


Abstract
Background: The main purified compound from soy protein isolates is genistein, but this purified phytoestrogen fails to recapitulate all the features of the soy-based diet that is associated with lower incidence of prostate cancer. Materials and Methods: Rat and human prostate cancer cell lines were treated with either soy protein isolates or purified genistein. In vitro cell growth was correlated with the associated genomic expression profiles using cDNA arrays. The data was subsequently bioinformatically analyzed within and across species to identify common changes in expression profiles associated with the soy protein or genistein treatments. Results: Gene expression profiling and data mining noted genes specific to soy; however, biological pathways showed the same gene regulation profiles between genistein and soy. Conclusion: Genistein is likely the major contributor to the effect of soy proteins on cellular pathways; however, the expression of different genes using soy protein isolates suggests complexity in the many compounds found in whole soy protein.
Prostate cancer is the most common solid tumor and the second leading cause of death among American men (1). Additionally, the risk of prostate cancer is 1 in 6 and the risk of death due to metastatic prostate cancer is 1 in 30 (2). Prostate cancer is a heterogeneous disease ranging from asymptomatic to a rapidly fatal systemic malignancy (3). Aggressive prostate cancer is significantly less prevalent among Asian men and more in African American men (4). Another common description of prostate cancer is that of hereditary versus sporadic cancers (3). This division occurs at the epidemiologic level but fails to distinguish their behavior or molecular activity. While some genes have been identified that confer a reasonable level of risk for prostate cancer in families, most prostate cancers appear to be sporadic. The sporadic tumors most likely arise from the interactions of multiple genes and environmental effects that in combination lead to increased prostate cancer risk and tumor formation (5). Factors such as diet are thought to cause a fair majority of the genetic alterations leading to cancer.
The incidence and mortality of prostate cancer vary widely in different regions of the world. Some research points to high rates of prostate cancer attributed to food products, most commonly animal fats. On the contrary, decreased incidences were found in those populations with increased intake of fruits, vegetables, and vitamins (5). A large number of different chemicals in foods has been implicated in cancer prevention. Particularly in regard to prostate cancer, the administration of anti-oxidants (e.g. selenium, vitamin E, lycopene) and the consumption of foods rich in carcinogen-detoxification enzyme inducers (e.g. vegetables containing sulforaphane) have been the focus of recent studies and clinical trials (5). Epidemiological studies show that the risk for death from prostate cancer in the United States is elevated compared to other countries, especially Asian countries. It has been suggested that this result may be rooted in the environmental affect on diet rather than genetic profile (6). This is based on the increased incidence of prostate cancer in Asian men that have altered their food consumption and become accustomed to an American diet.
Soy protein in particular is consumed in much higher quantities in Asian cultures than in regular American diets. High consumption of foods containing soy results in high plasma, urine, and prostatic fluid concentrations of phytoestrogens, including genistein, and may play a role in the lower incidence in hormone-dependent cancer in Asian countries (4-8). Phytoestrogens are plant compounds found to have weak estrogenic affect on cells by showing a chemical structure similar to 17β-estradiol. Lignands, flavonoids, and isoflavonoids make up the different categories of phytoestrogens. The isoflavonoids that are highly studied include genistein, daidzein, formonentin, and biochanin A. However, genistein and daidzein are thought to be the most important isoflavones in prostate cancer. Many different foods carry varying concentrations of these phytoestrogens, but soy holds the highest concentration. It is this source of protein that is consumed in high quantities by Asian peoples in their native countries and may play a part in the lower incidence of hormone-dependent cancer in those countries (5).
Previously, Dr. Morris Pollard had performed studies showing decreased cancer development in Lobound-Wistar rats that consumed a soy diet compared to controls (10-12). Other animal studies, such as those with the TRAMP mouse model, have shown dose-dependent reduction in the incidence of prostate cancer in relation to increasing levels of genistein (4). Similar affects have been shown in treating tumors of LNCaP androgen-sensitive prostate cancer cell line in SCID mice fed low-fat and high soy diets (13). Thus using various animal models, soy continues to recapitulate the effect seen in the earlier epidemiological studies showing the decreased incidence of hormone-dependent prostate cancer in association with high levels of soy consumption, showing probable cause for human studies. Though multiple different rat models are promising, human studies have shown mixed results.
