


Abstract
Formation of metastases from various tumor entities in the brain is a major problem for the treatment of advanced cancer. We describe target molecules and tools for the delivery of small molecules or proteins across the blood–brain barrier (BBB), and the treatment of brain tumors and metastases with antibody-related moieties. In addition, drugs preventing formation of metastases or interfering with the growth of established metastases are described, as well as pre-clinical metastasis models and corresponding clinical data. Furthermore, we discuss the delivery of effector proteins and antibody-based moieties fused with an antibody-based scaffold across the BBB in several model systems which might be applicable for the treatment of brain metastases.
- Antibody-effector protein fusion
- antibody–antibody fusion
- tropism of metastasis
- transferrin receptor
- insulin receptor
- animal metastasis models
- receptor-mediated endocytosis
- molecular Trojan horses
- review
Although some primary brain tumors such as glioblastoma are associated with a dismal prognosis (1), metastatic lesions originating from various primary tumors outside the brain are in fact responsible for 90% of cancer deaths from brain malignancies (2). Thus, primary brain tumors such as glioblastoma mainly act through local invasive growth and rarely metastasize to organs outside of the brain, whereas many peripheral tumors, such as pancreatic carcinoma or small cell lung carcinoma (SCLC), have usually metastasized to distant organs before diagnosis (1, 3). Moreover, while metastases derived from certain types of tumors such as SCLC colonize various distant organs, others exhibit a pronounced organ-specific tropism of metastasis (3-7). Prostate cancer preferentially metastasizes to the bones, colon cancer to the liver, and sarcoma to the lungs (4). Importantly, distant brain metastases most commonly arise from primary tumors of the lung (50-60%), breast (15-20%), melanoma (5-10%) and the gastrointestinal tract (4-6%) (8, 9). The incidence of such brain metastasis is estimated to be approximately 170,000/year in the USA, 10-times the number of primary brain tumors (10). Accordingly, successful treatment of primary brain tumors and brain metastases poses a major medical problem which almost certainly will be significantly affected once efficient and reliable delivery of drugs into the brain become available.
The Blood–Brain Barrier (BBB)
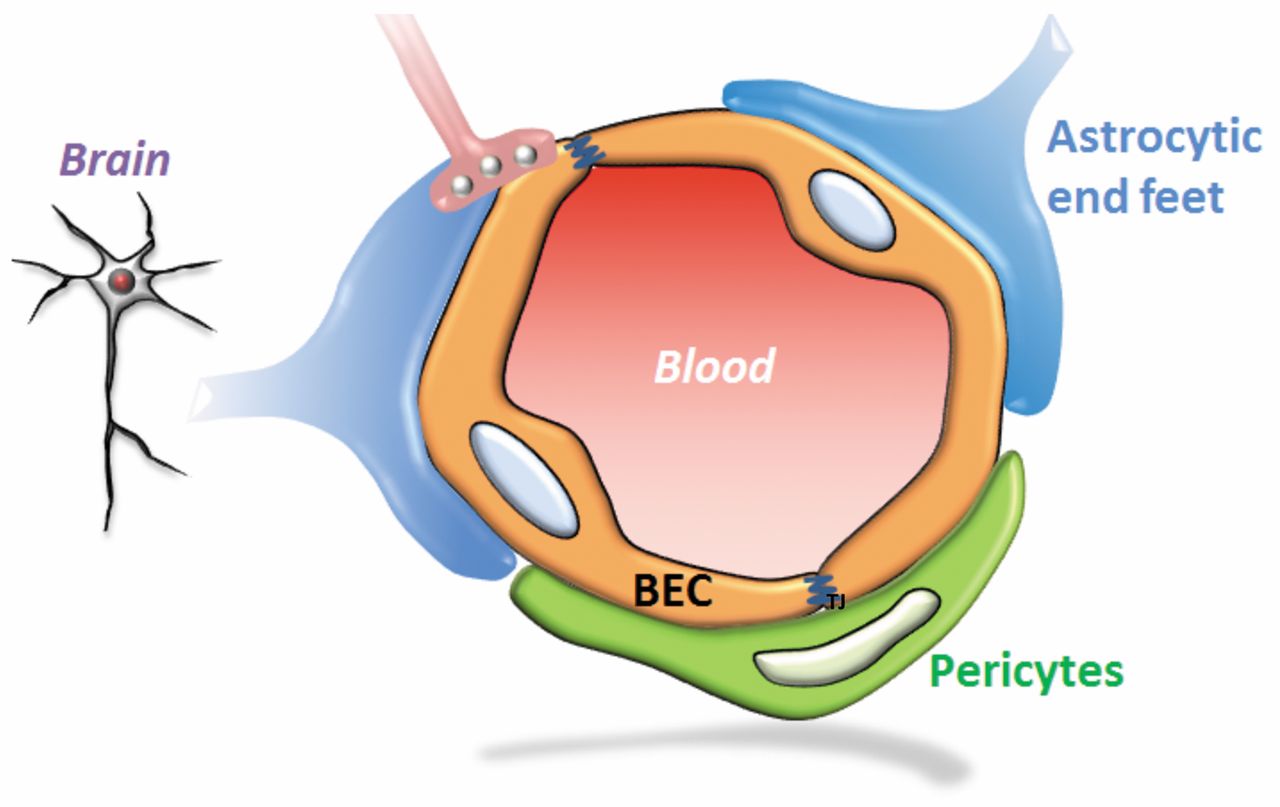
The BBB is a highly selective barrier regulating the uncontrolled diffusion of most molecules into the brain. Thus, tissue homeostasis is maintained by enabling the transport of selected substances necessary for the brain, while access to the brain is blocked for most other molecules, most notably for toxic metabolites and xenobiotics (11). Thus, for a small molecule, free diffusion across the BBB requires both lipophilicity and a molecular mass