacy of PA and FU against HSV-1 and HSV-2 were evaluated in cell-based assays. The antiviral activities of PA and FU against HSV-1 and HSV-2 were evaluated using viral plaque reduction assays.
The antiviral effects of PA and FU on HSV-1 and HSV-2 on DNA synthesis were measured by quantitative detection of HSV DNA using RT-PCR assays (Light Cycler™; Roche, USA).
The cytotoxicity activities of PA and FU were evaluated using the fibroblast cell line HEL-299, and red dye uptake assays.
Antiviral activity and toxicity of PA and FU against CMV. The efficacy of PA and FU against CMV were evaluated using AD169 and HEL-299 cells. The antiviral activities of PA and FU against CMV-infected cells were evaluated using viral plaque reduction assays.
The antiviral effects of PA and FU on CMV DNA synthesis were measured by a quantitative detection of CMV DNA using RT-PCR assays (Light Cycler).

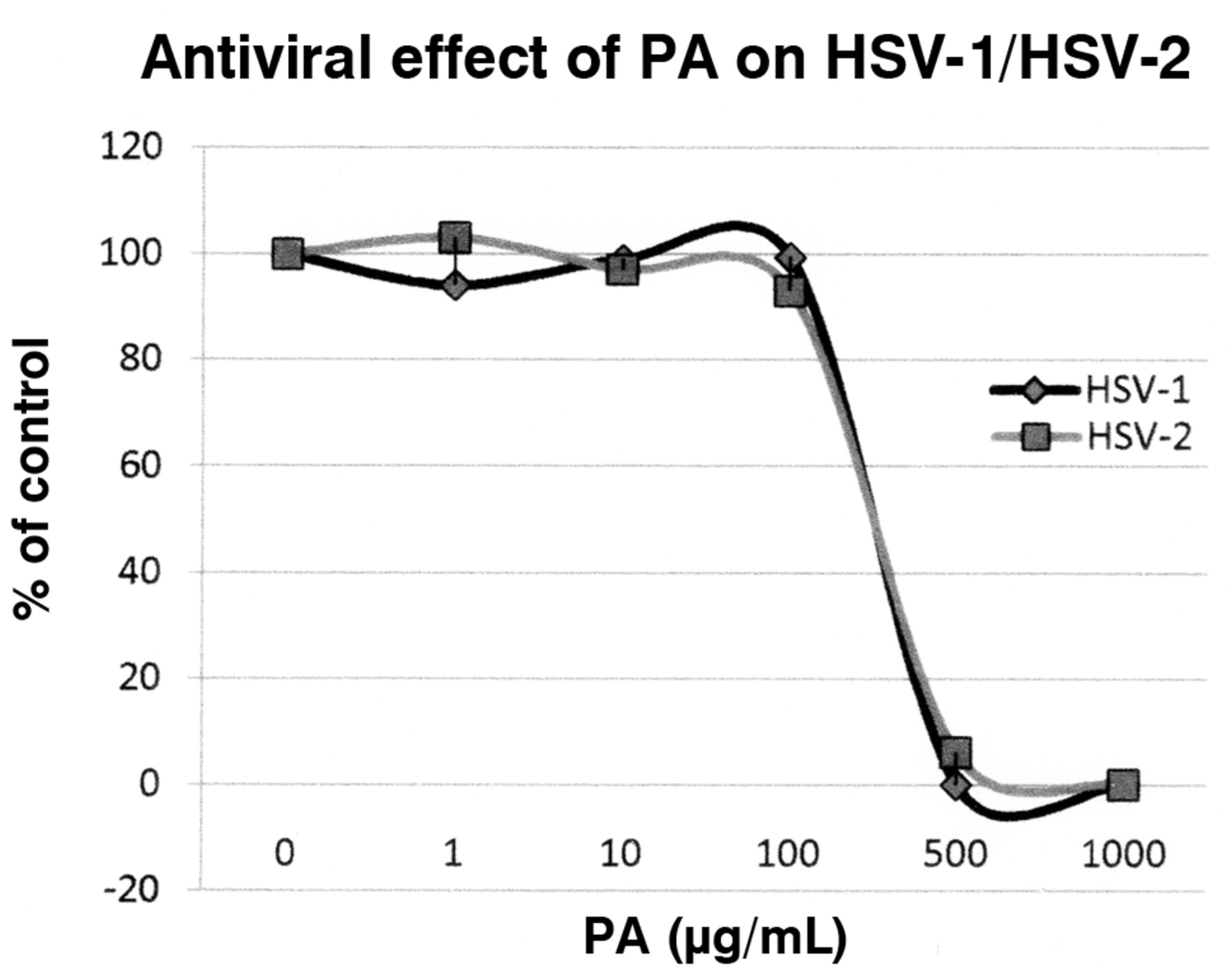
The antiviral effect of picolinic acid (PA) on HSV-1 and HSV-2 viruses was demonstrated by plaque reduction assay. The effect of PA is expressed as percent of control. This percentage was determined by the mean plaque counts in PA-treated cells divided by the mean plaque counts in control cells.
The cytotoxic activities of PA and FU were evaluated using the fibroblast cell line HEL-299, and red dye uptake assays.
Antiviral activity and toxicity of PA and FU against VZV. Confluent HEL-299 cells in 24-well plates were washed and subsequently infected with VZV Ellen strain at an MOI of 0.5 pfu/cell for 2 h at 37°C. The viral inoculums were removed and the infected cells were washed and culture media containing increasing concentrations of PA or FU were added for 72 h. The culture medium was then collected and the released virus DNA was extracted. The culture medium was digested with proteinase K at 37°C for 18 h, and further processed for extraction of extracellular virus DNA.
For purification of the intracellular VZV DNA, the attached cells were washed with PBS and lysed with proteinase K SDS lysis buffer. The released virus DNA was digested with proteinase K at 37°C for 18 h. The VZV DNA was purified by standard procedures.
The detection of VZV DNA was by RT-PCR with the Light Cycler assay. The PCR reaction contained the following: dH2; MgCl2 (25 mM); 10 μM sense primer; 10 μM antisense primer; Light Cycler Probe 1 (fluorescein-labeled, 5 μM); Light Cycler Probe 2 (LC RED 640-labeled, 10 μM); LC Fast Start DNA Master Hyb Probes (2 μL); and uracil DNA glycosidase (1 μL). PCR mix was aliquoted and 5 μL DNA template was then added to the PCR mix (20 μL). VZV DNA standard was diluted (1×107, 1×106,...) and used for quantitation. The cycling conditions were similar to those described by the manufacturer.
Antiviral activity and toxicity of PA and FU against hepatitis B virus (HBV). HepG cells containing the HBV genome were used to assay the antiviral activities of PA and FU against HBV. The test was performed in 96-well trays by standard tissue culture procedures previously described. After 9 days of continuous exposure to the drugs, the culture medium containing released HBV virions was collected, the cells were washed twice with PBS and subsequently lyzed by addition of lysis buffer. All samples were stored at –70°C until assayed.

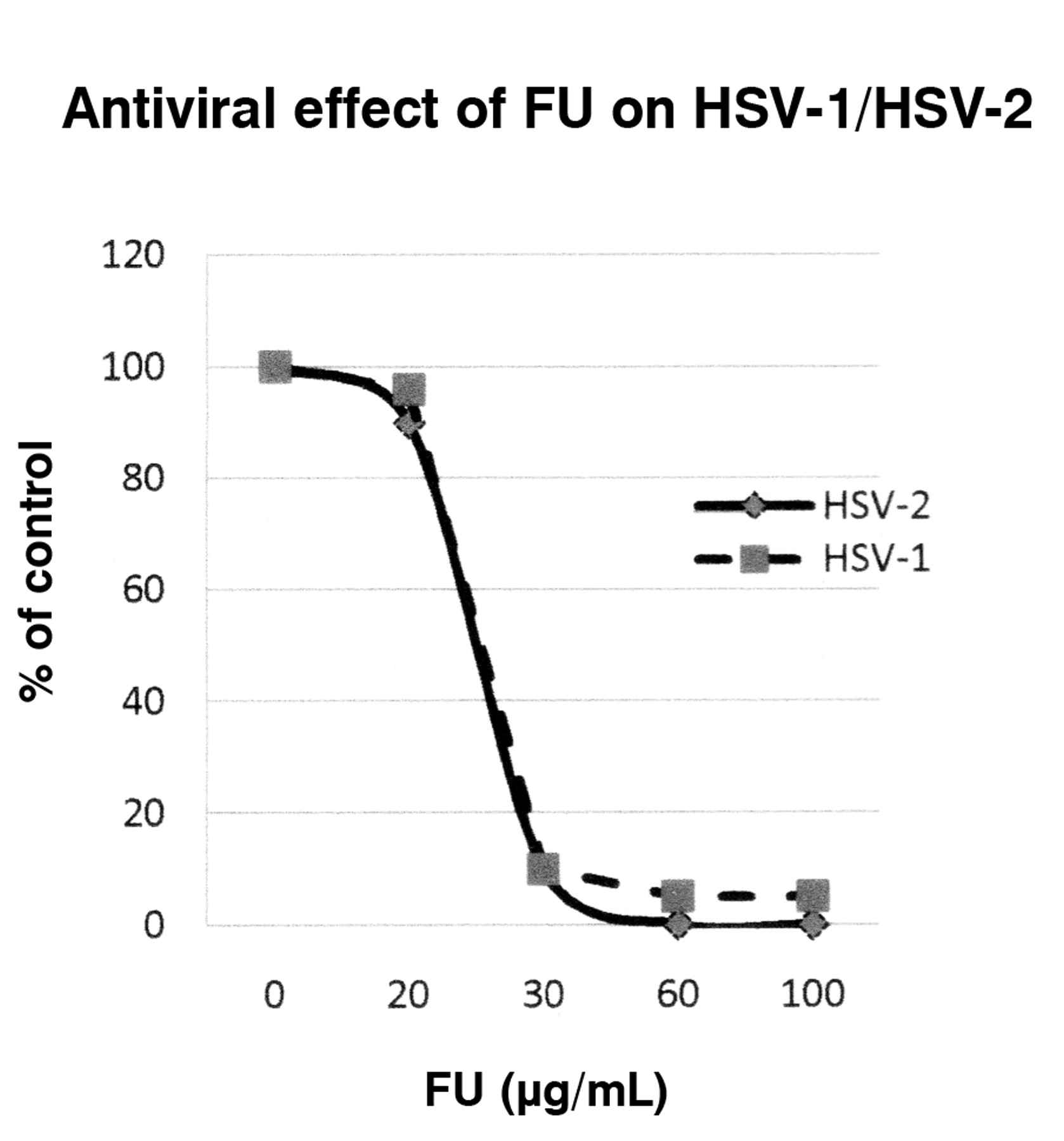
The antiviral effect of fusaric acid (FU) on HSV-1 and HSV-2 viruses was demonstrated by plaque reduction assay. The effect of FU is expressed as percent of control. This percentage was determined by the mean plaque counts in FU-treated cells divided by the mean plaque counts in control cells.
HBV DNA from cell culture media and cell lysates were extracted using the commercially available QIAamp DNA kit, according to the manufacturer's instructions.
The detection of HBV DNA was by RT-PCR with the Light Cycler assay, similarly as described above. The cycling conditions were similar as described by the manufacturer.
Results
Antiviral, cytotoxic and apoptotic activities of PA and FU against HSV-1 and HSV-2. We have previously published the antiviral, cytotoxicity and apoptotic effects of PA against the human HSV-2 infected cells in tissue culture cells (3). The results showed that PA initially causes cytotoxicity, which in turn results in apoptosis of cells infected with HSV-2 and thus reduces the amount of viral particles and arrests viral replication (3).
In this paper the antiviral activities, mechanism of action, and toxicity profiles of PA and FU against HSV-1 and HSV-2 were evaluated using cell-based assays under different conditions than those previously described (3).
The antiviral activities of PA and FU against HSV-1 and HSV-2 (Figures 1 and 2) were evaluated using viral plaque reduction assays. The results indicate that both agents are active against HSV-1 and HSV-2 cellular infections. For PA, the EC50 (defined as the effective concentration giving 50% plaque assay reduction) was approximately 236 μg/mL for HSV-1 and 248 μg/mL for HSV-2. The EC90 (effective concentration reducing 90% plaque formation) was about 500 μg/mL for both HSV-1 and HSV-2. For FU, the EC50 was approximately 19 μg/mL for HSV-1 and 18 μg/mL for HSV-2. The EC90 was approximately 30 μg/mL for both HSV-1 and HSV-2. The cytotoxicity profiles of both agents were determined on Vero cells with CC50 (cytotoxic concentration causing 50% cell death) of approximately 800 μg/mL for PA and 100 μg/mL for FA, respectively (Figure 3).
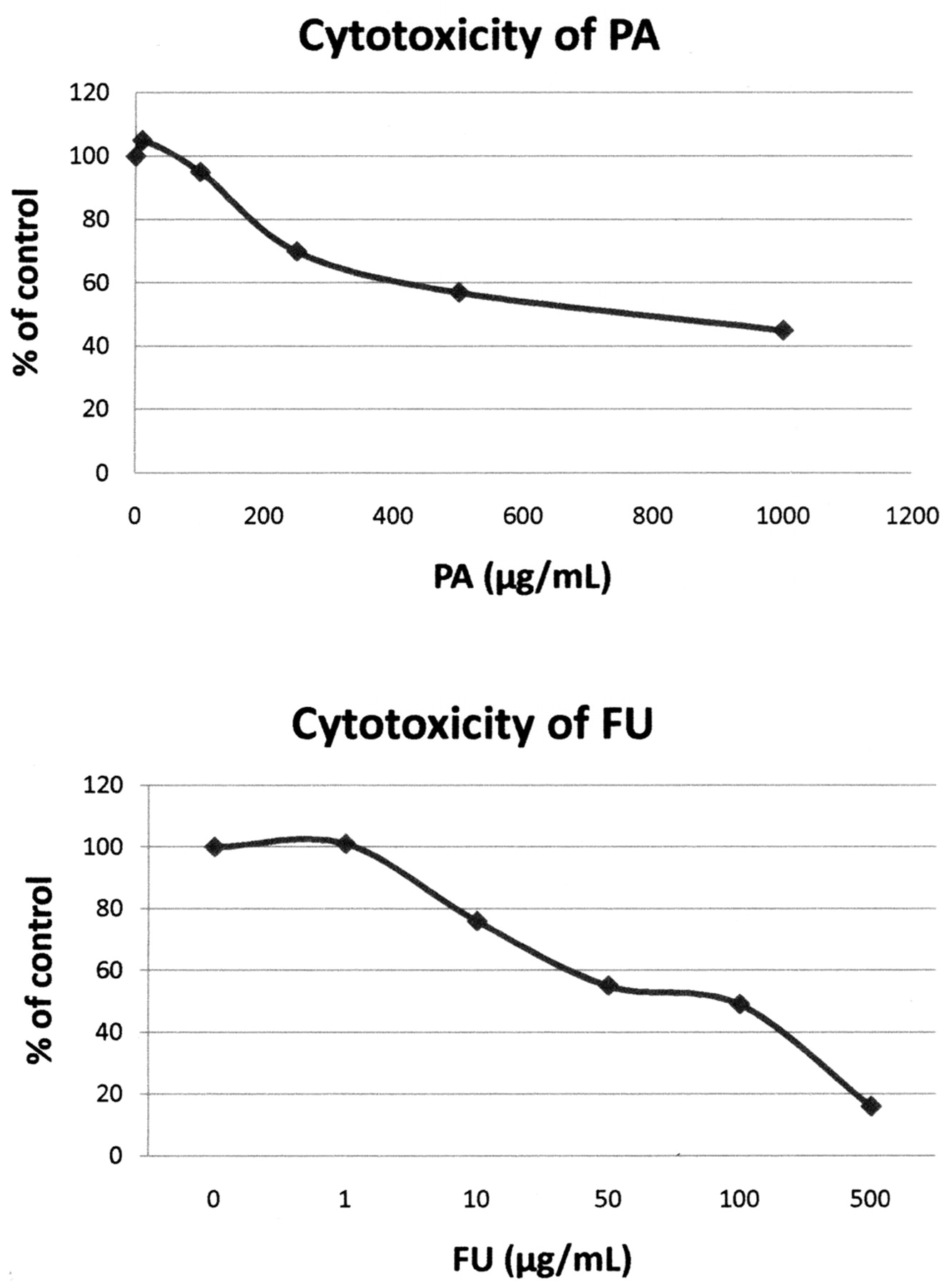
The dose-dependent cytotoxicity of picolinic acid (PA) and fusaric acid (FU) towards Vero cells were determined by vital dye uptake assay. The cytotoxicity of both agents at increasing concentrations is expressed as percent of control. This percentage was obtained by determining the mean O.D. at 550 nm (neutral read) in PA- and FU-treated cells and dividing by the mean O.D. at 550 nm in control cells.
When FU was added at 50 μg/mL to HSV-1-infected cells from 0 to 8 h post infection (p.i.), a period of the viral cycle in which viral DNA is replicating, inhibitory effects were about 99% effective. When FU was added at 20 h p.i., a stage at which the virions are in the process of being released, the inhibitory effects of FU were reduced by 50% (Figure 4).

