


Abstract
Background: Breast cancer is one of the most frequent tumors worldwide. Currently applied diagnostic approaches are frequently not able to recognize early stages in tumor development therefore impairing outcomes. The focus of this study is the creation of a non-invasive predictive diagnostic approach by pathology-specific blood proteome analysis. Patients and Methods: Circulating leukocytes were isolated from fresh blood samples of breast cancer patients, benign breast pathologies and healthy controls. In patients with all kinds of breast pathologies, blood samples were taken before core needle biopsy of the lump. Comparative protein mapping was performed by 2D-PAGE followed by MALDI-TOF analysis and Western-blot quantification of differentially expressed protein spots. Results: By protein mapping, 64 protein spots were identified. Pathology-specific differential expression patterns comprised microfilamental network-associated proteins: Calgranulin A (S100), LyGDI (Rho GDIβ), RhoA and profilin 1. RhoA and profilin values discriminated between healthy controls and patients with all breast pathologies. Conclusion: Microfilamental network-associated proteins are involved in the regulation of a variety of central cellular processes functionally linked with each other and known to be highly relevant for all stages of tumorigenesis including precancerous lesions and metastases. Pathology-related molecular patterns are currently considered for the creation of a novel highly sensitive minimally-invasive approach for predictive diagnosis of breast cancer.
- Breast cancer
- clinical proteomics
- microfilamental network
- disease-specific molecular patterns
- predictive diagnosis
Abbreviations: 2D-PAGE: Two-dimensional polyacrylamide gel electrophoresis; IPG: immobilized pH gradient; DTT: dithiotreitol; SDS: dodecyl sodium sulphate; MALDI-TOF: Matrix-assisted laser desorption/Ionisation - Time-of-flight; Mr: molecular weight; pI: isoelectric point; ACN: acetonitrile; CHCA: alpha-cyano-4-hydroxycinnamic acid; TFA: trifluoroacetic acid; ATP: adenosine triphosphate; NADH: nicotinamide adenine dinucleotide.
Breast cancer is the most common cause of cancer death among women. In 2005, breast cancer led to 502,000 deaths (7% of cancer deaths; almost 1% of all deaths) worldwide (1). Early diagnosis has been demonstrated to be highly beneficial for significantly enhanced therapy efficiency and possible full recovery. During the past three decades, the incidence of breast cancer has continually increased worldwide. Although population screening by mammography and application of adjuvant therapies have kept breast cancer mortality mostly unchanged or even persistently declining over the last ten years (2), currently applied diagnostic approaches, however, are frequently not able to recognize very early stages in tumor development results in the outcome. Although breast MRI is currently the most sensitive diagnostic tool for breast imaging, its specificity is limited resulting in negative impact for surgical management in approximately 9% of cases (3).
Identification of the affected pathways would significantly improve the understanding of molecular pathomechanisms of breast cancer and provide a fundamental platform for the creation of more specific diagnostic technologies and more effective targeted therapies. During the past three decades, much effort has been put into the investigation of molecular pathways in human breast cancer tissue and cell lines by ex vivo and in vitro approaches, respectively, as well as using corresponding animal models. An important data pool has been collected at the level of DNA (particularities of epigenetic control, pathology-related mutations), cancer-specific changes in transcriptional and translational gene expression levels, altered post-translational protein-modification, potential therapy-targets, etc. as reviewed by Ross et al. in 2005 (2). Advanced biotechnologies such as the “-omics” have been applied. In terms of potential diagnostic applications, however, both in vitro conditions of cell lines and animal models are unable to simulate in vivo conditions of human breast cancer adequately. The big issue-related disadvantage of utilizing the biopsy samples is generated by the mixed cell population, which strongly varies from case to case resulting in differential gene expression with low pathology-related specificity.