smaller than 400-500 Da (12, 13). Most large molecules such as immunoglobulins, however, are not allowed to cross the BBB unless transported actively into the brain. On the other hand, due to the presence of neonatal fragment crystallizable region (Fc) receptors (FcRn) and other clearance mechanisms, IgG antibodies are actively transported from the brain into the circulation (14). In patients with brain tumors, leakiness of the BBB has been observed (15). The complete surface area of the BBB comprises approximately 20 m2, with a total length of capillaries in the brain of approximately 600,000 m (11). The pivotal component of the BBB (Figure 1) is a monolayer of endothelial cells in brain capillaries which are connected by tight junctions (TJ). Absence of fenestration also contributes to the barrier properties of brain endothelial cells. In addition, pericytes, astrocytes and neurons are involved in controlling permeability and other functions of the BBB (11-13, 16-17). The TJ between endothelial cells are composed of proteins such as occludins, claudins and junctional adhesion molecules. The endothelial cells are also covered by a specialized extracellular matrix and the basement membrane which is composed of collagen (particularly type IV), fibronectin, laminin, tenascin and proteoglycans.
Three classes of transport systems have been identified in the BBB: receptor mediated transport (RMT), carrier-mediated transport (CMT), and active efflux transport (AET) (11). RMT is based on endocytosis at the luminal (blood) side, intracellular transport, and exocytosis at the abluminal (brain) side of the endothelium. RMT allows large molecules such as proteins to be transported, thus being potentially useful for the specific delivery of anti-tumor protein therapeutics into the brain. Examples for receptors involved in RMT are transferrin receptor (TfR), insulin receptor (IR), leptin receptor, and lipoprotein-related protein-1 and -2 receptors (18, 19). CMT is mediated by members of the solute carrier (SLC) receptor family (20). This system is involved in the transport of specific substrates such as amino acids, sugars, organic anions and cations, metabolites, nutrients and neurotransmitters. Finally, AET is an ATP-dependent mechanism responsible for the transport of compounds from the brain into the circulation and is mediated by multidrug resistance protein1 (ABCB1) (21), breast cancer resistance protein (ABCG2) (22) and the multidrug resistance-associated protein (ABCC1) (23) family of proteins. For example, ABCB1 is involved in the resistance of central nervous system (CNS) tumors to treatment with paclitaxel and anthracyclines (21). ABCG2 substrates are mitoxantrone, topotecan, methotrexate and sulfated conjugates (22). ABCC1 family members are efflux pumps and capable of transporting structurally diverse lipophilic drugs such as anthracyclins, vinca alkaloids, methotrexate and camptothecins (23).