This current research seeks to elucidate the underlying molecular basis of the effects of soy protein on prostate cancer, while providing potential biomarkers for future evaluation of clinical trials using soy proteins. The studies include solublizing soy protein isolates into cell culture medium, applying this medium to human and rat prostate cancer cell lines, and observing changes in cell proliferation through growth curves. In addition, RNA was isolated and examined using cDNA microarray studies to identify changes in gene transcription. This information will provide a basis for developing a molecular explanation of soy protein anticancer affect.
Materials and Methods
Cell culture and soy or genistein treatment. The study used human and rat prostate cancer cells lines PC3 (human) and PAIII (rat). The PAIII cell line was selected due to its origin and commonality to the Lobund-Wistar rat prostate cancer (metastatic origin). Soy protein isolates have been obtained from Dr. Morris Pollard (Notre Dame, IN, USA), and are from the identical lot used in the associated Lobund-Wistar rat studies (10-12). The material was originally obtained from Harlan TekLad Diets in Madison, Wisconsin. Genistein was obtained from Sigma (St. Louis, MO, USA) (MDL Number: MFCD00016952). Cell lines were received from ATCC, Rockford, MD, USA (LNCap), from Drs. Paul Lindholm and Andre Kadjacsy-Balla (MCW, Milwaukee, WI, USA) (PC3), or Dr. Morris Pollard and Mark Suckow (Notre Dame, South Bend, IN, USA) (PA-III). These cells were cultured in RPMI supplemented with 10% fetal calf serum, 10 mM glutamine, and 10mM sodium pyruvate, and passaged 1:8 or 1:10 when the cells reached 70-80% confluence with trypsin-EDTA passaged before use in the experimental protocols. Soy protein was solubulized in a 0.005% NaOH and water solution. After solublization and sterile filtering, the soy protein was added to the medium of the cancer cells. These cells were then treated with various log concentrations (from 10 to 1000 mg/mL) of the soy protein isolates and their growth rates were measured by growth curves on PAIII, PC3 and LNCaP cell lines (Figure 1a, 2a, and 3a, respectively). From these growth curves using log concentration, the concentration of 100 mg/mL was selected for this study. These preliminary growth curve studies were also performed with genistein, a well-studied soy protein isolate. All growth curves were performed in triplicate and the average number of cells is displayed for each data point. Concentrations of 20 mM and 100 mM genistein were used for growth curve studies in both cell lines (Figure 1b, 2b, 3b, respectively).
RNA isolation and quantization. RNA was isolated from cells using Trizol (cat #15596018; Invitrogen, Carlsbad, CA, USA) and subsequently examined for quality using agarose gel electrophoresis and Gelstar nucleic acid stain against known RNA standards and failed to demonstrate significant degradation based on the presence of high molecular weight RNA species, and intact 28S and 18S ribosomal RNA bands. RNA, DNA and protein were isolated from the cells treated with either 100 mg/mL soy protein isolates or 20 mM genistein grown in 80% confluence over 5 days.
DNA library preparation and amplification. Libraries were prepared as previous described (14, 15). Sequence-verified rat and human libraries (Research Genetics, Huntsville, AL, USA, and University of Iowa cDNA clone set, IA), consisting of 41,472 human clones and 36,000 rat clones were used as a source of probe DNA. Subsets of 200 randomly selected clones had previously been chosen from these libraries, resequenced locally and demonstrated clone accuracy of 92%. We have opted to reformat libraries from 96 to 384-format for culture growth/archiving, PCR, purification, and printing. This has reduced the number of plates of our 41,472 human clone library from 432 to a more manageable 108, and the rat clone library from 375 to 94. The library was reformatted and subsequently manipulated using slot pin replicator tools (VP Scientific, San Diego, CA, USA). Cultures were grown in 150 μl Terrific Broth (Sigma, St. Louis, MO, USA) supplemented with 100 mg/ml ampicillin in 384 deep-well plates (Matrix Technologies, Hudson, NH, USA) sealed with air pore tape sheets (Qiagen, Valencia, CA, USA) and incubated with shaking for 14-16 h. Clone inserts were amplified in duplicate in 384-well format from 0.5 μl bacterial culture diluted 1:8 in sterile distilled water or from 0.5 μl purified plasmid (controls only) using 0.26 μM of each vector primer (SK865 5'-fluorescein-GTC CGT ATG TTG TGT GGA A-3' and SK536: 5'-fluorescein-GCG AAA GGG GGA TGT GCT G-3') (Integrated DNA Technologies, Coralville, IA, USA) in a 20 μl reaction consisting of 10 mM Tris-HCl pH 8.3, 3.0 mM MgCl2, 50 mM KCl, 0.2 mM each dNTP (Amersham, Piscataway, NJ, USA), 1 M betaine, and 0.50 U Taq polymerase (Roche, Indianapolis, IN, USA). Reactions were amplified with a touchdown thermal profile consisting of 94°C for 5 min; 20 cycles of 94°C for 1 min, 60°C for 1 min (minus 0.5° per cycle), 72°C for 1 min; and 15 cycles of 94°C for 5 min; 20 cycles 94°C for 1 min, 55°C for 1 min, 72°C for 1 min; terminated with a 7 min hold at 72°C. PCR reactions analyzed for single products by 1% agarose gel electrophoresis analysis. Products from replicate plates were pooled and then purified by size exclusion filtration using the Multiscreen 384 PCR filter plates (Millipore, Bedford, MA, USA) to remove unincorporated primer and PCR reaction components. Forty wells of each 384-well probe plate were quantified by the PicoGreen assay (Molecular Probes, Eugene, OR, USA) according to the manufacturer's instructions. After quantification, all plates were dried down, and reconstituted at 150 ng/μl in 3% DMSO/1.5M betaine.