Time-dependent antiviral effect of fusaric acid (FU) on HSV-1. The cells were treated with FU at the following times: T-2, 2 hours before infection; T0, simultaneous addition of HSV-1 virus and FU at the initiation of the experiment (time=0); 2, 4, 8, 17, and 20 hours after the initiation of the experiment (T=0). At each time point, the culture medium was aspirated and replaced with methylcellulose containing increasing concentrations of FU to perform the plaque assay. The antiviral action of FU is shown as a percentage of the control.
The antiviral effects of PA and FU on HSV-1 DNA synthesis were measured by quantitative detection of HSV DNA using RT PCR assays (Light Cycler). The results showed that PA at concentrations higher than 100 μg/mL and FU at concentrations higher than 10 μg/mL reduced HSV-1 DNA synthesis in drug-treated HSV-1-infected cells (Figure 5).
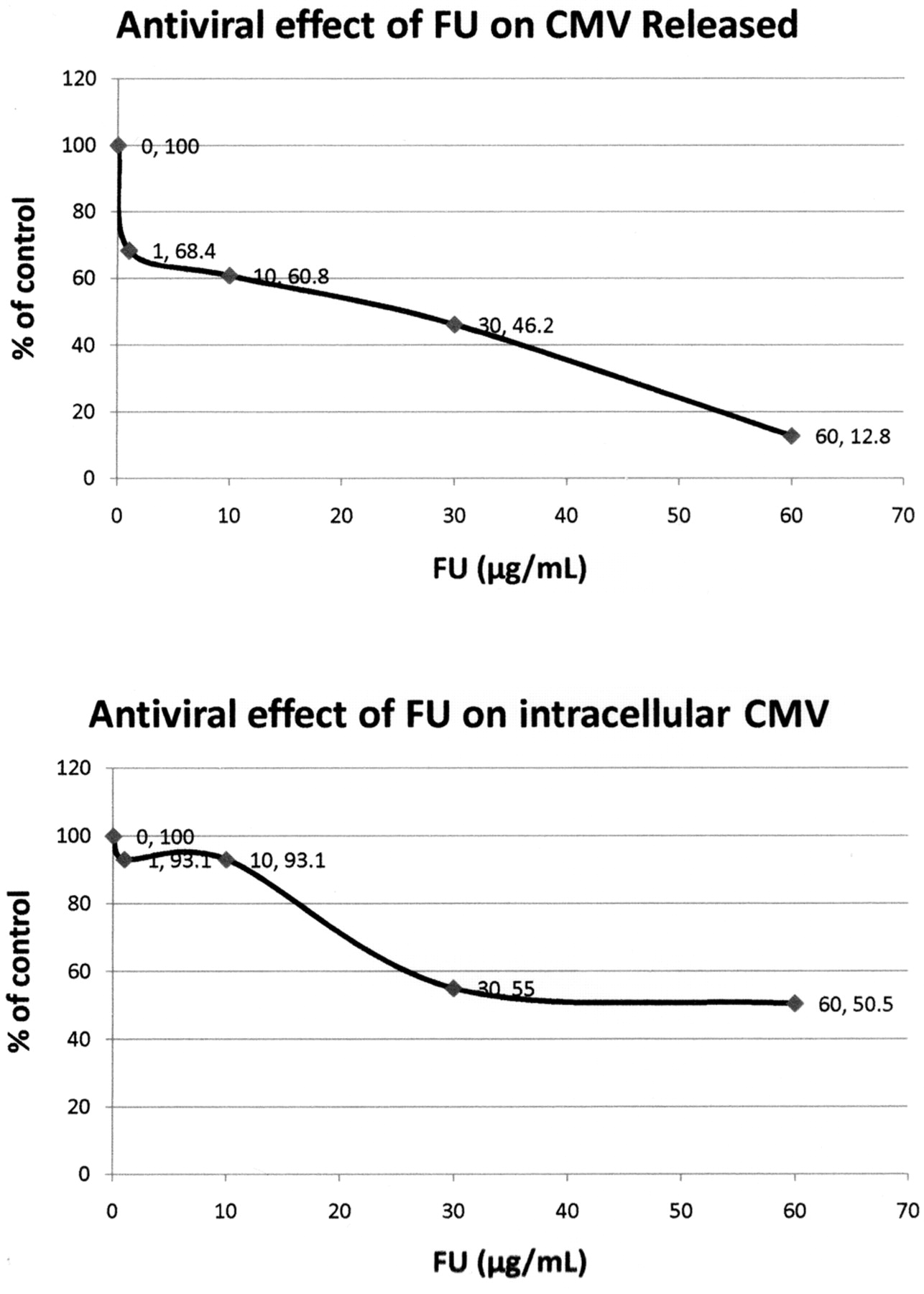
Antiviral, cytotoxicity and apoptotic activities of PA and FA. The antiviral effects of PA and FU against CMV–infected cells were evaluated using the reference strain AD169 and HEL-299 cells. The CMV-infected cells were treated with increasing concentrations of PA and FA for 72 h. Both the culture media and the cells were collected for extra- and intracellular virus. The results show that both PA and FU are active against CMV replication with EC50 of about 230 μg/mL for PA and approximately 69 μg/mL for FU (Figures 6 and 7).
Cytotoxicity was observed at approximately 1000 μg/mL PA and 100 μg /mL FA. Antiviral, cytotoxicity and apoptotic activities of PA and FA on varicella-zoster virus (VZV).
The antiviral effects of PA and FU on VZV were evaluated using VZV reference strain Ellen and HEL-299 cells. Virus yield reduction assay was performed by quantitative detection of total VZV DNA copies as an end-point for anti-VZV susceptibility test for 72 h. The culture medium was collected and the cells were lysed for both detection of released virion DNA and for detection of intracellular virus DNA. Quantitative detection of VZV DNA copy number was determined by Light Cycler assay.
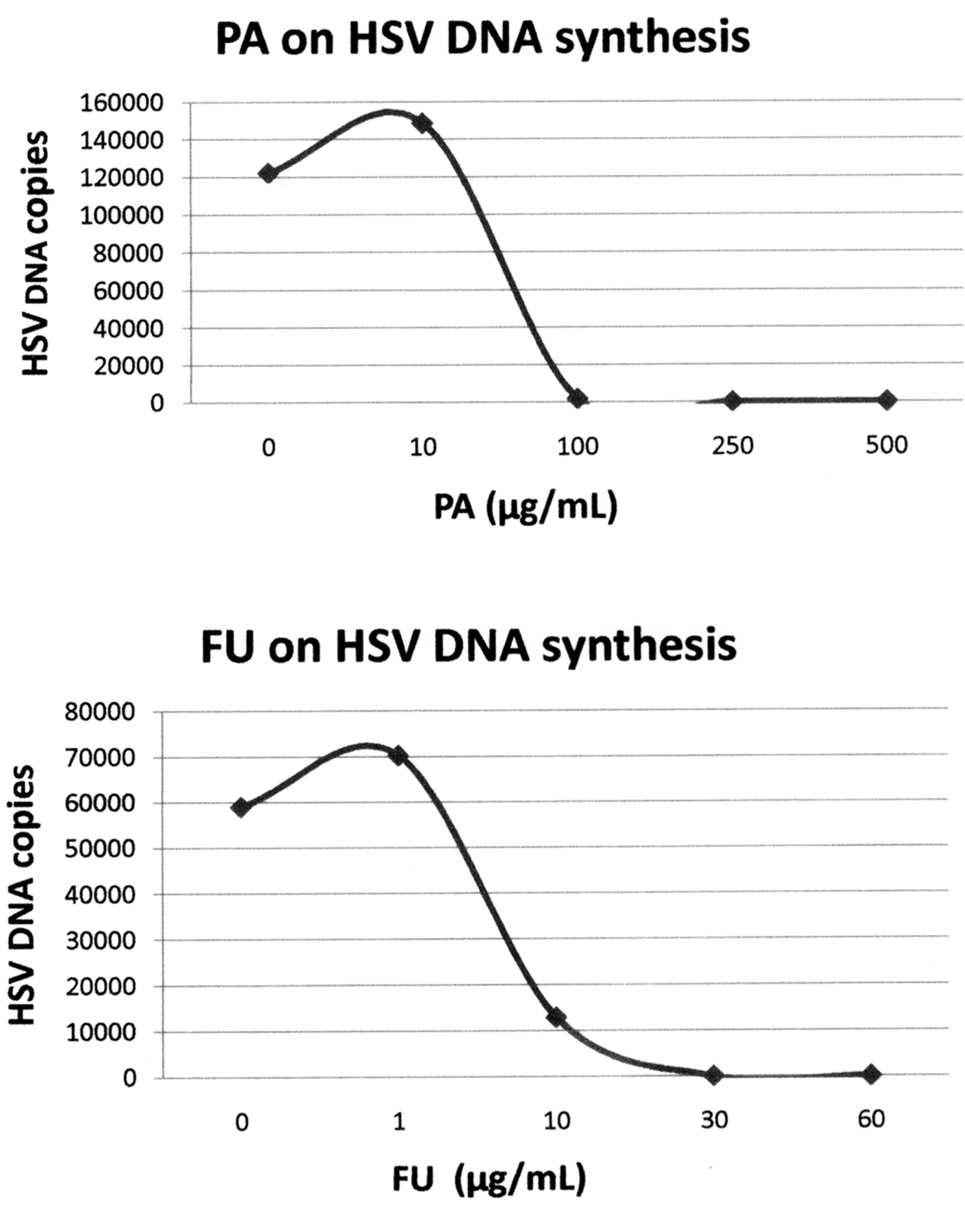
Effects of picolinic acid (PA) and fusaric acid (FU) on HSV-1 DNA synthesis. Vero cells were infected with HSV-1 as described in the text. After infection, they were exposed to increasing concentrations of PA or FU. After 18 hours post-infection, total DNA (cellular and viral) was extracted from the cells. HSV-1 DNA copies were quantitated by the Light Cycler assay.
The results show that both PA and FU are strongly active against VZV (Figures 8 and 9), with EC50 of approximately 89 μg/mL and 14 μg/mL, respectively. PA was active against VZV replication, with EC50 of approximately 89 μg/mL; the EC90 for PA was 500 μg/mL. FU also showed anti-VZV activity, with EC50 of approximately 14 μg/mL and EC90 of approximately 41 μg/mL (Figures 8 and 9). No significant cytotoxicity was observed on HEL cells treated with PA (500 μg/mL) and FU (60 μg/mL).
Antiviral, cytotoxicity and apoptotic activities of PA and FA HBV.
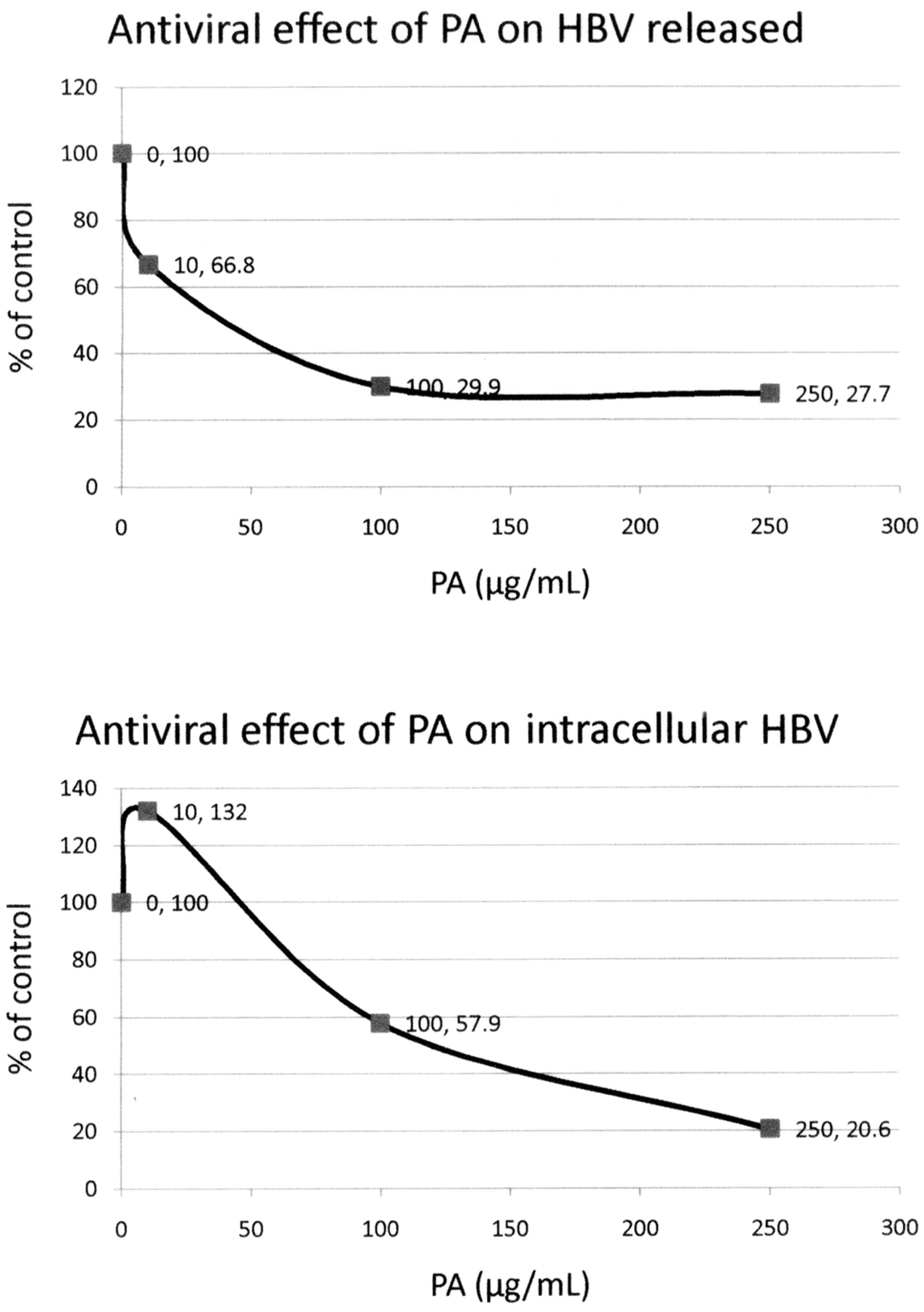
The antiviral effects of PA (Figure 10) and FU (Figure 11) on HBV were examined using HepG2.2.15 cells, treated for 9 days with media changed every 24 h. After 9 days, the medium was collected and the extracellular virus DNA was extracted. The cells were lyzed with 0.5% SDS and the intracellular virus HVB DNA was extracted using a Qiagen column and then quantitatively detected by RT-PCR assay.