General Remarks on Drug Delivery Across the BBB
In the following, we focus on the delivery of drugs across the BBB; we do not discuss direct delivery of drugs into the CNS such as intracerebral administration (intrathecal or intraventricular), or convection-enhanced distribution. We cover delivery of small-molecule and protein therapeutics both via targeted and non-targeted approaches. With regard to targeted approaches, we focus on receptor-mediated endocytosis of protein therapeutics. Delivery of other moieties such as liposomes, dendrimers, quantum dots, nanoparticles and micelles using internalizing antibodies (24) are not the focus of this review. Similarly, exosome-mediated delivery (25), localized exposure to high-density focused ultrasound, use of vasoactive compounds such as bradykinin, blockage of active efflux transporters, modulation of tight junctions by enterotoxins (26) and cationisation of drugs (27) are not discussed.
Animal Models of Brain Metastasis
In animal models of brain metastasis, tumor cells were shown to leave the blood vessels, migrate along those vessels, and interact with the inflamed microenvironment of the brain, thus establishing micrometastases. In addition, communication with activated microglia and astrocytes was observed. Once established tumors have formed, angiogenesis can be initiated by vessel cooption, but also dormant solitary tumors were observed to reside within the brain (28-30). Accordingly, numerous animal models for brain metastasis have been established based on syngeneic rodent models, human rodent xenograft models, both ectopic and orthotopic, and orthotopic genetically modified mouse models. In some cases, variants of tumor cell lines prone to form metastases were established by repeated in vivo passages and selection for brain tropism (31). In addition, intracardiac, intracarotid, or orthotopic inoculation are possible means of establishing experimental brain metastases (31-36). For example, triple-negative breast cancer cell lines such as MDA-MB-231, 4T1 and CN34 were selected for brain tropism of metastasis (37-39). Similarly, human epidermal growth factor receptor-2 (HER2)-transfected 231-BR cells, a metastatic cell line derived from MDA-MB-231, have a 2.5- to 3-fold greater potential to form brain metastases than the non-transfected parental cell line (40, 41).
Three types of microenvironment allowing the generation of brain metastases have been identified: the perivascular niche, the brain parenchyma, and the cerebrospinal fluid (CSF) or leptomeningeal niche, which is the microenvironment containing the CSF and the lining of the brain (42). Using transcriptional profiling of CN34 and MDA-MB-231 breast cancer cells, cyclo-oxygenase 2, epidermal growth factor receptor (EGFR) ligand, heparin-binding epidermal growth factor and α-2,6 sialyltransferase (STGALNAC5) were identified and validated as mediators for cancer cell passage through the BBB (43). The functional role of STGALNAC5 in brain metastasis was verified using BBB trans-migration assays and short-hairpin RNAs (37). Importantly, STGALNAC5 underscores the role of protein and, as a consequence, cell surface glycosylation in organ-specific metastatic interactions through cooption of an enzyme which is normally expressed in the brain. EGFR ligands and cyclo-oxygenase 2 were previously linked to breast cancer infiltration of the lungs, but not the bones or the liver (44, 45). RNA analysis of the metastasis-forming cell line 231-BR indicated down-regulation of several metastasis-suppressor genes and up-regulation of hexokinase 2, and correlation of hexokinase 2 up-regulation with poor prognosis and its possible value as a therapeutic target (45).
Other pre-clinical models were used to identify additional signaling pathways and components important for brain metastasis formation from primary breast cancer such as up-regulation of cysteine X cysteine (CXC) chemokine receptor 4/CXC chemokine ligand 12 (CXCL 12), vascular endothelial growth factor (VEGF), Notch, and phosphatidyl-inositol-3-kinase (46-48). Similarly, overexpression of transforming growth factor-β2 in B16 melanoma cells induced formation of metastases in brain parenchyma, while no impact on leptomingeal metastasis formation was observed (49). Transfection of A375 melanoma cells possessing a brain tropism due to a signal transducer and activator of transcription 3 expression vector also increased the incidence of brain metastases, which was linked to decreased expression of suppressor of cytokine signaling (50).