a: Cell growth curves of rat PAIII prostate cancer cell line treated with 100 μg/mL soy protein isolate vs. control. b: Cell growth curves of rat PAIII prostate cancer cell line treated with genistein (100 μM and 20 μM) vs. control.

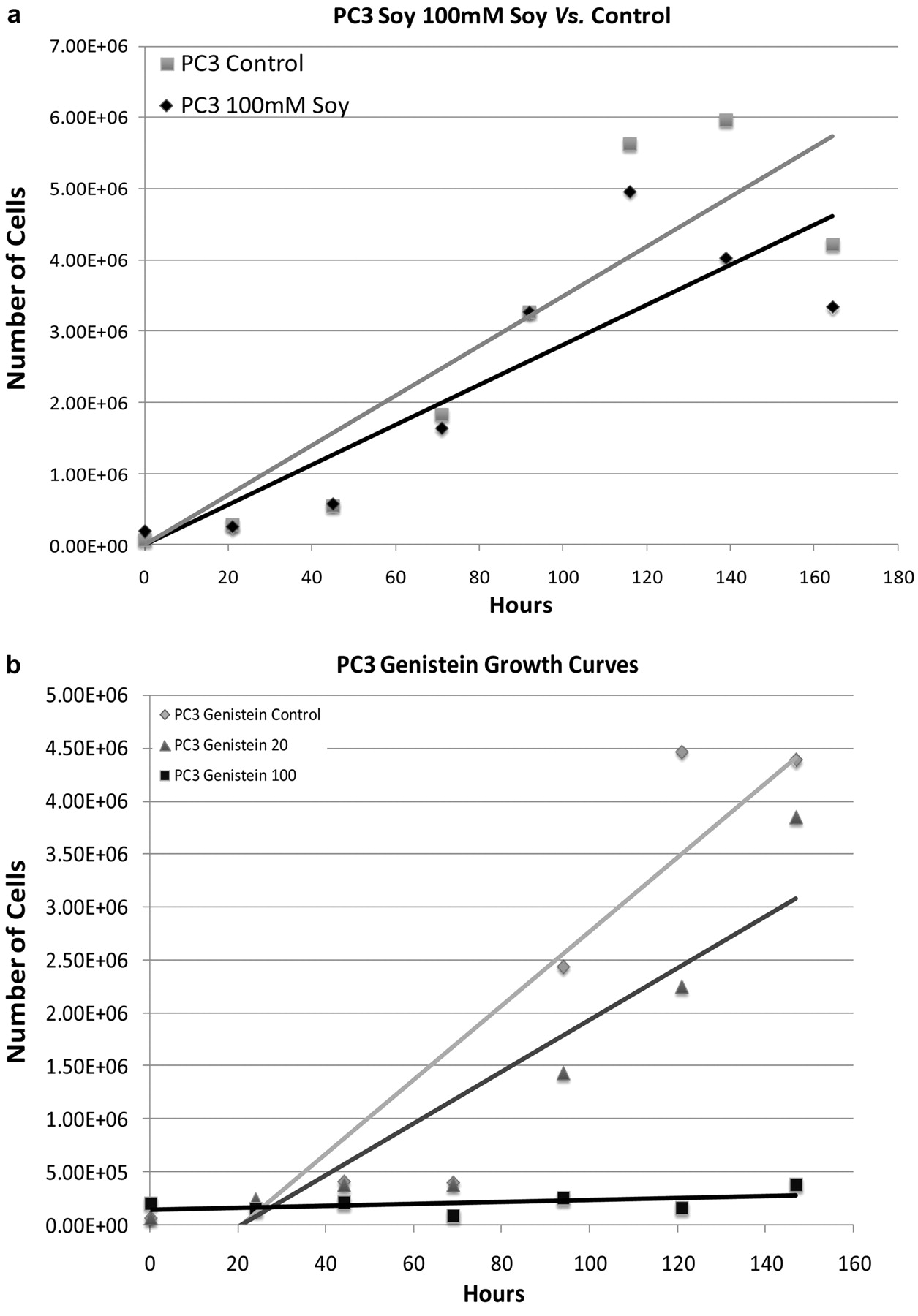
a: Cell growth curves of human PC3 prostate cancer cell line treated with 100 μg/mL soy protein isolate vs. control. b: Cell growth curves of human PC3 prostate cancer cell line treated with genistein (100 μM and 20 μM) vs. control.