Current efforts are focused on the creation of precise pathology-specific diagnostics with predictive power in terms of individual predisposition to breast cancer as well as personalized therapy planning based on an application of advanced minimally-invasive molecular biotechnological approaches such as pathology-specific proteome of blood cells (4-6). The central idea behind this approach is molecular imaging of individual stress response towards a variety of exogenous and endogenous factors. Stress response depends on the currently available individual stress proteome, which consequently regulates the majority of down-stream molecular and cellular processes such as energy metabolism, DNA-repair, cellular proliferation and death, i.e. clue events in both physiologic cellular activity and carcinogenesis. Therefore, the same central molecular players are involved in both physiologic and pathologic processes, which, however, dramatically differ from each other in specific expression patterns as it has been clearly demonstrated for cell cycle checkpoints and related genes (7). In damaged cells, an inadequate regulation of this molecular pathway leads to plenty of pathologies including senescence, degeneration and the most frequent causes of human morbidity and mortality - cardiovascular diseases and cancer. A growing body of evidence supports these conclusions. Thus, diabetic patients suffering from highly increased stress factors by impaired glucose/insulin-metabolism have been demonstrated to be predisposed to cardio-vascular pathologies, dementia, pronounced degenerative processes, and several malignancies including breast cancer (8, 9).
The important mission of clinical proteomics is to identify the protein expression patterns specific for each kind of above mentioned pathologies. Identified for pre-cancerous lesions, those specific patterns would serve as a powerful tool for predictive diagnosis followed by targeted preventive measures. This task was followed in the presented experimental strategy to favor its potential application in the predictive diagnosis of breast cancer. A minimally-invasive molecular biotechnological approach of differential proteomics in blood cells has been applied.
Patients and Methods
Recruitment of patients and controls, blood sample collection. Twenty nine patients with breast tumors and 9 controls with no history of breast pathology were recruited at the “Centre of Excellence for Breast Cancer Research”, Rheinische Friedrich-Wilhelms-University of Bonn. According to the diagnosis, the recruited patients were grouped as follows: invasive lobular carcinoma (9 patients of group A1); invasive ductal carcinoma (10 patients of group A2); and benign breast pathologies (10 patients of group B1). Data of patients and controls recruited are summarized in Table I, where all benign breast pathologies of the B1 group are listed. Healthy controls (group B2) were selected by negative breast examination at the Department of Radiology. Blood samples of all patients with breast tumors were taken prior to core needle biopsy at the Department of Obstetrics and Gynecology. All study participants were informed about the purposes of the study and signed their consent of the patient. All investigations conformed to the principles outlined in the Declaration of Helsinki and were performed with permission by the responsible Ethic's Committee of the Medical Faculty, University of Bonn.
Isolation of leukocytes. Each freshly collected blood sample underwent an immediate isolation of circulating leukocytes. Blood samples (20 mL) anti-coagulated with lithium-heparin were collected from recruited patients. Leukocytes were separated using Ficoll-Histopaque gradients (Histopaque 1077, Sigma-Aldrich, USA) as described elsewhere (10), frozen as dry pellets and processed for complete molecular biological analysis.
Two-dimensional Polyacrylamide Gel Electrophoresis (2D-PAGE). Altogether twelve 2D-PAGE-images were performed for expression patterns in circulating leukocytes of six breast cancer patients and six healthy controls. A 200 μg aliquot of each protein sample was used for 2D-PAGE analysis, which was performed twice for each sample. First-dimensional separation was performed in immobilized pH gradient (IPG) strips (Bio-Rad, USA) in the range of IP 4-7 as described by the supplier. Protein samples of 125 μL containing re-hydration buffer (8 M urea, 10 mM DTT, 1% CHAPS, 0.25% Bio-Lyte, pH 4-7) were loaded onto the IPG-strips and subjected to 14 kVh overnight at 20°C in a PROTEAN IEF Cell (Bio-Rad). Following first-dimensional separation, the extruded IPG-strips were equilibrated in gel equilibration buffer I (50 mM Tris-HCl, 6 M urea, 30% glycerol, 2% SDS, 1% DTT), followed by equilibration in buffer II (50 mM Tris-HCl, 6 M urea, 30% glycerol, 2% SDS and 260 mM iodacetamide) for 10 min before loading them onto polyacrylamide gels (12% SDS-PAGE) for the second-dimensional resolution in Mini-PROTEAN 3 (Bio-Rad). After electrophoresis, the separated proteins were visualized using silver staining (Silver Stain Plus™; Bio-Rad). Differential gene expression was analyzed using specific software (Bio-Rad).