Schematic representation of the blood–brain barrier. The interaction of brain endothelial cells (BEC) with astrocytes and pericytes is shown.
Many therapeutic compounds have been evaluated in models of metastasis with brain tropism. End-points were usually either the prevention of the development of metastases, or shrinkage or stabilization of already established metastases (39). The 231-BR breast cancer model was used to identify critical factors regarding the treatment of brain metastases (38). For example, variable penetration of paclitaxel into 231-BR-derived brain metastases was observed. Unrestricted uptake of radiolabeled drug was seen in 10% of the lesions (≥50-fold higher than in normal brain), 14% of the lesions showed drug uptake as in normal brain tissue, and the remainder of the lesions fell in between these extremes. Not surprisingly, tumor cell death was observed only in 10% of metastases with the highest paclitaxel penetration (38). Similar observations were made for lapatinib. Here, 17% of the lesions were freely accessible while the others behaved like normal brain (38). Thus, mice bearing 231-BR tumors which were injected with lapatinib 5 days after tumor inoculation showed a 54% reduction of large metastases 24 days after the start of treatment which corresponded with reduced HER2 phosphorylation (41). Similarly, again using the 231-BR model, vorinostat (suberoylanilide hydroxamic acid), a BBB-permeating histone-deacetylase inhibitor, prevented formation of micrometastases (28%) and large metastases (62%) when the administration was started 3 days after tumor cell inoculation (51, 52). In some pre-clinical models, VEGF was shown to promote the formation of brain metastases, and treatment with anti-angiogenic agents led to reduction in metastatic outgrowth (28, 53). In additional pre-clinical models, inhibition of VEGF receptor 2 allowed for a critical period of blood flow normalization which optimally potentiated the therapeutic effect (54, 55). However, while treatment of Mel 57 melanoma-derived brain metastases with the VEGF receptor inhibitor ZD 6474 resulted in initial growth inhibition, this was followed by the formation multiple proangiogenic lesions which sustained tumor progression (56). Finally, using an orthotopic BT474 brain metastasis model, the combination of monoclonal antibodies against HER2 and VEGF resulted in reduced microvascular density and increased necrosis in brain lesions, and improved survival in comparison to treatment with the single agents (57).
Treatment of Brain Tumors and Brain Metastases with Small Molecules in the Clinic
The mainstay of treatment of primary brain tumors and brain metastases is surgery and radiation therapy, albeit with limited therapeutic benefit (58, 59). In addition, cytotoxic/ cytostatic agents with good CNS penetration are temozolomide, irinotecan, topotecan, procarbazine and carboplatin. Out of these, temozolomide is an approved agent for the treatment of astrocytoma and glioblastoma, whereas the clinical efficacy of other agents for treatment of brain tumors and metastases is still under investigation (59). In order to facilitate BBB passage, ANG 1005, a novel derivative of paclitaxel, was conjugated to a peptide vector (Angiopep-2) and is currently being evaluated in clinical trials (60). Interestingly, it was shown that non-SCLC-derived brain metastases express increased levels of EGF and phosphorylated EGFR compared to primary tumors from these patients (61). Thus, erlotinib, gefitinib and afatinib, which are EGFR inhibitors for the treatment of non-SCLC with activating EGFR mutations, are being evaluated in nine clinical trials for treatment of non-SCLC-derived brain metastases, with gefitinib and lapatinib as single agents, or in combination with temozolomide or whole-body irradiation. Here, there are promising hints of efficacy, which are yet to be confirmed in randomized clinical studies (61). The echinoderm microtubule-associated-protein-like 4/anaplastic lymphoma kinase (EML4–ALK) translocation which leads to constitutive activation of ALK has been identified in 3-5% of non-SCLC. There is evidence that the ALK-inhibitor crizotinib inhibits brain metastases. However, crizotinib is a substrate for drug efflux pumps and newer compounds might have a better efficacy profile (62-64).