Antiviral effect of picolinic acid (PA) against cytomegalovirus (CMV)-infected cells. CMV-infected cells were treated with increasing concentrations of PA for 72 hours. The culture medium was collected for measurement of released CMV and the cells were collected for determination of intracellular virus. The results show that PA inhibits CMV replication with EC50 of approximately 200 μg/mL (released CMV) to 230 μg/mL (intracellular CMV). The antiviral effect of PA on CMV was determined by quantitation of the mean released or intracellular CMV DNA copy number using real-time PCR. The results are shown as a percentage of the control.
The results showed that PA at 250 μg/mL inhibited the released HBV DNA by 72.3% and the intracellular HBV DNA by 79.4%, with overall EC 50 of approximately 139 μg/mL (Figure 10). FU at a concentration of 60 μg/mL inhibited released HBV DNA by 76.5% and intracellular HBV DNA by 28%, with overall EC50 greater than 60 μg/mL (Figure 11).
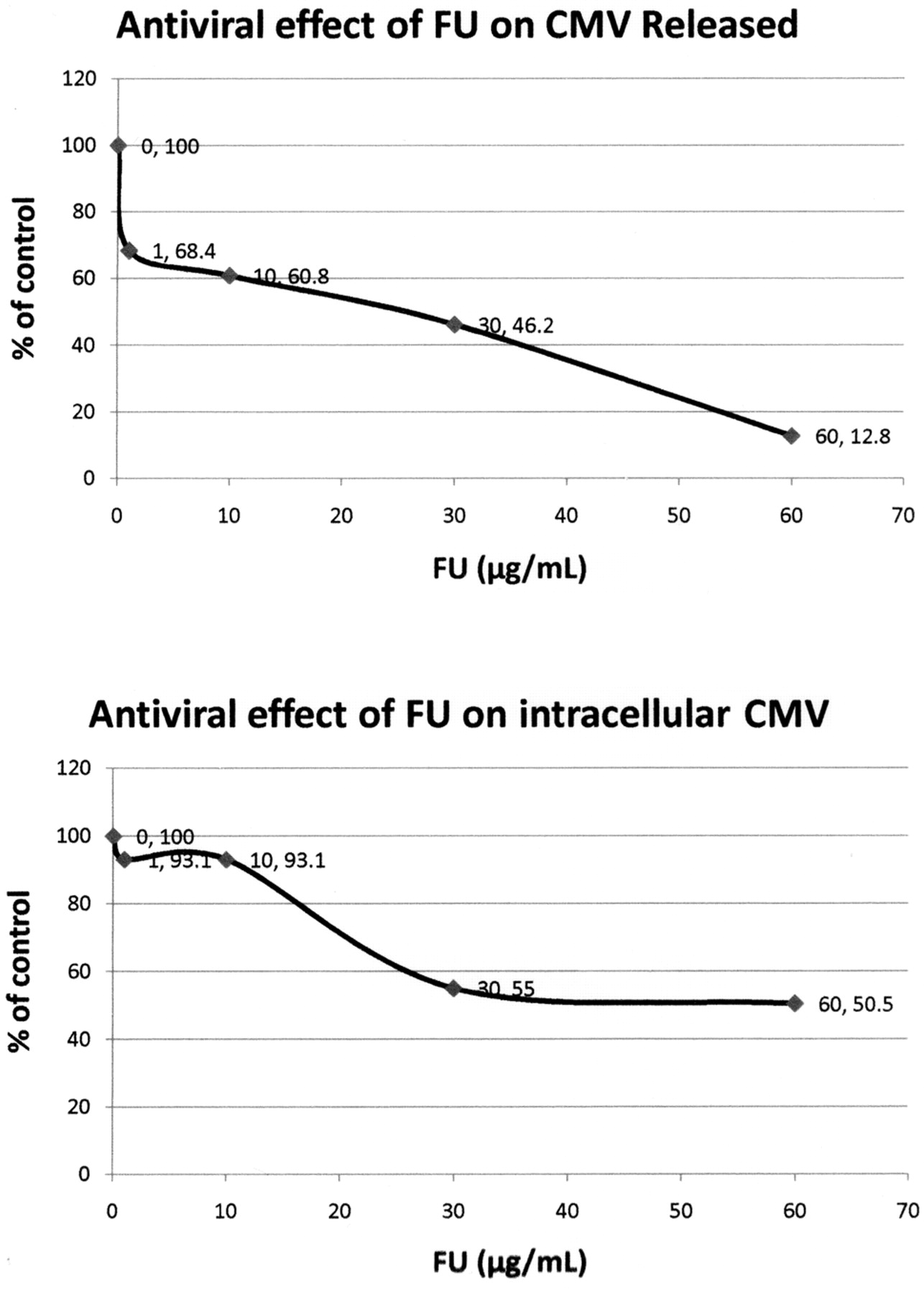
Antiviral effect of fusaric acid (FU) against cytomegalovirus (CMV)-infected cells. CMV-infected cells were treated with increasing concentrations of PA for 72 hours. The culture media was collected for measurement of released CMV and the cells were collected for determination of intracellular virus. The results show that PA inhibits CMV replication with EC50 of approximately 200 μg/mL (released CMV) to 230 μg/mL (intracellular CMV). The antiviral effect of PA on CMV was determined by quantitation of the mean released or intracellular CMV DNA copy number using real-time-PCR. The results are shown as a percentage of the control.
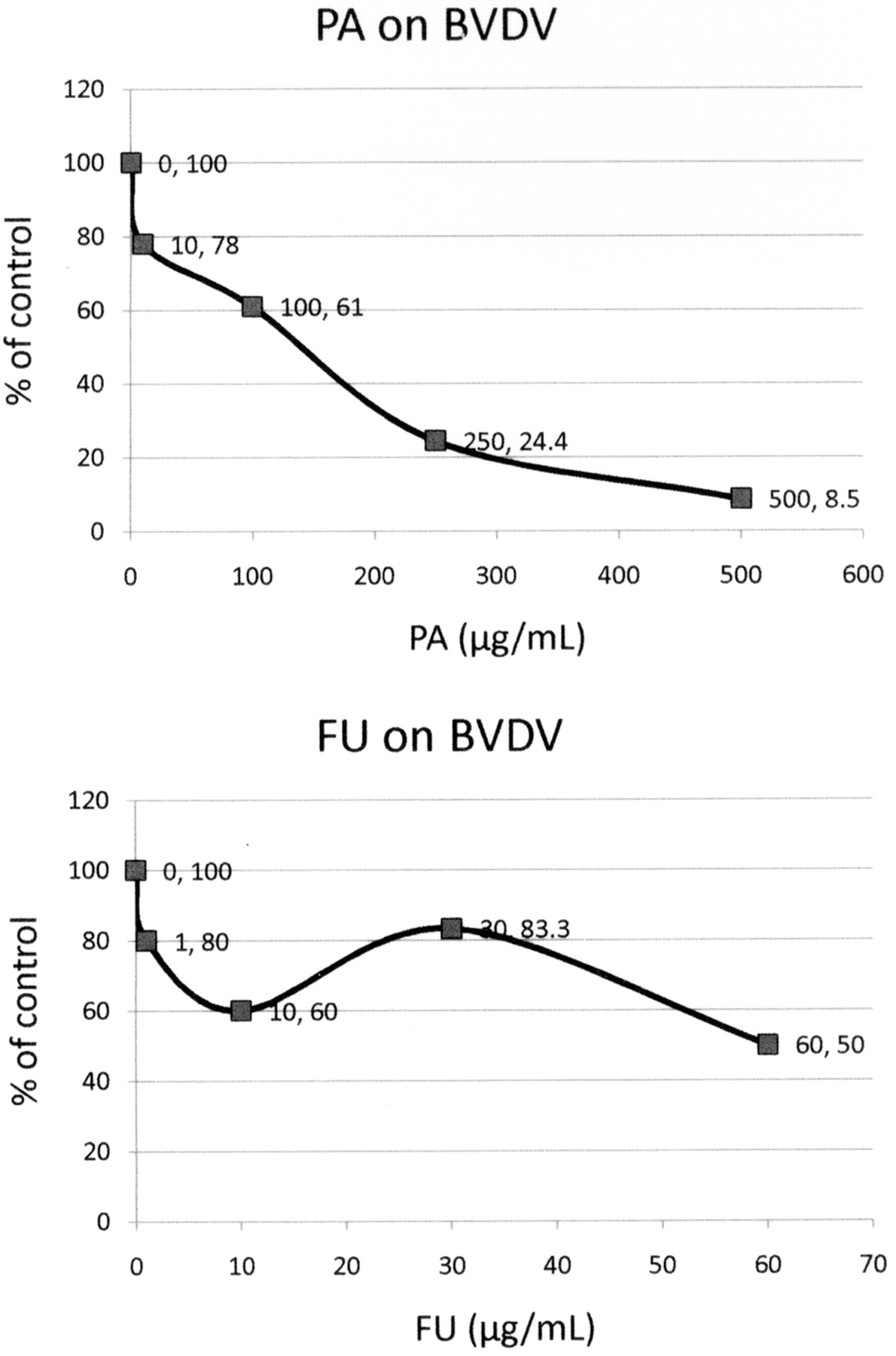
Antiviral, cytotoxic and apoptotic activities of PA and FA on bovine viral diarrhea virus (BVDV). The antiviral effects of PA and FA against BVDV were evaluated using BVDV Trangie strain and MDBK cells (Figure 12). BVDV yields were determined by immunoperoxidase staining with monoclonal antibodies against NS3. The results showed that after treatment of infected cells for 72 h with PA at 250 μg/mL reduced BVDV virus yield by 76% (EC50=130 μg/mL). FU at 60 μg/mL inhibited BVDV virus yield by 50% (EC50=60 μg/mL). Thus, PA and FU are active against BVDV. PA at 1000 μg/mL and FU at 100 μg/mL were both cytotoxic.

Antiviral effect of picolinic acid (PA) on Varicella-Zoster virus (VZV). The antiviral effect of PA on VZV was determined by quantization of the Mean Total VZV DNA copy numbers using real-time PCR. The results are shown as percent of control. EC50=90.0 μg/mL; EC90=500 μg/mL.