Matrix Assisted Laser Desorption/Ionisation - Time Of Flight (MALDI-TOF). Selected spots were cut out from gels. Sample preparation was performed as described elsewhere (11). Proteins were in-gel digested by incubating the samples with porcine trypsin (Promega, USA) at 37°C overnight. Peptide mixtures were then purified by ZipTip C18 according to the manufacturer's instructions (Millipore, USA). Elution was performed using 50% ACN/water solution saturated with CHCA. A total of 1 μL of each sample was spotted onto sample anchor and allowed to dry at room temperature. After drying, 0.7 μL of re-crystallization solution (ethanol: acetone: 1% TFA in the ratio 60:30:10) were added. Peptide mass fingerprint (PMF) was performed using a MALDI-TOF mass spectrometer (Bruker Daltonics, USA) operated in positive ion reflector mode with an acceleration voltage of 25 kV. For internal calibration the trypsin autolysis peptides were applied. Mass spectra were analyzed automatically using Bruker software. The following search parameters were set: a) mono-isotopic masses, b) mass tolerance of 50 ppm, c) one missing cleavage per peptide, and d) possible oxidation of methionine-residues. No restrictions on Mr or pI were made. A minimum of four matching peptides covering at least 15% of the overall sequence were required for protein identification. The sequence similarity search was carried out using BLASTP software.
Patient and tumor characteristics. *Patients with the following benign breast pathologies have been selected for the group B3: one patient with fibroadenoma, three patients with fibrocystic disease, one patient with adenosis, two patients with lipoma, one patient with trauma, one patient with ductal hyperplasia, and one patient with phylloides tumor.
Western blot analysis. All analyses were performed twice for each sample. Primary anti-body incubation was performed at room temperature using a 1:200 dilution of the specific goat polyclonal IgG anti-bodies: calgranulin A (sc-8112; Santa Cruz, USA), Ly-GDI (sc-6047; Santa Cruz), profilin-1 (sc-18346; Santa Cruz), RhoA (sc-418; Santa Cruz) and actin (sc-1616; Santa Cruz). The protein-specific signals were measured densitometrically using the Quantity One® imaging system (Bio-Rad, USA). The expression rates of the target genes were normalized through the corresponding expression rates of actin (the house-keeping gene).
Statistical analysis. Statistical analyses were carried out using SPSS 14.0 software (SPSS, Chicago, IL, USA). Descriptive statistics of protein expression levels are reported as median and the standard deviations of 25% - 75% . To examine the significance of protein expression differences among groups of comparison, the non-parametric Mann-Whitney U-test was applied. The level of significance was chosen at p<0.05. To analyze interrelations among single proteins in groups of comparison, Spearman rank correlations were calculated.
Results
Protein mapping in circulating leukocytes of breast cancer patients. Protein mapping (Figure 1) resulted in 64 identified spots which are listed in Table II. The identified proteins belong to the following general functional groups: 1. microfilamental network-associated proteins related to cell-migration (fibrinogen, vimentin, plastin, tropomyosin, actins, tubulin, keratin, profilin, vinculin, twinfilin, calgranulin, RhoA, LyGDI, GDI2); 2. energy metabolism and transfer (mitochondrial ATP-synthase, cytochrome b, pyruvate kinase, carbonic anhydrase, phosphoglycerate kinase & mutase, mitochondrial NADH-ubiqinone oxidoreductase, triose-phosphate isomerase, lactate dehydrogenase); 3. Ca2+-dependent membrane-architecture and -signaling proteins (annexins, chloride intracellular channel protein 1); 4. stress-response, anti-oxidant defense and detoxification proteins (14-3-3 protein, heat-shock proteins, endoplasmin, catalase, peroxiredoxin, glutation S-transferase, purine nucleoside phosphorylase, alpha-1-antitrypsin, delta-aminolevulinic acid dehydratase); 5. regulatory protein-synthesis & -modification enzymes (mitochondrial Elongation factor Tu, protein disulfide-isomerase, cytosol aminopeptidase, peptidyl-prolyl cis-trans isomerase, proteasome activator); 6. tissue-remodeling enzymes (matrix-metalloproteinase-9).
Differential expression of microfilamental network-associated proteins. Differential proteomics carried out for breast cancer patients versus healthy controls demonstrated significant alterations particularly stable for protein spots related to microfilamental network-associated proteins. Regarding their functions related to a cell-migration potential, the following individual proteins were quantified by “Western-blot”-analysis: Calgranulin A (S100), LyGDI (Rho GDIβ), RhoA, profilin 1 and actin (house-keeping gene). Figure 2 (A-E) demonstrates expression levels of individual proteins calculated for the groups A1, A2, B1 and B2. Relations among corresponding medians with calculated significance of differences are represented in Figure 2F and Table III, respectively. To identify potential trends in differential regulation of individual genes, the threshold for significance was put as p<0.1. Correspondingly, significant down-regulation of calgranulin, RhoA, profilin and actin was registered for breast cancer patients (group A, subgroups A1 and A2) as well as patients with all breast pathologies (group C) versus healthy controls (group B2). No pathology-specific alterations in LyGDI regulation were monitored.