Brain metastases are frequent in patients with malignant melanoma and are a common cause of death (65). Here, hyperactivation of rapidly accelerated fibrosarcoma (RAF) signaling is often observed (66). In clinical studies with a small number of patients, B-RAF inhibitors such as dabrafenib induced shrinkage of brain metastases or led to complete responses (67, 68). Unfortunately, however, these promising phase I results could not be reproduced in a prospective open-labeled phase II study in patients with untreated progressive melanoma-derived brain metastases. Overall, disease control in patients with B-RAF inhibitors was 72%, but complete responses were rare (69). It will be interesting to see whether ipilimumab can overcome immune tolerance of brain metastases after immune activation by cytotoxic T-lymphocyte antigen-4 blockade (70).
CNS metastases are quite common in patients with HER2-positive or triple-negative breast cancer. In fact, 30-55% of patients with HER2-positive breast cancer will develop brain metastases (71-74). Small-molecule HER2 inhibitors, such as papazanib, lapatinib, neratinib, afatinib and ARRY-380, are under investigation in patients with progressive CNS metastasis (67). Other agents under clinical evaluation in the control of breast cancer metastases are poly ADP-ribose polymerase inhibitors (such as olaparib, iniparib and veliparib), vorinostat, mammalian target of rapamycin inhibitor everolimus and phosphatidyl-inositol-3-kinase inhibitor BK 120 (67). Finally, brain metastases were visualized with 89Zr-labeled trastuzumab, indicating that at least to some extent this antibody can pass the BBB (75).
Treatment of Brain Tumors and Brain Metastases with Antibody-derived Moieties
Disruption of the BBB and change of composition of the extracellular matrix by CNS tumors is well-demonstrated (13). Accordingly, in patients with brain tumors, the BBB is leaky at the tumor center. The endothelium of blood vessels feeding the proliferating edge of the tumor and the surrounding brain tissue, however, is not compromised in its function and is as impermeable as an intact BBB (76). Thus, only 0.1-0.2% of administered therapeutic antibodies cross the BBB, enter the CSF, and are found in the brain at steady-state concentrations (77-79). Penetration of the BBB by antibodies can also be facilitated by cell-mediated immune responses leading e.g. to paraneoplastic neurological disorders in a fraction of patients with SCLC-derived brain metastases based on an immune response against autoantigen HuD protein shared by SCLC and neurons (80). In this respect, approval of bevacizumab for treatment of recurrent glioblastoma is a milestone for antibody-based treatment of brain tumors (13). In addition, bevacizumab therapy is currently being evaluated for the treatment of newly diagnosed glioblastoma, CNS lymphoma and secondary cerebral metastases derived from glioblastoma (13). However, regarding its mode of action, one should be aware of the fact that bevacizumab probably does not have to cross the BBB to function because of its ability to neutralize VEGF in the lumen of the capillaries of blood vessels in the brain (13). In general, however, the problem of limited CNS accessibility for antibody-derived therapeutics has to be solved in order to allow a more widespread use against brain tumors and metastases. These issues are presently being addressed in numerous pre-clinical and clinical investigations, targeting molecules such as EGFR, EGFR variant III (EGFRvIII), HER2 and tenascin C (81-86). In that respect, recent clinical studies have shown that brain metastases may be responsive to immunomodulatory treatment. Thus, ipilimumab, a cytotoxic T-lymphocyte antigen-4-based immune-checkpoint modulator, was evaluated in a clinical trial that enrolled both steroid-naïve and steroid-treated patients with melanoma and brain metastases (87). Partial responses were observed in patients of both cohorts. Occasional follow-up responses at distant sites were observed after treatment with ipilimumab and radiation. This phenomenon is referred to as the abscopal effect and a possible explanation is the unmasking of tumor antigens by radiation at local and distant sites, and the subsequent induction of an immune response (88, 89).