Antiviral effect of fusaric acid (FU) on VZV. The antiviral effect of FU on VZV was determined by quantitation of the mean total VZV DNA copy numbers using real-time PCR. The results are shown as a percentage of the control. EC50=14 μg/mL; EC90=30 μg/mL.
Antiviral activities of PA and FA on HCV and Sindbis replicons. In pilot experiments, we studied the effects of PA and FU on HCV replicons and Sindbis replicons and by qRTPCR/icycler.
The cells were grown in the presence of PA or FU in 24 well culture plates, for 24 hours. Sindbis-Huh7b cells and HCV replicon-Ava.5 cell lines were tested. Normalization was by standard OD and serial dilutions using the untreated cell lines. RNA was extracted as described elsewhere.
The results presented in Figure 13 with HCV replicon shows that alpha-IFN inhibits by 20% to 10% the HCV replicon at 10U and 100U of alpha-INF, respectively. PA at 100, 200, 300 and 400 μg/mL inhibited HCV replicons to about 50, 51, 41, 36 and 30% of control, respectively. FU at 5, 25, 50 and 100 μg/mL inhibited HCV to 56, 41, 14, and 10% of control, respectively. No detectable cytotoxicity by OD was observed in any of the conditions tested. The results show that the HCV replicon system appears to show a dose–response relationship for both PA and FU, FU being significantly more active than PA (about 4-fold more potent FU than PA).
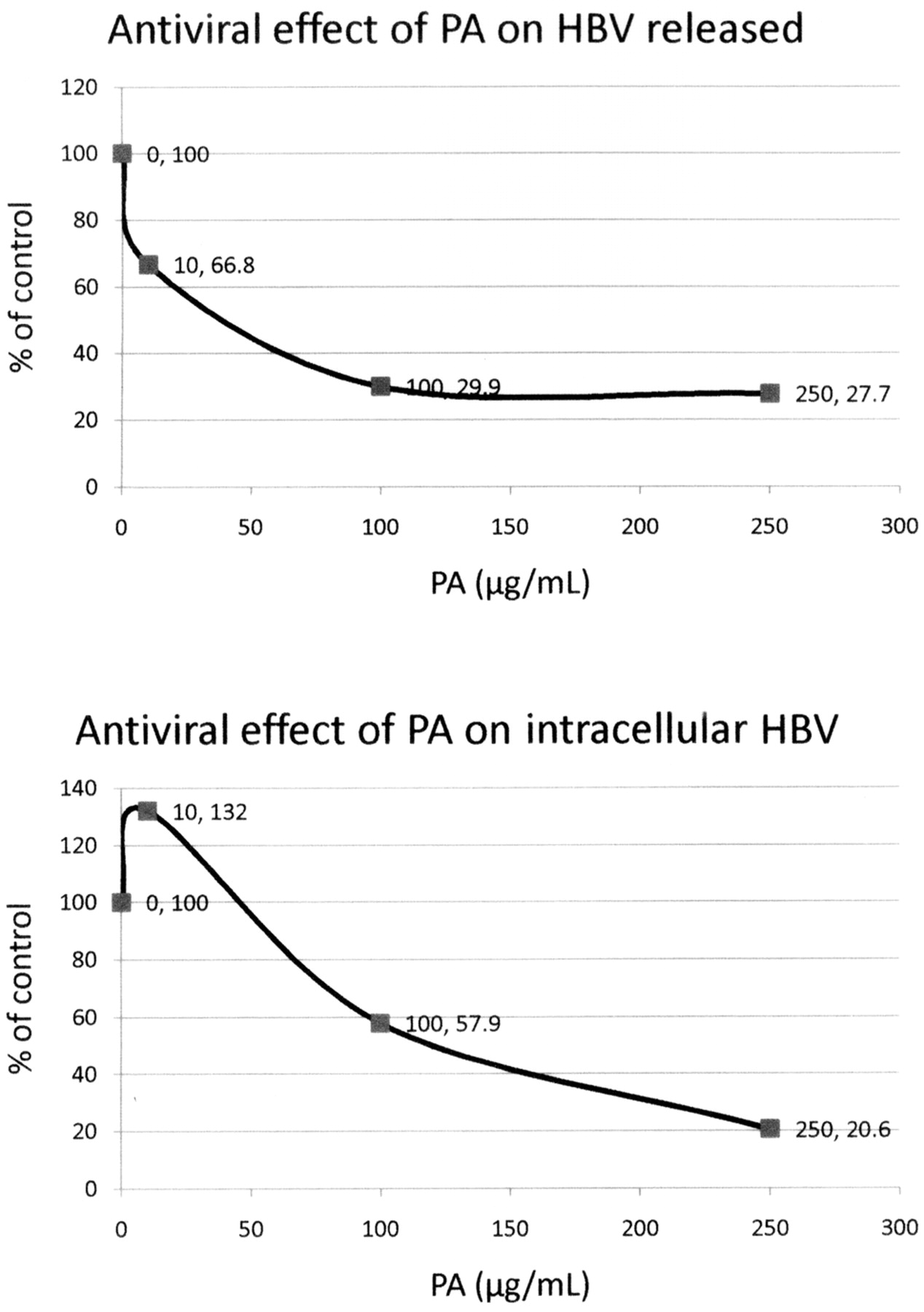
Antiviral effect of picolinic acid (PA) on HBV. HepG2 cells infected with the HBV virus were treated with increasing concentrations of PA for 9 days. Total released HBV DNA and total intracellular HBV DNA were determined by quantitation of DNA copy numbers using real-time PCR. The results are shown as a percentage of the control. Released: EC50=40 μg/mL.
The results presented in Figure 14 with Sindbis replicons, which include alpha-interferon (10 and 100 Us), PA (100 to 400 μg/mL) and FU (5 to 100 μg/mL), had greater than 85% inhibition of replicons at all concentrations of both agents tested. No detectable cytotoxicity by OD was observed in any of the conditions tested. The results show that the Sindbis replicon system is quite sensitive to PA and FU and perhaps a dose-response relationship could be obtained at lower concentrations of these agents.

. Antiviral effect of fusaric acid (FU) on HBV. HepG2 cells infected with the HBV virus were treated with increasing concentrations of FU for 9 days. Total released HBV DNA and total intracellular HBV DNA were determined by quantization of DNA copy numbers using real-time PCR. The results are shown as a percentage of the control. Released: EC50 approximately 45.8 μg/mL.
Potential mechanism of inhibition of FA and PA on HIV-1 RNA virus replication as demonstrated by NMR.
NMR studies of ligand binding by a ZFP from HIV-1: Inhibition of HIV-1 infectivity by zinc-ejecting 2-pyridine carboxylic acid compounds. Retroviral ZFPs with the sequence (CCHC) bind zinc stoichiometrically and with high affinity of the order of 10–12 M. Under physiological conditions of ionic strength and pH of 7.4, a 10-fold excess of EDTA removes less than 50% of the zinc from the N-terminal CCHC zinc finger domain of the HIV-1 nucleocapsid protein Np7. An illustrative diagram of the potential interaction of PA with the Np7 HIV nucleoprotein is shown in Figure 7.
To determine whether PA or FU (Figure 15; Table I) were capable of ejecting zinc from segments of ZFPs corresponding to retroviral-type ZFPs, we performed NMR studies with the following components in the reaction mixture: (i) an 18 amino acid peptide with a sequence corresponding to the N-terminal ZFP-binding domain of the HIV-1 nucleocapsid protein, denoted ZF-peptide 18 [VKCFNCGKEGHIARNCRA]; (ii) Zn2+ which binds zinc etoichiometrically and with a Kd of 10–12; and various PA and FU analogues. Structural studies have shown that the peptide adopts a structure that is essentially identical to that of the corresponding domain in the intact nucleocapside protein under physiological conditions; and (iii) a synthetic oligonucleotide with a sequence corresponding to a region of the HIV-1 psi-packaging signal, d(ACGCC) (17, 20).
NMR of Zn (HIV-1-F1) and pyridine carboxylic acids interactions. We studied the interaction of FU and PA with the ZFP Zn (HUV1-F1) by NMR spectroscopy. The results of these studies indicate that both molecules bind Zn (HIV-1-F1) at the hydrophobic crevice (a hollow between two parts) where the peptide binds an unpaired guanosine base, its natural substrate which was described by South and Summers (1993). Dose-dependent experiments with each of the two molecules caused shifts in the 1H resonances of Zn (HIV-1-F1) that are similar to those observed when the peptide binds analogs of its natural substrate. Two-dimensional NOE correlation spectroscopy (NOESY) of the solution containing Zn (HIV-1-F1) and FU produced cross peaks at frequencies that are consistent with contacts between FU and hydrogen nuclei in the hydrophobic cleft of the peptide.
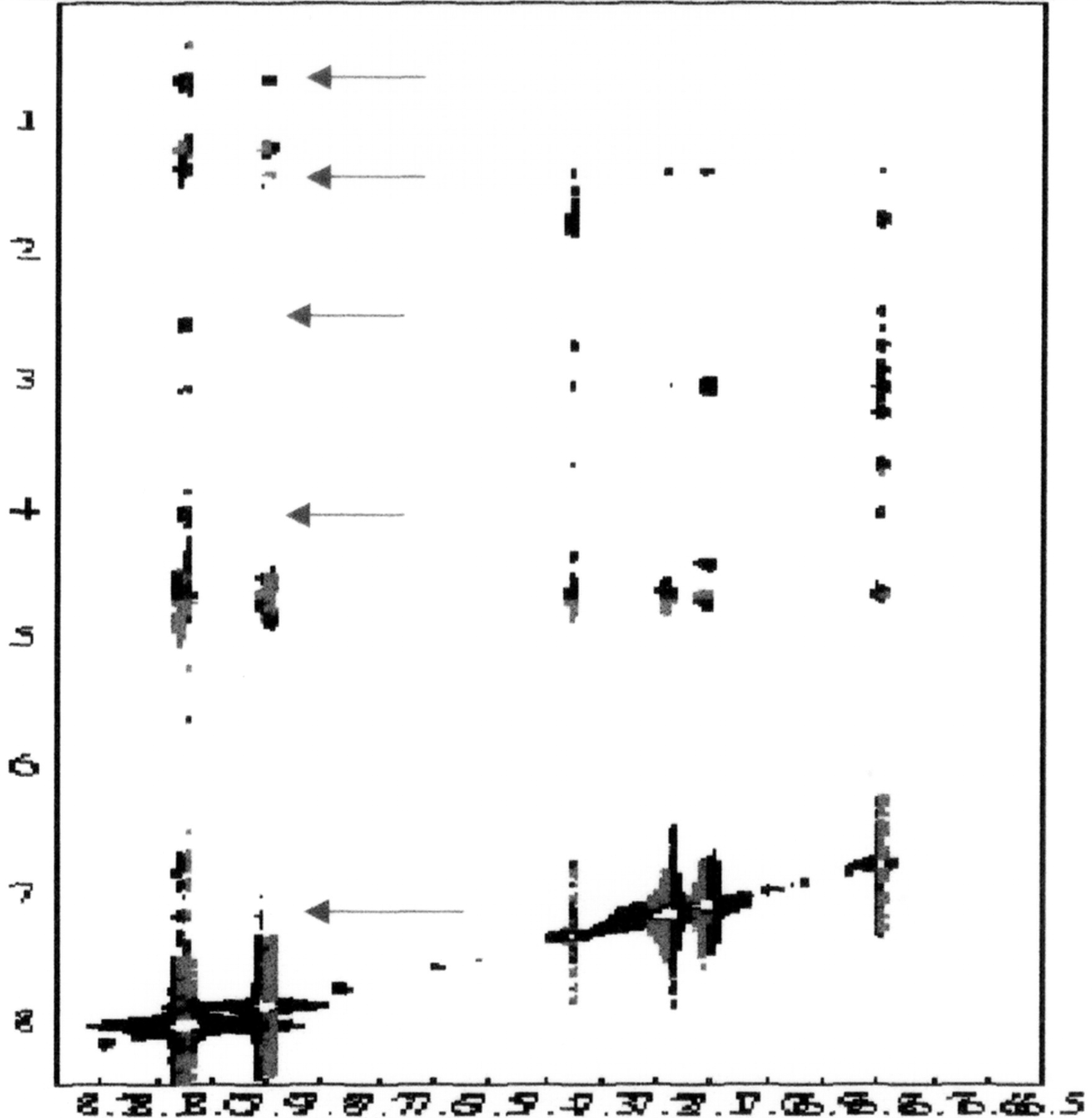
Titration of the peptide Zn (HIV-1-F1) with FU (Figure 16). A sample containing peptide (1.63 mM) and Zn2+ (2.0 mM) was titrated with a solution of FU. The interactions were followed by one-dimensional 1H NMR spectroscopy.
The free zinc in the buffer was titrated with FU. Addition of FU yielded spectral shifts in the spectrum of the peptide. These results indicate that FU competitively binds both Zn and the Zn-peptide complex. These results led us to run a second 2D NOESY experiment, this time at higher concentrations of FU.
2-Dimensional NOESY spectrum of the peptide–FU complex (Figure 17). The NOESY spectrum at high FU concentration was considerably different than that measured at low FU concentration. At high FU concentration, the sign of the NOE contacts involving either the drug or the peptide were positive, indicating that NOEs involving the ligand arose from the ligand–peptide complex. The NOESY spectrum shows cross peaks between an aromatic FU resonance at ∼8 ppm and resonances at 4.06 (petide C-alpha-H), 2.60, 1.41, and 070 ppm. The presence of cross peaks and tentative assignments for these peaks.
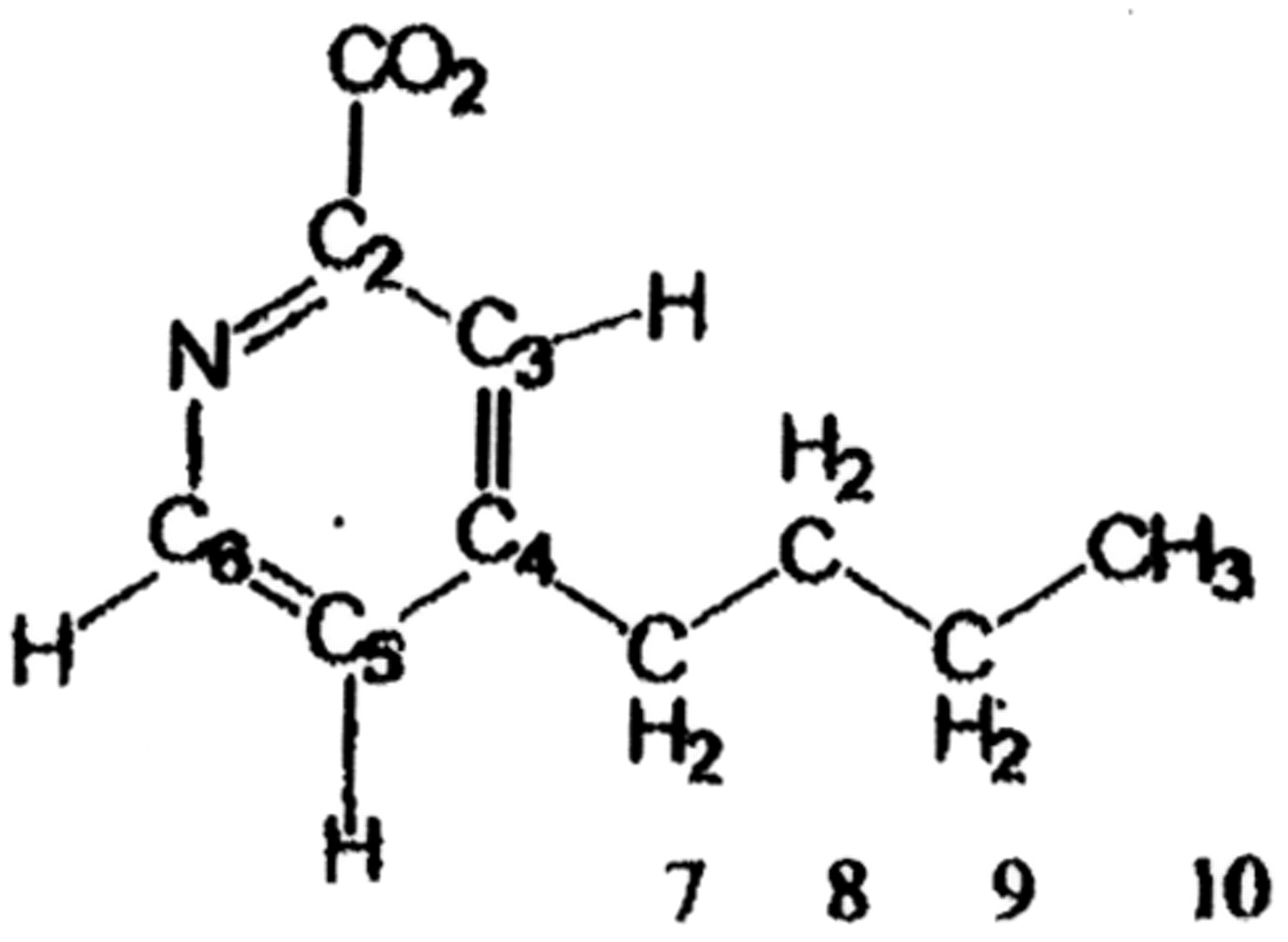
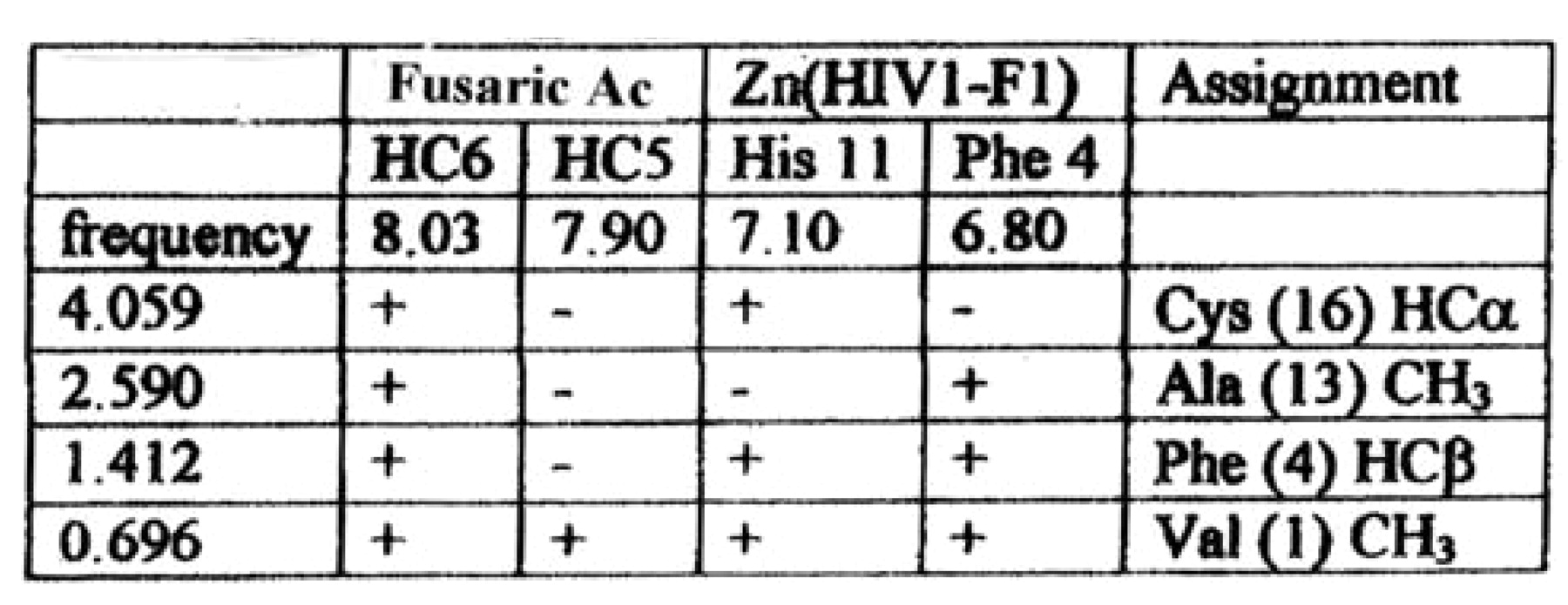
The shapes of the cross peaks involving the FU resonance at 8.03 ppm indicate that these interactions involve a doublet resonance (either HC5 or HC6) and not with the singlet (HC3). Literature reference values indicate that pyridine hydrogen's in the 1 and 6 positions have higher resonant frequencies than those in the 2 and 5 positions. The 8 ppm resonance was therefore assigned to be that of the HC6 hydrogen. The assignment was supported by considering the first NOESY spectrum, where conditions did not favor Zn (HIV1-F1) binding. Under this condition, cross peaks (with negative sign) are observed between the HC3 singlet and the resonance of the HC7 alkyl group (at 2.86 ppm), and between the doublet at 7.90 ppm and the HC7. This confirms that the 7.9 ppm resonance is attributable to HC5 and the 8.05 ppm resonance contains the contribution from HC6.
While the resonance frequencies of the alkyl (H2C7, H2C8, and H3C10) resonances correspond closely to the 2.6, 1.4, and 0.7 ppm cross peaks, neither the HC3 nor HC5 shows a cross peaks at these frequencies. Since both HC3 and HC5 are closer to the alkyl group than HC6, the cross peaks most likely arise from contact between the HC6 of FU and hydrogen nuclei of the peptide, and not those of the FU alkyl chain.
The peak at 1.4 corresponds to the second FU alkyl chain hydrogen frequency, but also covers the Ala 13 methyl resonance. It should be noted that this is one of the resonances that underwent a substantial shift in the 1D titration experiments.
A number of the peptide beta-hydrogens have chemical shifts in the region of the 2.6 ppm cross peak (C3; 2.79, F4; 2.61, N5; 2.80, C6; 2.53, N15; 2.56). The Phe 4 hydrogen assignment is favored because of its spatial proximity to other hydrogens whose resonant frequencies are affected by FU (this residue is in the hydrophobic cleft). We note that the 3.1, 2.6, 1.4, and 0.7 ppm resonances also shows cross peaks to the Phe 4 aromatic resonances, indicating that FU binds in close proximity to this residue.
The data do not support the idea that the FU methyl group contacts the FU aromatic hydrogens. The problem is that the resonance of the FU methyl overlaps those of the peptide methyl group from Val 1 and Ile 12, both of which are in the hydrophobic cleft, and which shift in the 1-D titration experiments. On the basis of other NOE contacts to the methyl group in question, it may be inferred that the contact is probably with Val 1.
It is unlikely that the ligand bind at two separate sites on FU. While the cross peaks that connect the aromatic resonances of FU to the aromatic resonances of both His 4 and Phe 4 peaks give the appearance that this ligand binds to two separate positions on the peptide, we believe that the cross peaks to the His resonances are not due to NOE transfer but due to spin diffusion through the Cys HC-alpha which is in contact with the ligand. The fact that the His aromatic resonances do not shift during the titration experiment support this notion.
Titration of the ZFP with PA. We have observed that under conditions in which the buffer becomes too acidic that zinc is lost from the peptide and is then complexed with PA. Under conditions in which the PA stock solution was neutralized with Tris base, binding could be measured. In this experiment, the effect of PA on the spectrum of the peptide was similar to that observed with FU. The same protein resonances observed to shift with addition of FU shifted in the same direction (although to a somewhat lesser extent) with PA.
One-dimensional NOE experiments failed to establish contacts between the peptide and PA. Since spectral overlap with FU leads to ambiguity in assigning NOEs, we ran a series of one-dimensional NOE difference experiments. One-dimensional NOE difference spectroscopy failed to detect NOEs between Zn (HIV-1-F1) and PA. We proposed that the method did not have sufficient sensitivity to measure these weaker contacts.
One-dimensional NOE difference experiments demonstrated NOE contacts between the aromatic hydrogens of PA and His 11 and between PA and Phe 4 but these were considerable weaker than those measured for FU.
Space models representing the structure of the peptide were generated from protein database file for the peptide complexed with dACGCC (17, 20). Figure 23 show a model of binding contacts for FU, relative to the guanosine base in the binding grove of the peptide (Figures 19, 20, 21, 22, 23).