a: Human LNCaP prostate cancer cell line treated with soy protein isolate vs. control, cell growth curves. b: Human LNCaP prostate cancer cell line treated with genistein (100 μM and 20 μM) vs. control, cell growth curves.

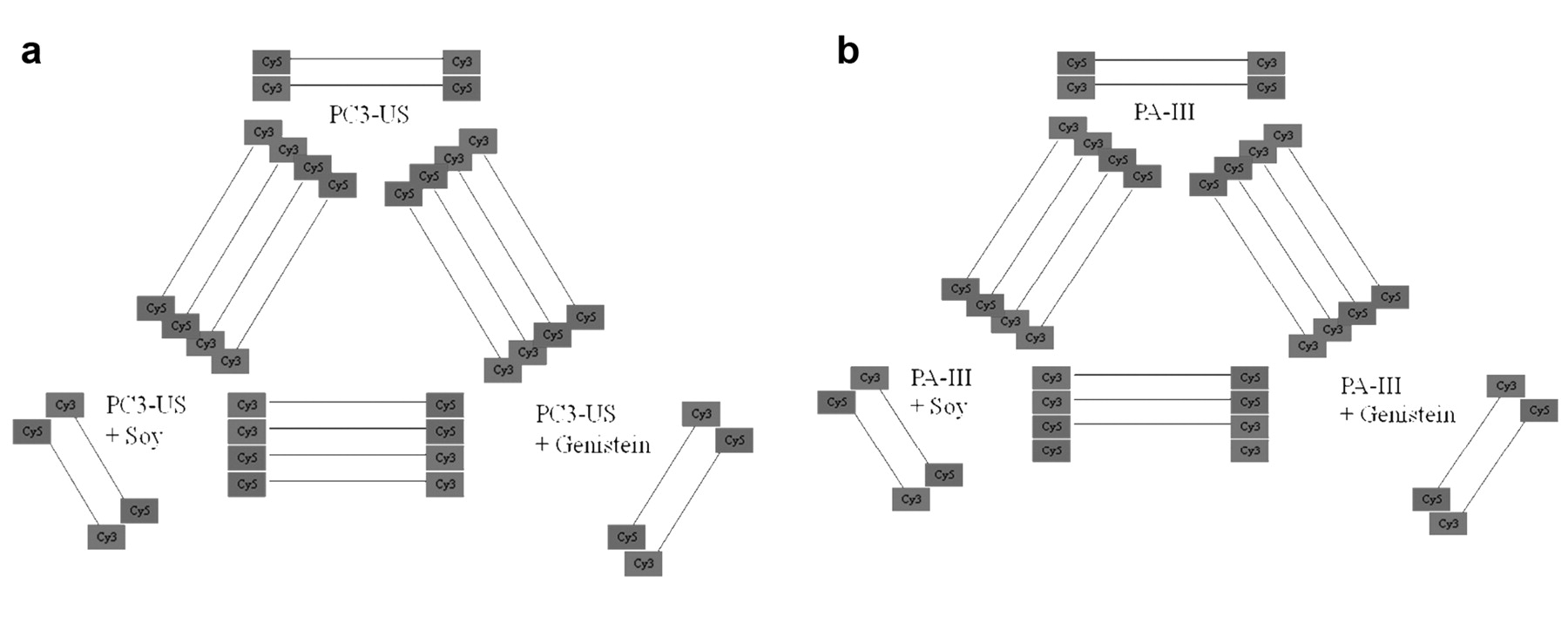
a and b: Array experimental design. Human and rat prostate cancer cell lines are compared separately under either control, soy protein isolate, or genistein treatment. Comparative studies are carried out using homologous genes on each array. Hybridizations are carried out in duplicate with dye flips. In addition, self-self hybridizations are used to establish the degree of non-specific expression changes.