Protein mapping in circulating leukocytes of breast cancer patients: protein products for altogether 64 spots were consequently identified by MALDI-TOF analysis. The list of the identified proteins is given in Table II.
The calculated expression levels normalized by corresponding expression rates of actin as the house-keeping-gene are given in Figure 3 (A-D). Relations among corresponding medians with calculated significance of differences are represented in Figure 3E and Table IV, respectively. These results demonstrate a significant down-regulation of RhoA and profilin in breast cancer patients versus healthy controls. Medians for calgranulin expression discriminated well between invasive lobular carcinoma and all other groups. Furthermore, normalized individual expression values for calgranulin, profilin and LyGDI were highly unified (the lowest standard deviations) in patients with invasive lobular carcinoma versus all other groups tested, who demonstrated remarkable deviations towards a corresponding median. A significant correlation between expression levels of individual proteins is demonstrated in Table V.
Sixty four spots have been identified by protein mapping to be stably (i.e. by all members of the group) expressed in circulating leukocytes of the group with breast cancer patients. The image is demonstrated in Figure 1. The corresponding accession numbers and names of the identified proteins are listed here.
In addition, age was considered as a potentially contributing factor in terms of gene regulation. Indeed, the age-dependent down-regulation of expression levels was demonstrated for RhoA, actin and profilin. Considering the normalized expression values, a significant age-related down-regulation of expression levels was demonstrated for RhoA only.
Discussion
The architecture of the microfilamental network plays a key role in a number of central cellular functions such as proliferation, differentiation, adhesion, cell-cell interactions, motility and migration. Members of the network are functionally linked with each other and are known to be highly relevant for all stages of tumorigenesis starting with precancerous lesions and including enhanced metastasizing potential of malignancies. In particular, the increasing metastatic potential of tumor cells is associated with loss of actin and poorly arranged actin-skeleton organizations (12). Moreover, reorganization of the actin cytoskeleton induced by a taxoid disruption of the microtubule equilibrium has been demonstrated as a part of chemotherapeutic mechanisms of anti-cancer drugs (13).

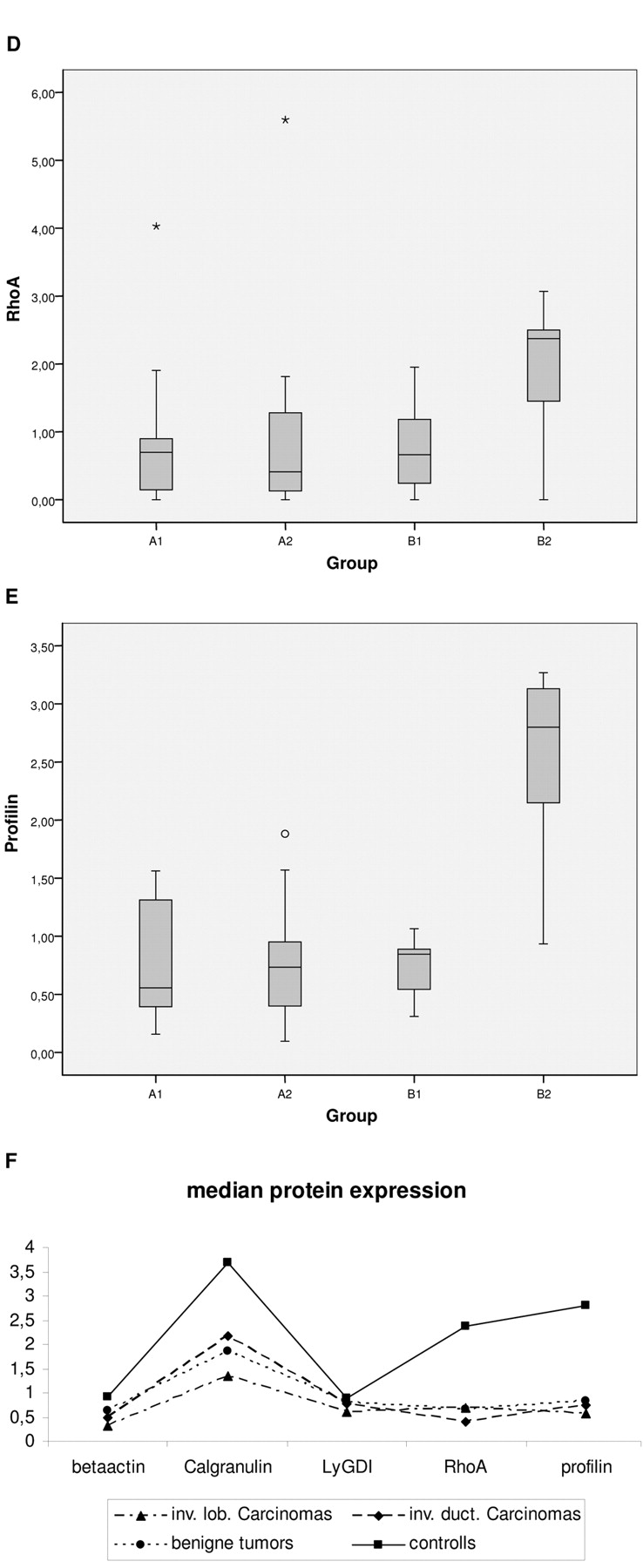
Median values as well as standard deviation of 25% - 75% calculated in sub-groups of comparison for A. actin, B. calgranulin, C. LyGDI, D. RhoA, and E. profiling; F demonstrates relations among corresponding medians in sub-groups.