Antiviral effects of picolinic acid (PA) and fusaric acid (FU) on BVDV. The antiviral activity of PA and FU were determined by using immunoperoxidase staining with monoclonal antibodies directed against the viral nucleoprotein NS3. Increasing concentrations of both PA and FU were used. The results are shown as a percentage of the control. For PA the EC50=110 μg/mL and the EC90=500 μg/mL. For FU, the EC50=60.5 μg/mL and the EC90 could not be determined.
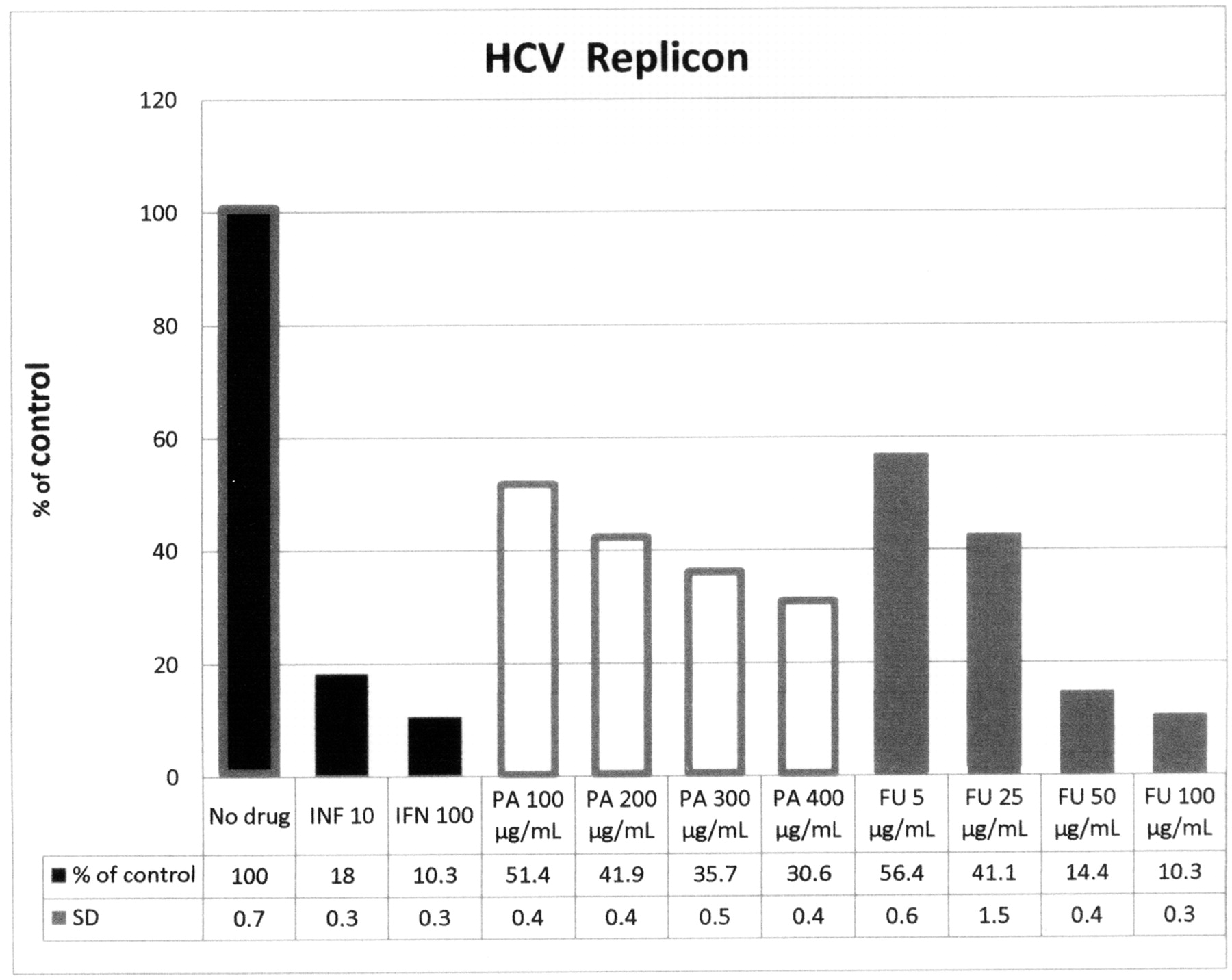
Antiviral activity of picolinic acid (PA) and fusaric acid (FU) on HCV replicons. The HCV replicons were contained in Ava.5 cell line. The cell lines were normalized by OD and serial dilution. The Ava.5-HCV replicons were exposed to the compounds for 24 hours. Interferon-alpha (IFN) was used as a positive control. After RNA extraction, the HCV replicon copy numbers were quantified using real-time PCR. The results are shown as a percentage of the control, no drug additions. The data indicates that for PA the EC50=100 μg/mL. For FU, the EC50 is approximately 5 μg/mL. The inhibitory effects of both PA and FU were dose-dependent. O.D. showed no overt cytotoxicity. SD did not exceed 0.7%.
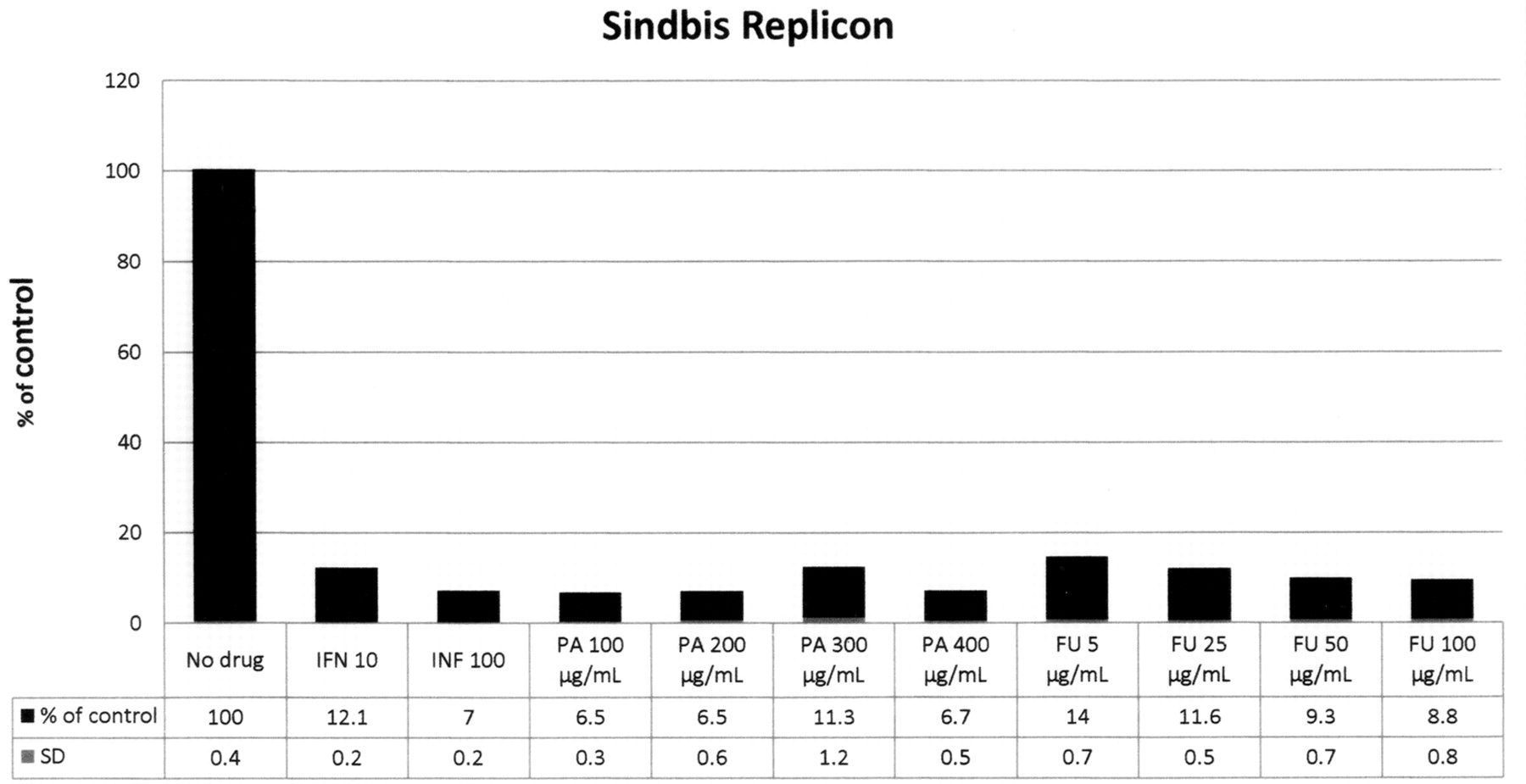
Antiviral activity of picolinic acid (PA) and fusaric acid (FU) on Sindbis replicons. The Sindbis replicons were contained in Huh 7b cell line. The cell lines were normalized by OD and serial dilution. The Huh 7b-Sindbis replicons were exposed to the compounds for 24 hours. After RNA extraction, the Sindbis replicon copy numbers were quantified using real-time PCR. The results are shown as a percentage of the control, no drug additions. The data show greater than 90% inhibition of Sindbis replicon at all concentrations of PA tested. FU show >85% inhibition of Sindbis replicon at all concentrations tested. No overt cytotoxicity was observed by OD. SD did not exceed 0.8%.