Receptor-mediated Drug Delivery Across the BBB
In order to improve the efficacy of delivery of potential protein-based therapeutic molecules across the BBB into brain tumors and metastases, several target molecules expressed on endothelial cells of the BBB and corresponding antibodies as delivery vehicles were examined. Such brain-delivery is based on receptor-mediated transcytosis, and the underlying principle is referred to as a molecular Trojan horse (MTH) strategy (90, 91). For example, the TfR is highly expressed in endothelial cells of the BBB (92), and is a recycling receptor which is internalized into clathrin-coated pits, independent of transferrin (92). TfR cycling is rapid and occurs within minutes and shuttles cargos from the lumen of the blood vessels into the brain parenchyma. Proof-of-concept experiments for brain delivery of TfR-specific antibodies were conducted with antibody OX26, a mouse mAb directed against rat TfR (93-95). The OX26 binding site on TfR is different from the Tf-binding site and therefore does not interfere with iron transport. Similarly, mouse TfR-specific antibodies 8D3 and R17-217 were isolated (96, 97). 8D3 conjugated to radiolabeled Aβ was used to visualize and quantify β-amyloid plaques in a mouse model of Alzheimer's disease (97). In an in vitro test using human brain capillaries, the most potent MTH was an antibody directed against the human IR (98). Similarly, in in vivo studies in mice, a genetically-engineered mouse–rat chimeric monoclonal antibody against the mouse TfR referred to as cTfRmAb was used successfully (99). In addition, ligands of low-density receptor related protein-1 and -2, receptor-related protein, apolipoprotein E and melanotransferrin/p97 were evaluated for delivery of paclitaxel or adriamycin in preclinical models (100-103). However, brain-specific drug delivery via lipoprotein-related proteins has not been shown yet and probably would require their higher expression on endothelial cells. Finally, the 15-kDa single domain antibodies FC5 and FC44 (104), derived from camelid phage libraries, enable receptor-mediated endocytosis of antibody–antigen complexes at the luminal side of brain endothelial cells (105, 106). The target of FC5 was identified as cell-division cycle 50A (106). However, the short plasma half-life of 10 minutes of these molecules is a critical issue.

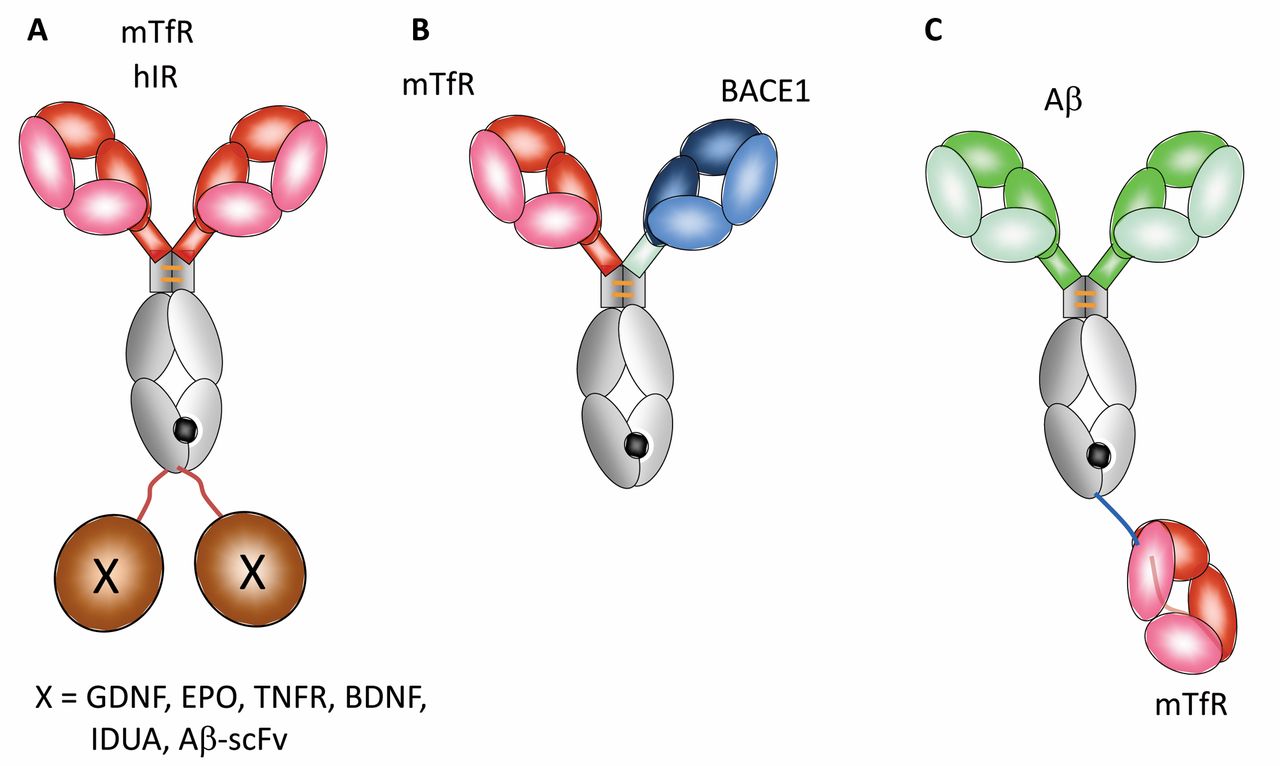