Significance calculated for differential expression (median values) of proteins tested in (sub)groups of comparison. Classification of the (sub)groups is given in Table I.
Suppression of several key cytoskeletal proteins such as tropomyosin-1 has been hypothesized earlier to contribute to poor cytoskeleton architecture followed by neo-plastic transformation (14). As for cytoskeletal reorganization in malignant breast cancer cells, it can be triggered by different stress factors including mechanical forces (15). Consequent cytoskeletal remodeling and alteration of adhesion quality to the extra-cellular matrix components has been proposed to be directed by Rho GTPase and inhibitors (14, 16).
Less is known about potential alterations in the microfilamental network of circulating leukocytes in breast cancer patients. In the case of generalized similarities in molecular mechanisms, the characteristic patterns of microfilamental network-associated proteins identified in blood cells might be of great help for the development of non-invasive approaches in favor of early and predictive diagnosis of breast cancer. The protein mapping carried out in this work revealed a list of microfilamental network-associated proteins the expression levels of which are potentially altered in circulating leukocytes of breast cancer patients versus healthy controls. Concomitant protein quantifications were focused on the network-members demonstrated to be important for rearrangement of cytoskeletal architecture and motility in leukocytes (17).
The results clearly demonstrate significant alterations in the repertoire of microfilamental network-associated proteins in circulating leukocytes of breast cancer patients compared to healthy controls: the pathology-specific suppression of expression levels of Rho A, profilin, calgranulin, and even actin, is evident.
Rho A belongs to the family of small GTPases which are involved in the re/organization of the microfilamental network, cell-cell contact and carcinogenesis. Among malignancies, the most dramatic difference in the expression of Rho GTPases was observed in breast tissue (18). Breast malignancies demonstrate particularly high levels of RhoA expression which is the stable biomarker for tumor progression from WHO grade I to grade III. The suppression of Rho A expression levels in circulating leukocytes was not described for breast cancer patients till now, and the pathology-related meaning of this deregulation is currently unclear. However, dramatic consequences for the overall reorganization of microfilamental network in circulating leukocytes of breast cancer patients can be assumed, since Rho A is involved in the control of the [filamentous actin]: [monomeric actin] balance which is critical for many different functions of T-cells including lymphoid polarization, chemo-taxis and migration of tissue-infiltrating activated lymphocytes (19).