and Table I. Structural formula for fusaric acid (FU) shown on the left. To the right is Table I, which shows the resonance frequency of different amino acids of Zn++ (HIV1-F1) peptide.
Discussion
Berthon et al. (10) found that in Archaea, by using Genome Context Analysis, conserved clusters of genes coding for proteins involved in translation, transcription, DNA repair and ribosome biogenesis, are systematically contiguous phylogenetically. Berthon et al. also found that that MPS-1(S27E) ribosomal protein has been a component of those clusters since the appearance of Archaea on earth. Previously, Fernandez-Pol et al. found that MPS-1 is instrumental in growth regulation and carcinogenesis in animals (1, 6). Thus, these results lead to the undeniable key role of MPS-1/S27 in controlling survival events in Archaea and eukaryotic animal and plant cells (10).

Titration of FU in solution of zinc finger peptide affects NMR spectra of both the ligand and the peptide. The left panel shows aromatic 1H resonances (=ligand resonance); the right panel shows aliphatic resonances (=shifted peptide resonance).
Genomic context analysis of genomes is valuable to explore evolution of genes involved in cell division and carcinogenesis, deduce additional gene functions, and predict the functional linking between proteins belonging to such clusters (10).
Berthon et al. used all the known DNA replication genes in Archaea genomes to determine which DNA replication proteins associate, how they are involved in cell division, and if they are related phylogenetically to other species (10).
They found two distinct sets of DNA replication genes which co-localize in Archaeal genomes: (i) the first gene cluster set includes genes for PCNA, the small subunit of the DNA primase (PriS), and Gins15; the second comprises the genes from MCM and Gins23.
Unexpectedly, a conserved cluster of genes coding for proteins involved in translation and ribosome biogenesis (S27E, L44E, aIF-2 alpha, Nop10) was found almost systematically contiguous to the group of genes coding for PCNA, PriS, and Gins15. This cluster encodes proteins conserved in Archaea and enkaryotes (10).
Genome context analysis showed that a conserved cluster of ribosome genes is involved in translation and ribosome biogenesis and is systematically associated with a replisome cluster in Archaea and Eurkarya. LSIN ribosome cluster codes for the following proteins involved in translation and ribosome biogenesis: S27E(MPS-1), Nop10, αIF-2α, and L44E. PPsG replisome cluster codes for proteins involved in DNA replication and repair: PPsG: Gins 15, Pri S (small subunit of the DNA primase) and PCNA (10).
The published data leads to the proposal that RPMPS-1, and perhaps the three companion cluster RPs (31), is a unique target that when specifically inactivated by the methods described in this paper, or others (Fernandez-Pol et al., 2004), may eradicate numerous types of cancer, prevent and cure numerous viral diseases, and control other proliferative diseases, such as rheumatoid arthritis. If the diagnosis is made early and methods are developed to prevent the appearance of pathogenic mutations of RPMPS-1, numerous diseases could be prevented or cured by eliminating a single class of protein alone or in combination with the companion cluster of RPs.
The evidence available supports the contention that carcinogenesis may be avoidable at the molecular level if RPMPS-1 mutations and overexpression are detected early by the presence of RPMPS-1 in the blood or tissue samples of patients (8, 9). The Author believes that the data indicates that the chances of eradicating carcinogenesis and therefore cancer are much higher than supposed today as new targets such as RPMPS-1 have been identified.

Two dimensional NOESY spectrums indicate that FU binds the Zn finger peptide. NOE contacts of FU have the same sign as those of the peptide.
Expression of the proteome is precisely regulated at the level of protein synthesis and thus translational control is a critical homeostatic mechanism that allows the cell to respond to internal and external signals, which continuously lead to entropy and thus decay of the cellular systems.
The translation of cellular mRNA is different from that of numerous viruses. Viral messages containing internal ribosome entry site elements (IRES) located in its 5' untranslated regions are insensitive to decreases or repression of global RPs and/or translation (22-26). These alternative mechanism(s) of translation are involved in the persistent synthesis of proteins that are necessary for the cells to recover from non-lethal stress. Furthermore, the IRES system is used by numerous types of viruses, as will be described later.
In addition to RPs, expression of various eukaryotic initiation factors is elevated in experimental cancers and promotes tumor growth (27). For example, eIF4E levels increase translation of growth factors which in many instances are the driving forces of malignant tumors (12, 27). When specific antisense oligonucleotide (ASOs) are used in cancer cells in tissue culture, ASOs designed to specifically target eIF4E mRNA for elimination, result in repression of eIF4E-regulated proteins such as VEGF, c-myc, and cyclin D1 (39, 40, 41). After treatment with ASOs, apoptosis is induced and that leads to destruction of neovascularization (27-29).
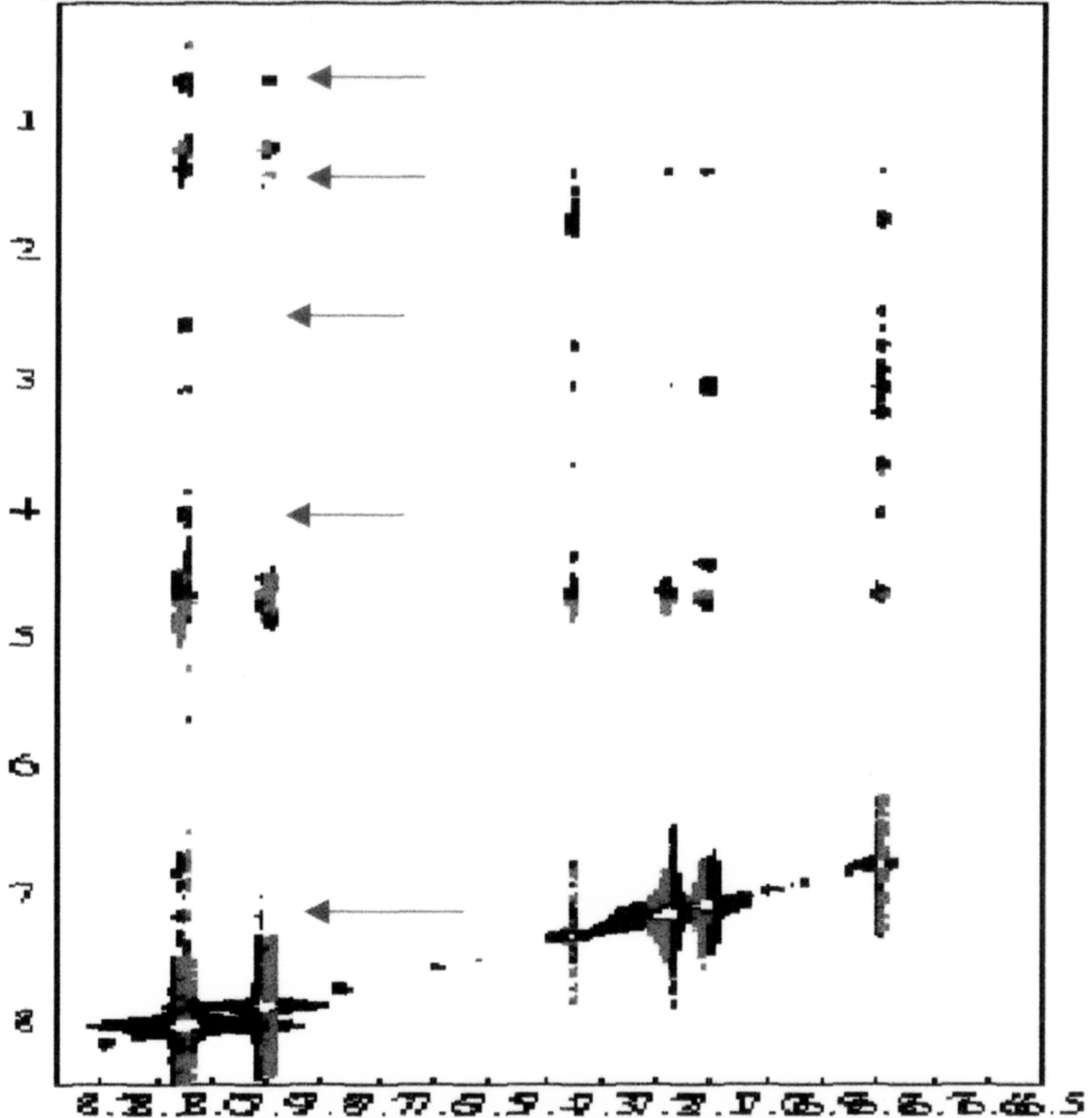
Cross peaks between FU and peptide resonances in NOESY spectrum indicate contacts in hydrophobic G-recognition site.
The nucleolus is the site of ribosomal RNA (rRNA) synthesis and assembly of ribosomal subunits. The appearance of the nucleolus in eukaryotic cells changes dramatically during the cell cycle. This is due to the fact that the components are dispersed at the onset of mitosis and reassembled at the end of cell division. When the cell initiates mitosis, the nucleolus decreases in size and then disappears; there is no nucleolus in metaphase. In telophase, small nucleoli reappear at the location of DNA regions known as nucleolar organizers.
Ribosomes catalyze protein synthesis and consist of a small 40S subunit and a large 60S subunit. The ribosome is composed of 4 different types of RNA and about 80 structurally different proteins. MPS-1/S27 gene encodes an RP that is a component of the 40S subunit. RPMPS-1 belongs to the S27E family of RPs. It is a C4-type ZFP that can bind zinc and other transition metal ions such as iron or copper. In the nucleus, the MPS-1 gene is located in chromosome 1q21 (1, 6, 7, 12, 21).
Fernandez-Pol et al. cloned the cDNA of MPS-1 gene, sequenced the gene and produced the protein in a Baculovirus expression system (6, 7). The MPS-1 protein was cloned by differential hybridization from human breast cancer cells stimulated with growth factors such as EGF plus TGFβ (TGF beta increases mRNA for the EGF receptor; 6).

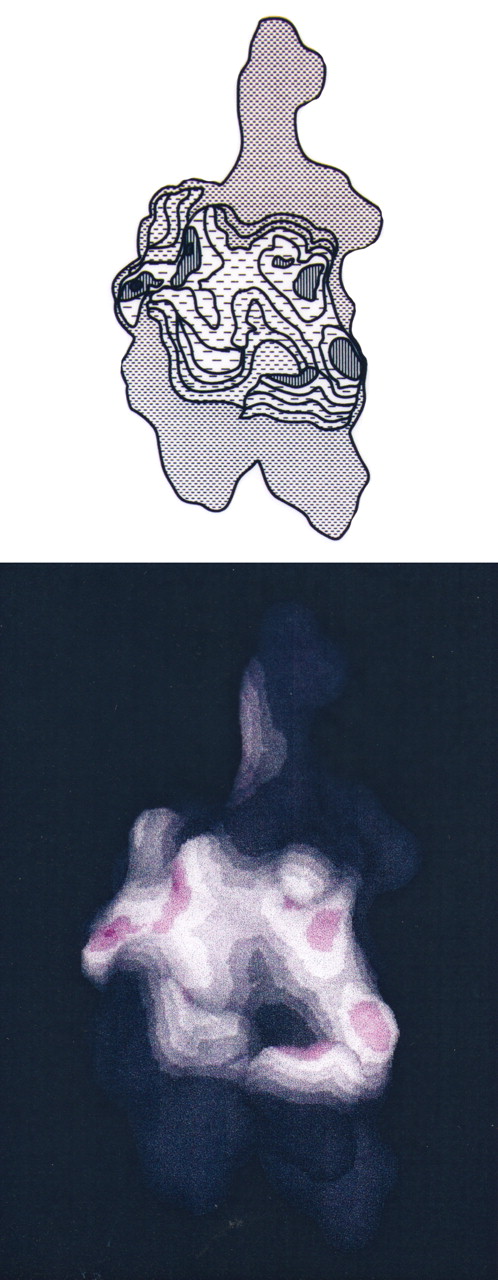
NMR structure of Zn finger peptide from nucleic acid-binding domain. This image shows the peptide alone with the RNA contact surface.