Antibody-based moieties for delivery to the brain. A: Monoclonal antibodies (mAb) directed against the transferrin receptor (TfR) or human insulin receptor (hIR) fused to protein-based payloads. Possible payloads: Glial cell line-derived neurotrophic factor (GDNF), erythropoietin (EPO), brain-derived neurotrophic factor (BDNF), α-L iduronidase (IDUA) and single-chain Fv anti-Aβ mAb (scFv-Aβ). B: Bispecific antibody directed against TfR and β-site amyloid precursor protein cleaving enzyme (BACE1). C: Antibody directed against Aβ fused with a Fab fragment directed against the TfR. Protein payloads are shown in brown, antibody constant regions in grey, anti-TfR or anti-hIR variable regions or Fab fragments, anti-BACE1 variable regions and anti-Aβ variable regions are shown in pink, blue and green, respectively.

Cellular routing of antibody–antibody fusion proteins in brain endothelial cells. A: Antibody directed against amyloid Aβ fused to two Fab-fragments (dFabs) against the transferrin receptor (TfR). B: Antibody directed against amyloid Aβ fused to a single Fab (sFab) fragment against TfR. The white circles represent endosomes, TfR is shown by blue pentagons, V regions of the anti-Aβ monoclonal antibody are shown in green, constant antibody regions in grey and TfR-Fab moieties are shown in red.
Antibody–Effector Protein Fusions as Therapeutic Agents in Pre-clinical Models
Recombinant fusion proteins can be engineered for brain penetration by fusing an antibody directed against an endogenous BBB receptor such as IR or TfR to a therapeutic effector protein (97) (Figure 2A). Here, the targeting antibody acts as an MTH to ferry the effector moiety into the brain. An earlier version of this technology was based on affinity-mediated coupling of antibodies and effector proteins via the biotin-avidin technology (97). These approaches are also to be preferred, because general BBB disruption leads to chronic neuropathological changes and therefore cannot widely be used in humans (107, 108). In the newer versions of delivery molecules, the respective effector moiety can be fused either to the N- or to the C-terminus of the heavy chain of the BBB-targeting antibody and usually does not interfere with its target binding or other biological functions. For example, 2-3% of brain uptake at steady-state levels of fusion proteins between cTfRmAb and glial cell-derived neurotrophic factor, tumor necrosis factor receptor and erythropoietin was observed (109-112). Similarly, antibody fusion molecules with brain-derived neurotrophic factor (112) or potentially therapeutic enzymes such as β-glucuronidase (113) and iduronate-2-sulfatase (114) were evaluated.
Pharmacological effects on the CNS after i.v. administration of fusion proteins as outlined above were observed in mouse and rat for erythropoietin in stroke, for glial cell-derived neurotrophic factor in Parkinson's disease (115), and α-L-iduronidase in Hurler's syndrome. However, as a rule, fusion proteins based on TfR-or IR-specific antibodies are rapidly cleared from the circulation due to expression of these receptors on peripheral tissues such as kidney and liver (93).