Array slide fabrication. Array slide preparation was performed as previously described (14, 15). A single printing array containing 19,200 elements (human) or 2 arrays of 9,600 (rat), were printed on poly-L-lysine coated slides prepared in-house (1-2 arrays/slide) as previously described (14). Printing was conducted with a GeneMachines Omni Grid printer (San Carlos, CA, USA) with 16 or 32 Telechem International SMP3 pins (Sunnyvale, CA, USA) at 40% humidity and 22°C. To control pin contact force and duration, the instrument was set with the following Z motion parameters, velocity: 7 cm/s, acceleration: 100 cm/s2, deceleration: 100 cm/s2. All slides were post-processed using the previously described non-aqueous protocol (13). Slide coating was performed as described previously (http://cmgm.stanford.edu/pbrown/mguide/index.html). Image files on all arrays were collected after blocking (fluorescein), and again after hybridization (Cy3 and Cy5) with a ScanArray 5000 (GSI Lumonics, Billerica, MA, USA) (Figure 4a and 4b).
Experimental design and bioinformatics-based data analysis. Microarray study design and analysis was performed in methods similar to what has been previously described for selenium studies from our lab (16). The microarray study experimental design utilized two biological replicates for each comparison with each replicate incorporating a Cy3/Cy5 dye flip. In addition, self-self hybridizations were performed for each sample to ensure experimental accuracy and evaluate expression bias. Comparisons were organized in a loop design for either human or rat prostate cancer cell lines, or were run as two-sample comparisons of baseline untreated control and soy or genistein treated cells. Array image TIFF files were analyzed with Gleams software (Nutec Sciences, Atlanta, GA, USA). Additional TIFF file analysis, data normalization, clustering, and principle components analysis was performed using the Spotfinder, MIDAS and MultiExperiment Viewer Software from The Institute for Genomic Research (TIGR, Rockville, MD, USA; http://www.tigr.org/software/tm4/) and used default values set in the MCW Practical Guide to TIGR Software Use (M.Datta, unpublished). In brief, image expression data was used as channel intensity minus background and intensity thresholds were set at a value of 300. Images were analyzed as dye flip pairs normalized using MIDAS with LocFit based LOWESS normalization and slice analysis set at two standard deviation cutoffs and a sample data population of 500 (16). Samples were then averaged across two dye flip replicate pairs with removal of zero/dropped values using locally developed averaging software from the BEAR microarray suite (17). These final averaged values were subsequently annotated using the BEAR suite annotator and used for pattern identification and correlation with gene homologs. Homologous genes were identified from the NCBI homologene database ftp files (ftp://ftp.ncbi.nih.gov/pub/HomoloGene/) and parsed using local scripts and databases present in the Bioinformatics Research Center (http://brc.mcw.edu). Additional data mining to identify references in the biomedical literature associated with specific genes and user chosen search terms was performed using the locally developed GeneInfo data tool (16). Raw data files, along with analyzed data subsets are available for use and study after contacting the corresponding author.
RNA expression profiling was performed by assessing the most closely related genes between rat and human cell lines (Figures 5a and 5b) for microarray analysis. After imaging, data acquisition, normalization, and slice analysis, experimental samples were filtered based on self-self hybridization data and the final sample replicates were averaged. Genes were filtered based on desired patterns of expression (up-regulation or down-regulation at various conditions) and the results were examined. Final data mining was performed using BEAR GeneInfo with the gene names and supplemental search terms searched against PubMed to identify associated references (17). The Genebank ID's generated were matched with corresponding SwissProt ID's in order to perform GeneMap analysis of related biochemical pathways to explain the proteins growth inhibition (18, 19).