. NMR structure of Zn finger peptide from nucleic acid-binding domain. This image shows contacts between peptide and RNA base denoted G3.
Asuta et al. identified MPS-1 as a member of a tumor-associated antigen in patients with breast cancer using different molecular biology techniques to isolate MPS-1 protein and gene (30).
The encoded MPS-1 protein has been shown to be able to bind specifically to both DNA and RNA (1, 6, 7). MPS-1 binds to DNA sequences encoding the cyclic AMP response element (7). In cancer cells, when MPS-1 is overproduced, the protein can be detected in significant quantities in the culture media. In Baculovirus cells infected with viral vectors containing MPS-1 DNA, overproduction of MPS-1 leads to release of large quantities of MPS-1, in the culture media and eventually apoptosis of the transfected cells (1, 7).
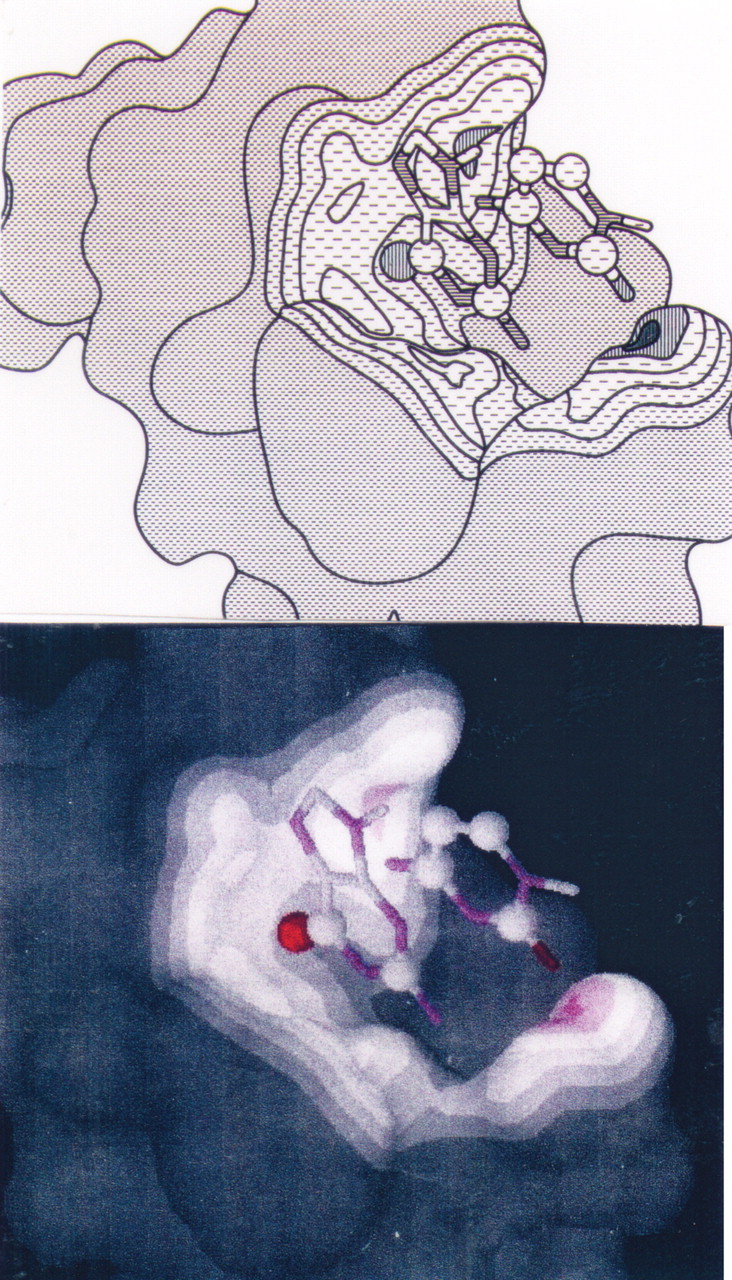
NMR structure of Zn finger peptide from nucleic acid-binding domain. This image shows the contacts between peptide, RNA base G3, and C4.
Revenkova et al. (31) unexpectedly found certain functions of MPS-1/S27) ribosomal protein in genotoxic stress responses in plants. The mutants were sensitive to methyl methanesulfonate (MMS) and to UV-C light. The T-DNA insertion disrupted one of the three genes encoding RPs MPS-1/S27 [A.B.C] in plants, one of the mutants was denoted ARS27A. The growth of ARS27A mutant was strongly inhibited by in the presence of MMS and UV-C light, in comparison with the wild-type. This inhibition was simultaneously associated with strong growth inhibition and subsequent formation of tumor-like structures. Thus, tumor growth is the result of mutation of MPS-1 (ARS27A) when Arabidopsis thaliana mutant plants are exposed to genotoxic MMS treatment or UV-C light. These results indicate that this isoform of RPMPS-1/S27 (mutant RPMPS-1/ARS27) is not required for the function of ribosomes, but is necessary for the elimination of damaged mRNA transcripts after UV irradiation (1, 12, 31).

NMR structure of Zn finger peptide from nucleic acid-binding domain. This figure shows the binding of the peptide to a region of the RNA molecule (peptide-RNA complex).
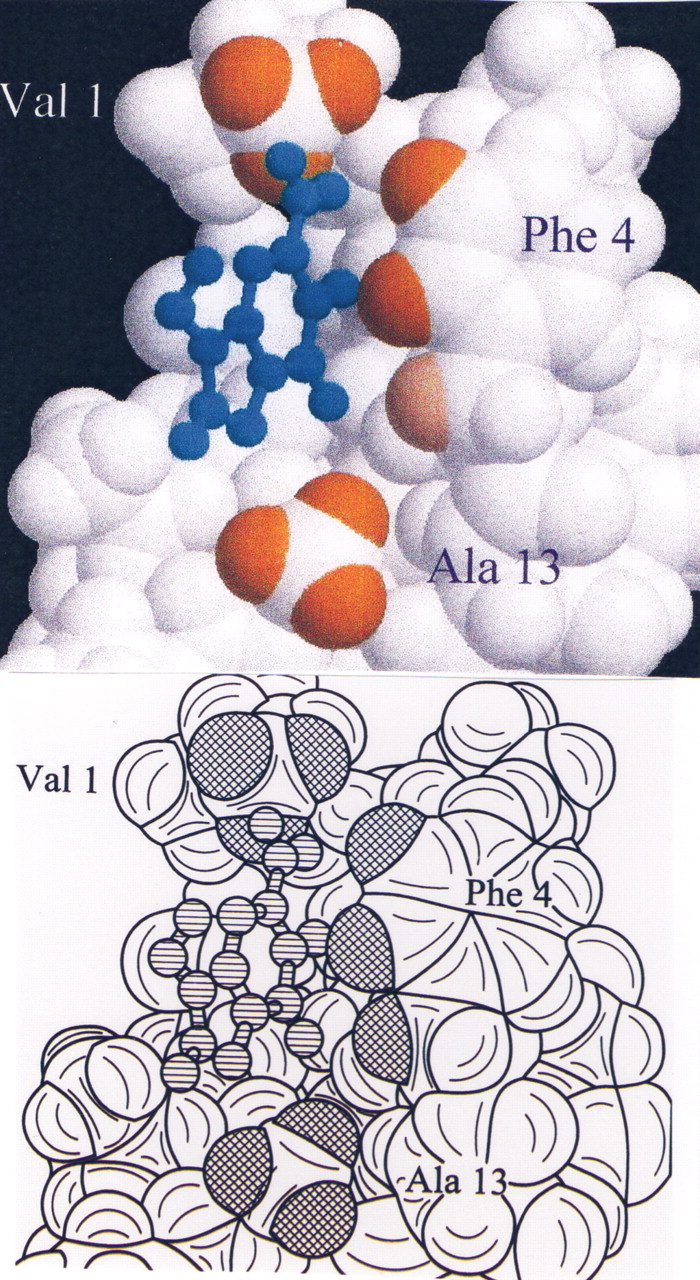
NMR structure of Zn finger peptide from nucleic acid-binding domain. Peptide 1H show NOE contacts to FU in orange (Val 1; Phe 4; Ala 13); the RNA G3 base is shown in cyan.
In vitro and in vivo, there is evidence that MPS-1 is involved in gastric cancer progression and tumorigenicity (32). Wang et al. found that 86% of gastric cancer tissues and all gastric cancer cells overexpressed MPS-1 (32). The cellular increase in MPS-1 correlated well with the tumor–node–metastasis clinical stage. Interestingly, The MPS-1 expression level was significantly decreased in cells transfected with MPS-1-specific siRNA expression of plasmid pRNAT-133. Furthermore, the transfected cells, when MPS-1 was inhibited showed an increase in apoptosis, decreased tumorigenicity, and decreased growth rate in nude mice (32).
Ribosomes, the organelles that catalyze protein synthesis, consist of a small 40S subunit and a large 60S subunit. Together these subunits are composed of 4 RNA species and about 80 structurally distinct proteins. RP S27E (metallopanstimulin 1; IPI00221096; MPS-1) is a component of the 40S subunit. The protein belongs to the S27E family of ribosomal proteins. It contains a C4-type zinc finger domain that can bind to zinc and other transition metal ions (1). The encoded protein has been shown to be able to bind to nucleic acids (12). It is located in the cytoplasm and attached at the external nuclear membrane as a component of complete ribosomes, but it is also detected in the nucleolus in large quantities in cancer cells by anti-MPS-1 antibodies (1). The cytoplasm/nucleolus fractional distribution is dependent upon the cell cycle and also upon presence of DNA damage and growth factor stimulation (1, 6, 7).
Studies in various systems indicate that RP MPS-1/S27 is located near the ribosomal protein S18 in the 40S subunit and is covalently linked to translation initiation factor eIF3 (12). As is common for genes encoding RPs (22-26), there are multiple processed pseudogenes of the MPS-1 gene dispersed through the genome (1).
The ribosome is essential for growth and development (22-26, 33, 34) and thus, in concert with hormone and growth factor stimulation they act as clocks for cellular division (1). Ribosomal defects are present in cancer cells and hereditary diseases and thus aberrations in ribosome biogenesis result in abnormal phenotypes (22-29).
In carcinogenesis, these aberrations lead to increased or decreased ribosomal biogenesis, depending upon the mutational status of the DNA (27). Increased ribosome biogenesis is associated with increased cell growth and proliferation (1, 22-29). Molecular analysis of gene expression in various types of cancer, such as ovarian, colon, pancreas, liver and viral-induced carcinogenesis, the ribosome number and many RPs and initiation factors are overexpressed, and can be used as prognostic markers for aggressiveness of cancer growth (1, 15, 34, 35).
Ribosomal mRNA translation and ribosome synthesis are tightly connected to cell cycle control (1, 33, 35). Overexpression of RPs in carcinogenesis must be a direct consequence of oncogene activation or tumor suppressor gene neutralization (1). Overexpression of RPs may have a direct consequence of permanently stimulating cell cycle progression resulting in uncontrolled cell proliferation. Compelling evidence that RPs are important for both controlled and uncontrolled growth is the result of numerous studies (1, 22-29). Overexpression of RP genes observed in tumors appears to be the result of increased translation of individual mRNAs rather than in up-regulation of global protein synthesis, as demonstrated by studies of growth stimulation with specific growth factors in cancer cells, differential hybridization of mRNA from quiescent cells and growth factor-stimulated cells (1, 7, 11, 22-29). Fernandez-Pol et al. showed that only about 140 proteins are required for cancer cell cycle progression and of these, about 15 RPs are consistently increased (1, 11, 6). It is worth mentioning here that the number of ribosomes is increased in numerous cancer cells when compared to normal cells (1, 35). Progressively reduction of MPS-1/S27 RP synthesis is observed in normal senescent cells (11). Reduction of MPS-1 and other RPs is also observed in many cancer cells, but this may be due to the large number of mutations in MPS-1 and other RPs that are present in advanced carcinogenesis (1). Thus, increases in ribosome biogenesis have been correlated with malignant transformation (1, 22-30).