Profilin - the positive effector of actin polymerization - is concentrated at the leading edge of polarized, i.e. motile, T-cells. Translation of a chemoattractant signal into cell migration sparks the formation of actin-filament structures. Filopodia and lamellipodia formation at the leading edge of migrating cells requires accumulation of profilin in an area immediately posterior to the leading lamella; the extension of lamellipodium in the direction of cell movement is followed by adhesive activity located at the leading edge area and by forwarding of the cell body. Minimal changes in profilin concentration lead to dramatic alterations in the quality of fillopodia formation and, consequently, in cell migration potential (19, 20) - the alterations which can be expected in circulating leukocytes of breast cancer patients caused by the significant down-regulation of profilin demonstrated in this work. The cause for the altered profilin expression profile in circulating leukocytes of breast cancer patients is currently unclear. One plausible explanation might be an increased oxidative stress to T-cells that modify cytoskeletal proteins including profilin and actin as has been demonstrated by red-ox proteomics (21). Profilin-dependent alterations in circulating leukocytes of breast cancer patients have not been reported till now, although a role of profilin in breast cancer pathology is well documented in the literature. In invasive breast cancer cells compared to its non-tumorigenic counterparts, gene expression profiling revealed a dramatic down-regulation of profilin-isoforms (22, 23). Moreover, an over-expression of profilin-1 results in morphological and cytoskeletal reorganization as well as in suppression of tumorigenicity of breast cancer cells (22). Profilin-1 is classified as a tumor suppressor gene, low expression levels of which are critical for cell differentiation (24). Consequently, an over-expression of profilin-1 has been suggested as potentially successful strategy for controlling the metastatic potential of breast cancer cells (25).

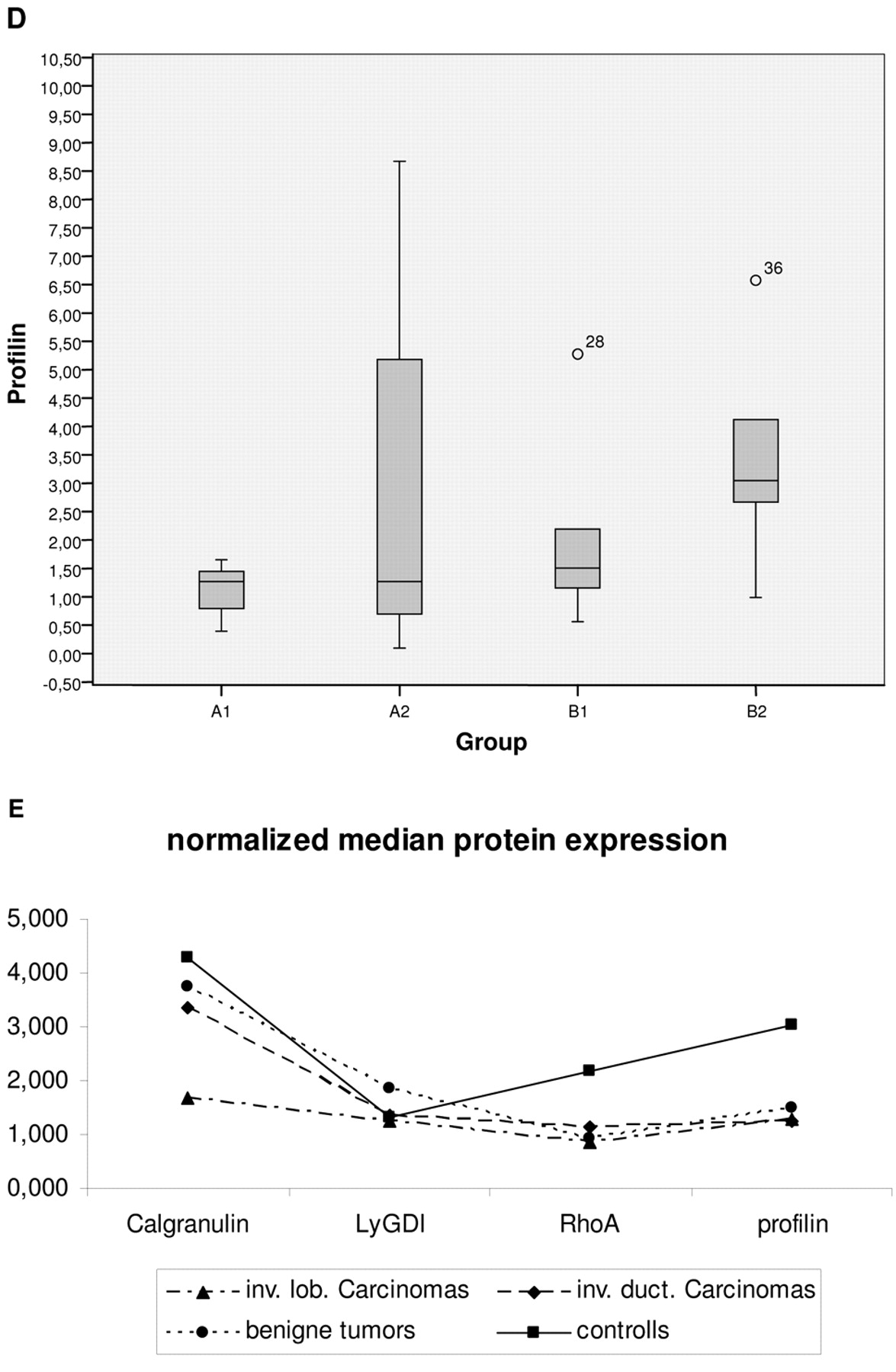
Expression levels of individual proteins tested were normalized by corresponding expression rates of actin - the house-keeping gene. Median values as well as standard deviation of 25% - 75% calculated in sub-groups of comparison are demonstrated for A. calgranulin, B. LyGDI, C. RhoA, and D. profiling; E shows relations among corresponding normalized medians in sub-groups.