Transfer of Antibody–Antibody Fusion Molecules Across the BBB in Pre-clinical Models
Therapeutic antibodies can be ferried across the BBB when fused to a binding moiety specific for an endocytosing BBB antigen. Such molecules were evaluated either as bispecific antibodies or as fusion proteins consisting of scFv or Fab effector moieties connected to a BBB-penetrating antibody (Figure 2B and C). The following examples are derived from studies on neurological disorders but probably can be extrapolated to treatment of brain cancer or secondary brain metastases. Proof-of-concept experiments were performed using a scFv Aβ-specific antibody fragment fused to a HIR-specific monoclonal antibody or to a cTfRmAb (116, 117). The anti-Aβ scFv was fused to both antibody CH3 domains, bound to Aβ amyloid, and caused disaggregation of the amyloid in the brain. Similarly, an antibody directed both against TfR and β-site amyloid precursor protein cleaving enzyme (BACE1) was evaluated in a bispecific format (118, 119). BACE1 is an aspartyl-protease involved in the production of Aβ in the brain and in the periphery, and inhibition of β-amyloid production in vivo was demonstrated using such fusion molecules. Using therapeutic doses (5-50 mg/kg), it was shown that low-affinity anti-TfR antibodies were transported more rapidly and efficiently into the brain, improving duration of exposure and brain uptake in comparison to high-affinity antibodies (117, 118). However, side-effects were observed: a single dose of 1 mg/kg caused severe acute clinical signs, as well as a profound reduction of reticulocyte counts in mice (118, 119). It was shown that the elimination of Ig-Fc-mediated effector functions prevented these acute clinical signs and mitigated the reduction of reticulocyte count (118, 119). These findings support efforts to identify target molecules more specific for the BBB.
An approach based on a TfRmAb–Fab fusion protein constructed in a modular manner was recently investigated (120, 121). One or two Fab fragments of an anti-TfR-specific antibody (122) were fused to an anti-Aβ monoclonal antibody. Both molecules were taken up by endothelial cells by transcytosis; however, whereas the monovalent construct mediated uptake, transcytosis and TfR recycling, the presence of two TfR-binding Fab fragments per anti-Aβ resulted in uptake and trafficking to lysosomes followed by reduction of TfR levels (Figure 3). In a transgenic mouse model of Alzheimer's disease, the antibody bearing one TfR-binding unit exhibited a 55-fold increase in amyloid plaque engagement compared to the parental antibody or the antibody with two TfR-binding moieties. Localization of the antibody with the monovalent TfR binder in the brain was shown by immunofluorescence-based signal amplification, however, information regarding quantitation in the brain and toxicity data are not yet available. Extension of these studies as outlined above to brain oncology might lead to therapeutically-promising approaches.
Concluding Remarks
Future progress in the treatment of brain tumors and brain metastases will depend on several issues. Importantly, from a technical point of view, optimization of delivery of small molecules and biological agents is a prerequisite for increasing treatment efficacy. In addition, inhibition of spread of established metastases might confer therapeutic benefit. Hopefully, stepwise increments of therapeutic benefit can be demonstrated in the years ahead.
- Received May 20, 2015.
- Revision received June 17, 2015.
- Accepted June 18, 2015.
- Copyright© 2015, International Institute of Anticancer Research (Dr. John G. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- The Blood–Brain Barrier (BBB)
- General Remarks on Drug Delivery Across the BBB
- Animal Models of Brain Metastasis
- Treatment of Brain Tumors and Brain Metastases with Small Molecules in the Clinic
- Treatment of Brain Tumors and Brain Metastases with Antibody-derived Moieties
- Receptor-mediated Drug Delivery Across the BBB
- Antibody–Effector Protein Fusions as Therapeutic Agents in Pre-clinical Models
- Transfer of Antibody–Antibody Fusion Molecules Across the BBB in Pre-clinical Models
- Concluding Remarks
- References
- Figures & Data
- Info & Metrics
Related Articles
Cited By...
- Improved Drug Delivery to Brain Metastases by Peptide-Mediated Permeabilization of the Blood-Brain Barrier
- Emerging Gene Fusion Drivers in Primary and Metastatic Central Nervous System Malignancies: A Review of Available Evidence for Systemic Targeted Therapies
- Impact of Blood-Brain Barrier Integrity on Tumor Growth and Therapy Response in Brain Metastases
- Dissection of the Process of Brain Metastasis Reveals Targets and Mechanisms for Molecular-based Intervention