The Flaviviridae family comprises three genera, Hepacivirus, Pestivirus, and Flavivirus. They contain a single strand of (+) positive-strand RNA that has mRNA activity. The C-terminal region of the NS5 protein contains five amino acid sequence motifs that form the signature of RNA-dependent RNA polymerases (RdRps). Dengue virus, hepatitis C and bovine viral diarhhea virus all contained such structure. A and B represent a model for Dengue virus NS5 RdRp, which is a combination of the P6 polymerase complex and the Hepatitis C virus complex with ribonucleotides.
Taken together, the results from several laboratories in the area of growth and carcinogenesis suggest a previously unrecognized regulatory network coupling DNA replication and translation in Archaea that also exist in enkaryotes (10).
We have also found by cloning of growth factor-stimulated animal cancer cells that some translation initiation and elongation factors can be directly involved in carcinogenesis (12).
The Author suggests that mutations in components of the ribosomal-translational apparatus are critical for the development of carcinogenesis in eukaryotes and that certain RPs may be used as a targets for highly specific anticancer agents. The widely conserved clustering of a particular set of genes encoding ribosomal proteins (L44E, S27E (MPS-1); aIF-2 alpha, Nop10) and the studies with MPS-1 in animal and plant eukaryotic cells suggest that MPS-1 has conserved roles that involve translation as well as ribosome biogenesis, transcription, and DNA repair (1, 7, 10). Furthermore, we have shown that in association with increases in MPS-1 after growth factor stimulation (6, 7), there is an increased of numerous RPs as well as initiation factors and the entire set of mitochondrial DNA coded proteins (12).
Certain drugs have been used to study regulation of rRNA and mRNA metabolism, but these drugs generally affect systhesis and processing of rRNA and mRNA, making them non-specific. The unique effects of two drugs, PA and 5-methylnicotinamide, on rRNA metabolism was first reported by Constantini and Johnson. (34). Treatment with these agents results in an inability to accumulate 28S rRNA and 60S ribosomal subunits in the cytoplasm. Treatment with PA results in fragmentation and disorganization of the nucleolus but with preservation of both fibrillar and granular components. Similar changes are observed in virtually all cells exposed to PA (34). These results, taken together with the antiviral effects of PA on viruses utilizing ZFPs for replication, some of them illustrated in Table II, suggest that the unique actions of PA in the nucleolus (34) may be an additional instrumental factor in the antiviral effects of PA and FU on numerous viruses, in addition to the effects of PA on viral ZFPs.
One interesting aspect of viral protein synthesis is the interaction of (+) mRNA viruses with specific ribosomal proteins. RNA viruses interact with the nucleolus to control host-cell functions and recruit nucleoli proteins to replicate (1, 22-29). For example, the HIV-1 virus directs the HIV-1 Rev protein to the nucleolus (1). Through interaction with the B23 nucleoli protein, it regulates the splicing of the HIV-1 mRNA. The disproportionate accumulation of 18S and 28S ribosomal RNA and the specific impairment in rRNA processing in cells treated with PA or 5-methylnicotinamide, is a unique action of PA and 5-methylnicotinamide (34). No other drugs have been found to have such effects (34). Thus, the conditions of imbalance in nucleoli rRNA induced by PA may be involved in the antiviral activity of PA in numerous viruses that utilize the nucleolus to replicate.
Increased nucleoli activity or deregulation of a few ribosomal proteins will result in overproduction of ribosomes or specific ribosomal proteins such as MPS-1/S27, which will eventually lead to uncontrolled cell growth (1, 10, 22-29).
By NMR and other techniques, the potential mechanism by which the CCHC zinc finger domain of retroviral nucleocapsid proteins Np7 and other ZFPs such as MPS-1/S27E are neutralized and its function disrupted by picolinic acid and derivatives thereof have been elucidated. The NMR data shown in Figures 16, 17, 18, 19, 20, 21, 22, 23, demonstrates how FA inactivates the zinc finger domain of the nucleoprotein Np7 of the HIV-1 virus. The same compounds inactivate MPS-1 by interacting with the ZFP-binding domain CCCC(1). Several classes of compounds have been discovered which can be used to inactivate HIV-1 viruses by attacking the CCHC or other type of zinc finger domain (Fernandez-Pol et al. US patent 2004).
ZFPs are extensively used for various essential functions by many families of pathogenic DNA or RNA viruses (1). Table II illustrates several examples of families of viruses using ZFPs or transition metal ion-dependent enzymes for replication and/or virulence that can be targeted with picolinic acid, fusaric acid or derivatives thereof. Other viral ZFP-disrupting agents, such as 2-thiophene-carboxylic acids and derivatives thereof, are also effective antiviral agents (Fernandez-Pol et al., US patent 2004).
One example of the importance of viral ZFPs is illustrated by the case of influenza A virus genome RNA and its encoded proteins (1). The encoded polypeptide M1 is a ZFP and thus a potential target for PA, FU and derivatives thereof. M1 gene is located in segment 7, contaminating 1027 nucleotides which encode the M1 polypeptide (252 amino acids), with a molecular weight of 27 kDa. The copy number per virion of the M1 ZFP is approximately 3,000 and thus is a major virion component involved in RNP transport out of the nucleus. These data suggests that targeting the M1viral ZFP may destroy the influenza A virus integrity and replicative capacity. If in addition and simultaneously, the host cell RPMPS-1/S27 is eliminated, which is disposable for the host, but critical for virus replication, including influenza A virus (25), the elimination of both M1 and MPS-1 proteins could lead to viral suppression, viral degradation, and subsequent prevention of influenza virus infection or disease progression (1).
RNA virus macromolecular synthesis and viral genome strategies. RNA viruses have adopted a number of different strategies for expression of their genes and the replication of their genomes (Figure 24). There is a wide-spread class of single strand (+) RNA viruses that utilize IRESs for translation and viral replication (22-26). They include Picorna viruses, Togaviruses and Flavivirus (e.g.: hepatitis C (HCV) and Dengue viruses) contain a single strand of positive (+)-strand RNA that has mRNA activity. Genome-wide RNAi screening has revealed specific sensitivity of IRES-containing RNA viruses to host translation inhibition. Cherry et al. performed a genome-wide RNAi screen in Drosophila cells infected with Drosophila C virus (DCV) (26). They identified 66 RPs that, when reduced or depleted, specifically inhibit DCV growth. However, non-IRSs-containing RNA viruses remained unaffected. Furthermore, treatment of Drosophila with a translation inhibitor was shown to be protective in vivo. Similar increased sensitivity to RP levels prevented infection of human cells with poliovirus, indicating the universality of these findings (26). Thus, these authors have found that viruses that are (+) sense RNA and that use IRES, require components of the ribosome. They have shown that when RpS27 (MPS-1) is knockdown, the production of the DCV is inhibited and thus, replication of this virus is abolished (26).
A few examples of families of viruses using zinc finger proteins or transition metal ion-dependent enzymes for replication and/or virulence that can be targeted with the agents of this paper.
Potential mechanism of inhibition of PA, FU and derivatives thereof on positive (+) single-strand RNA human virus replication. There is a wide-spread class of single-strand (+) RNA viruses that use IRESs for translation and viral replication. They include Picorna viruses, Togaviruses and Flavivirus (HCV and Dengue viruses). All these viruses contain a single strand of positive (+)-strand RNA that has mRNA activity. The strategies utilized by single-stranded RNA virus replication are summarized in Figure 24.
When culture cells infected with human hepatitis C virus (HCV) are treated with PA, FU, or derivatives thereof, the (+) mRNA single strand of HCV is destroyed as demonstrated by RT-PCR (Figure 13). RPs are also reduced or depleted, including RPMPS-1/S27, suggesting that inhibition of HCV is unable to replicate as a result of at least two events: i) degradation of (+) single-strand mRNA for HCV and inhibition of RPMPS-1/S27 which is decreased after treatment with PA, FU and derivatives thereof. Similarly, pretreatment with PA and derivatives thereof results in a decreased in ribosomal protein levels which prevent infection of human cells with HCV. BDV, and Sindbis virus, where also inhibited by PA and derivatives thereof indicating the potential universality of these findings. Thus, we have found that viruses that are (+) sense RNA and that have been shown to use IRES depend upon components of the ribosome for replication. The results also suggest that when RPMPS-1/S27 is disrupted, the production of HCV, BDV and Sindbis virus is inhibited and thus, replication and infectivity of these virus is abolished. The results with HCV, BDV and Sindbis virus treated with PA and FU, discussed above, are illustrated by the experimental findings shown in Figures 12, 13 and 14.
Amsterdam et al. (22) studied numerous RP genes in Zebra fish and they found that many RP genes may act as tumor suppressor genes in fish. Many RP genes might also be genes involved in carcinogenesis in humans (22). The method used by these authors is not genome-wide and they found the role of a number of RP by chance, in the process of studying heterozygous fish. They found early mortality in an RP mutant line which resulted from multiple types of cancer. MPS-1 (S27E was not found in these narrow studies. However, they found S18, which is adjacent to MPS-1 and S29 which is an RP protein with one zinc-finger motif (-C2-X-C2-) similar to that of MPS-1/S27 (-C2-X-C2-). Thus, S29 has the potential for binding to bind nucleic acids. These motifs also bind to RNA, but the exact details are not known. Interestingly, the human ubiquitin carboxyl-terminal extension protein CEP80 is ribosomal protein S27a, but its function in carcinogenesis in humans remains unclear.
Volarevik et al. (24) have shown that the ability of liver cells to grow and their ability to proliferate can be dissociated by conditionally deleting the S6 gene, which is one of 33 40S ribosomal protein genes, in the liver of adult rats. They found that proliferation but not cell growth can be blocked by conditional deletion of 40S ribosomal protein S6. Considering that proliferation of eukaryotic cells expends about 80% of its energy on the biogenesis of RPs, S6 protein may be a good target for arresting cell growth with specific pharmacological agents.
Giaever et al. (23) have determined the effect of genome-wide deletion to understand gene function. They found that when ribosomal genes of Saccharomyces cerevisiae are eliminated, the replicative capacity of this organism is decreased, but the deletion is not lethal.
Different families of viruses have developed different strategies in the host cells (to use host-cell proteins) for replication (of viral proteins) and virion assembly and all viruses must use the host-encoded translation system, which is the complex ribosomal machinery that is exploited by the different viruses to be able to translate their genomes (1; Figure 24).
Several families of viruses have by-passed the 5' m Gppp cap, which is a characteristic of host cell mRNAs, and instead utilize a unique viral RNA sequence, which allows cap-independent utilization of ribosomes to translate the viral RNA (25). This specific viral mechanism for translation of viral proteins suggests that specific inhibitors of IRES-dependent translation can be used to prevent or arrest viral infections.
Many human pathogenic viruses using the IRES mechanism, such as Flaviviruses which include Togaviruses, HCV, and Dengue viruses, may be susceptible to block replication of these types of viruses by specific anti-IRES-antiviral agents. We suggest that inhibition of MPS-1/(S27E) RP may result in RP depletion, which will not affect normal cells, and simultaneously, the cells may block viral infection by viruses that utilize IRES strategies (Figure 24).
It should be emphasized here that ribosomal proteins are globally decreased when one single ribosomal component is blocked or deleted (22-29). In normal cells, this type of inhibition is tolerated with minimal or no toxicity when picolinic acids and certain derivatives thereof are utilized. Normal cells when substantially depleted of ribosomal proteins by the use of PA or FU may be protected from the family of viruses mentioned above as well as other viruses which act by different mechanisms. Finally, the fact that inhibition of MPS-1/(S27E) can be specifically inhibited by PA, FU and derivatives thereof, indicates that modulating or inhibiting MPS-1 with the purpose of prevention or therapy of specific viral diseases, such as HCV or Dengue (40-41), may lead to effective antiviral prevention and treatment of viral diseases of humans and animals susceptible to (+) single-strand RNA viruses.
We have shown that pharmacological control of MPS-1/S27 can be achieved by different means (1). Due to their low toxicity in animal experiments, we have focused on agents derived from PA, a naturally occurring substance able to neutralize MPS-1/S27 and induced virally-transformed cells to enter into apoptosis (1, 3).
Viruses can be classified by genomic composition and pathways of mRNA synthesis. The Flaviviridae family comprises three genera, Hepacivirus, Pestivirus, and Flavivirus. They contain a single strand of (+) positive-strand RNA that has mRNA activity (Figure 24).
The C-terminal region of the NS5 protein contains five amino acid sequence motifs that form the signature of RNA-dependent RNA polymerases (RdRps) (40, 41). Dengue virus HCV and BVDV all contained such structure. Figure 24, (A and B), represents a model for Dengue virus NS5 RdRp, which is a combination of the P6 polymerase complex and the HCV complex with ribonucleotides. The model (A and B) was created by Yap et al. (40). Figure 24 (A), shows NS5 which contains two zinc ion-binding motifs; and Figure 24 (B), shows the NTP entry channel. (Figure 24 data obtained from RCSB Protein Data Bank; P299991; and Ref. 40).
Flaviviridae are viruses with positive-strand RNA genomes that have been classified in three genera, Hepacivirus, Pestivirus, and Flavivirus. The genomic RNA of Dengue virus is translated into a single polyprotein which is cleaved, and seven nonstructural proteins (NS1-NS2A-NS2B-NS3-NS4A-NS4B and NS5) which are cleave by viral and cellular proteases. NS5 is the most conserved viral protein of Dengue virus (41). The C-terminal region of NS5 RdRps contains five motifs that characterize, specific assays for which are used for HCV and BVDV and Dengue virus.
All these polymerases share an architecture composed of palm, fingers, and thumb domains and a common catalytic mechanism that use two zinc atoms (37). These structural and mechanistic identities allows, for example, testing the efficacy of antiviral agents against HCV by performing efficacy testing against BVDV as a surrogate assay for HCV. Comparative studies suggest that HCV is closely related to Dengue virus, and more closely to pestiviruses, such as BVDV.
BVDV is used as a model for HCV due to the difficulty in culture of HCV, and BVDV resembles HCV-persistent infection. The antiviral effects of PA and FU on BVDV are shown in Figure 12.
We have also determined the effects of PA and FU on: (i) HCV and Sindbis replicons (by qRTPCR/icycler) (Figures 13 and 14); and (ii) ava5 and Huh7b/Sindbis cells (MTS). The results showed that after treatment of infected cells for 3 days with PA at a concentration of 250 μg/mL, BVDV virus yield was reduced by 76% (EC50=60 μg/mL. These indicate that both PA and FU are active against BVDV (Figure 12).
Figure 13 shows the results of RT-PCR experiments with human HCV treated with PA and FU. The results clearly indicate that PA and FU are antiviral agents able to treat HCV infections in tissue culture conditions.
Figure 14 shows the Sinbis replicon data for both PA and FU at various concentrations. The results show greater than 90% inhibition of Sindbis replicon at all concentrations of PA tested. By OD, no overt cytotoxicity was observed. FU caused >85% inhibition of Sindbis replicon at all concentrations tested. No overt cytotoxicity was observed by OD.
PA has macrophage-stimulatory activities (35-39). Studies performed by the group of Varesio et al. have demonstrated that PA can act as co-stimulus for macrophages to enhance their antitumor activity in animals (35-39). Furthermore, PA in conjunction with IFN-gamma inhibits retrovirus production by macrophages and simultaneously induces the expression of the nitric oxide synthase gene (36, 38-39). PA also protects mice against lethal infections with the pathogen Candida albicans and induces production of numerous cytokines (1, 35). Extensive information indicates that PA can be used pharmacologically to enhance and/or modulate macrophage defensive functions (35-39). Thus, antiviral effects of PA, or derivatives thereof, beyond direct cytotoxicity towards virus infected cells, are mediated by macrophage activity.
PA is used as a dietary supplement to carry transition metal ions such as zinc, iron, copper or chromium and these compounds are widely distributed and consumed by humans (1, 42). The recognition of important physiological, pharmacological, and nutritional effects of PA as carrier of transition metal ions led to studies on PA in biological fluids (1, 42). PA concentrations are difficult to measure because PA forms complexes with metalloproteins in vitro (1, 42). When specific techniques for measument of PA and derivatives thereof were created, PA was detected in tissue culture supernatants, serum, cerebrospinal fluid, human milk, pancreatic juice, and many different animal tissues (1, 42).
In summary, RPMPS-1/S27 and other RPs (e.g. S29) are being used as a target for some novel low toxicity antiviral agents such as PA, FU, or derivatives thereof. These therapies aim to eject the zinc from the ZFP motifs of the RPMPS-1, thus rendering it inactive or to form ternary complexes which result in degradation of RPMPS-1. These therapies have shown promise for the treatment and prevention of carcinogenesis and viral diseases in laboratory experiments. Our efforts with viral ZFPs and RPMPS-1/S27 have identified several novel classes of potent agents chemically unrelated to PA, FU, or derivatives (Fernandez-Pol et al., US patent, 2004), that are undergoing evaluation as non-toxic and potentially highly effective anti-viral agents inhibitors of viral ZFPs and RPMPS-1/S27. Figure 24 illustrates RPMPS-1/S27 as a target for several families of viruses and in particular for viruses of the Flaviridae family which contain a single strand of (+)-positive strand RNA that has mRNA activity. Figure 24A and B represent a model for DENV NS5 RdRp protein: A: NS5 contains two zinc ion-binding motifs; B: NTP is the entry channel for single strand (+) viral mRNA that potentially can interact with RPMPS-1/S27 and RPS29 in the channel, where they are closely related. (Model adapted for the mechanism of action of PA and FU from RCSB Protein Data Bank; P299991; and Ref. 40). Finally, the antiviral effects of PA, FU and derivatives thereof, beyond direct destruction of viral ZFPs and mutated RPMPS-1 in virally infected cells, are mediated by activation of macrophage activity.
Acknowledgments
This work was supported by Antagoras Agrobusiness, LLC., St. Louis, MO, USA, designated funds.
- Received February 8, 2011.
- Revision received February 27, 2011.
- Accepted February 28, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Host genetic variants regulates CCR5 expression on immune cells: a study in people living with HIV and healthy controls
- A Novel Marker for Purkinje Cells, Ribosomal Protein MPS1/S27: Expression of MPS1 in Human Cerebellum
- Increased Serum Level of RPMPS-1/S27 Protein in Patients with Various Types of Cancer Is Useful for the Early Detection, Prevention and Therapy