Calgranulins are members of S100 family of Ca2+-binding proteins many of which are encoded by epidermal differentiation cluster on chromosome 1q21 - the area which is frequently rearranged in human tumors (26). Being potent chemo-attractants, S100 proteins modulate cytoskeletal interaction, cell growth, differentiation, contact, dissociation, migration activity as well as metastasis in cancer (27). Some of the family members demonstrate a tumor suppressor activity such as S100A2 gene which found to be silent in breast cancer due to hypermethylation of CpG-islands (28). Calgranulins A and B (S100A8 and S100A9, respectively) form a heterodimeric molecule - calprotectin - abundantly expressed in inflammatory and endothelial cells (27). S100A9 directly affects leukocyte tissue invasion and mediates cell contact dissociation via its calcium binding properties (29). Down-regulation of S100A8 in leukocytes of breast cancer patients was not reported till now, although perturbations in the expression profile of both calgranulin A and B were observed, particularly, in breast cancer tissues (26). S100A8 has distinct functions implicated in embryogenesis, growth, differentiation, immune and inflammatory processes (27). Inactivation of calgranulin A is embryonic lethal and a protective role against oxidative tissue damage has been proposed for the calgranulin A-homodimer disulfide-bonds which are highly sensitive towards oxidative stress.
Significance calculated for differential expression (median values) of proteins normalized by corresponding expression rates of actin as the house-keeping-gene. Classification of the (sub)groups of comparison is given in Table I. The best significances are given in bold figures.
Protein mapping carried out for circulating leukocytes of breast cancer patients revealed protein overlapping with those previously identified in breast cancer tissue and demonstrated pathology-specific perturbations in expressional levels including heat-shock proteins 60 and 70, cytokeratin, vinculin, enolase, alpha-1-antitrypsin, peroxiredoxin and actin (30). This overlap in altered expression profiles between malignant breast tissue and circulating leukocytes of the patients opens good perspectives for minimally-invasive diagnostic approaches based on the pathology-specific expression patterns of blood cells.
Comparative proteomics resulted in the selection of a relatively big group of microfilamental network-associated proteins which are functionally linked with each other. Perturbations in expression levels of these proteins result in dramatic cytoskeletal reorganization, altered cell migration potential and have been implicated specifically in breast cancer pathology. For the first time, these alterations are reported here for circulating leukocytes of breast cancer patients. One of the plausible explanations for the detected alterations might be an increased oxidative stress to blood cells, since all proteins tested are highly sensitive towards and regulated/modified by stress conditions.
Correlation between normalized protein expression levels in pairs of comparison.
Pathology-specific expression patterns of RhoA, profilin-1 and calgranulin A identified in this work should be further taken into consideration to create a novel highly sensitive minimally-invasive approach for predictive diagnosis of breast cancer.
Acknowledgements
The authors thank Mrs. H. Mönkemann, PhD for blood collection and Mrs. G. Windisch-Schuster for her excellent technical assistance in the performance of “Western-blot” analysis.
Footnotes
- Received November 4, 2008.
- Revision received December 16, 2008.
- Accepted December 19, 2008.
- Copyright© 2009 International Institute of Anticaner Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.