Generation of common genes and homologs. Sequence validated gene libraries for both the rat and human DNAs were obtained from Research Genetics (Huntsville, AL, USA), and were supplemented with additional DNA samples obtained from the University of Iowa rat clone sequencing program (14, 15). The majority of the rat DNAs, and a subset of the human DNAs were resequenced by Dr. J. Quackenbush at TIGR through a joint Program in Genomic Applications consortium. The GeneBank accession numbers for the 19,200 individual human or rat clones present in the recent slide printings were used to query the NCBI Unigene database to return the associated Unigene IDs. Unigene IDs were returned for virtually all identified clones, and were placed in an Oracle database where they were compared with the downloaded NCBI Homologene data set (build 106) of rat, mouse, and human homologs. Of the 19,200 clones, 5,740 genes were identified with homologues present on both the rat and human slides. This homolog set was used for the subsequent comparisons across species.

a,b: Graphing of the results demonstrated that the differentially expressed genes from human and rat cell lines fall on a straight line, indicative of a linear relationship between the expression changes (Figure 5a). A similar comparison for rat and human genes differentially expressed by genistein yielded similar results (Figure 5b).
Results
Affect on growth. Soy and genistein were used in cell culture studies to show their affect on cell proliferation. The cell lines chosen for these experiments allowed us to compare androgen-sensitive prostate cancer cells (LNCaP) and androgen-insensitive cells (PC3 and PAIII). Previously, the main effect of soy and genistein was to influence cell proliferation mainly through the androgen receptor due to the phytoestrogen component of soy and genistein (8). The effect of decreased cell proliferation has been described in other studies, yet never compared to that with whole soy protein (20, 21, 22, 23). The cell proliferation was studied by manually counting the cells and observing the growth along with the dose-related affect of the compounds.
The initial growth curve of genistein was similar across the 3 cell lines used in this study (LNCaP, PAIII, and PC3) in that the cell proliferation decreased without visual evidence of cell death. Soy, on the other hand, had the most affect at decreasing the growth of the LNCaP cell in particular (Figure 3a), while having less of an affect on the PAIII and PC3 cells (Figure 1a and 2a, respectively). The dose-related effect on cell proliferation from treatment with genistein was stronger. Genistein was used in doses of 20 mM and 100 mM to observe the influence of different concentrations on cell growth. The 100 mM dose caused the cells across all cell lines to stop proliferating without associated cell death (Figures 1b, 2b, and 3b). The 20 mM dose of genistein demonstrated a less dramatic inhibition of cell growth. For this reason in particular, we chose to use the 20 mM Genistein concentration for the RNA extraction and further study through cDNA microarray. As for soy isolate studies, the soy powder was solubilized at the highest concentration possible that did not lead to precipitate formation (saturation) which was 100 μg/mL. Overall, genistein decreased proliferation in all cell lines; whereas solubilized soy protein decreased proliferation exclusively in the LNCaP cell line.
Identification of differentially expressed genes in PC3 and PAIII prostate cancer cells treated with soy or genistein. Differential gene regulation was compared between two cells lines, one rat (PAIII) and one human (PC3). These cell lines were chosen for their similarity (non-androgen dependent) and for cross species analysis. We compared differentially expressed genes after treatment with soy in both rat and human cell lines. To confirm the genomic similarity of the responses in the two species, we compared the differential expression for homologous rat and human genes to see if they were similar. Graphing of the results demonstrated that the differentially expressed genes fell on a straight line, indicative of a linear relationship between the expression changes (Figure 5a) Thus the genes showed similar changes across species. A similar comparison for rat and human genes differentially expressed by genistein yielded similar results (Figure 5b). This technique also yielded similar results when performed in previous studies in our lab comparing Selenium response across genomic species (16). The human and rat genes from the microarray expression studies were compared with respect to soy (Table I). In this table, 10 genes that were similar in regulation among rat and human were selected due to the number of Medline listed references described in the right of Table I. The references were identified using the GeneInfo program. Five of these genes were up-regulated and 5 were down-regulated with regard to the soy to control ratios. The Medline function of the program helps to identify those genes that have been studied in the literature and helps to identify those genes whose importance in prostate cancer has been noted.
Genes that were differentially regulated in both human and rat prostate cancer cell lines treated with soy.
Genes that were differentially regulated in both human and rat prostate cancer cell lines treated with genistein.
Differentially expressed genes in rat and human cell lines with respect to genistein (Table II) showed 13 genes of interest: 7 genes were up-regulated and 6 genes were down-regulated with respect to the genistein versus control ratios. These results identified different genes from those seen with soy. The genes are displayed in summary in Table III showing overexpression of genes with respect to genistein or soy and their correlative Medline search reference lists. These are the genes that are up-regulated with treatment of soy product showing the differences in soy versus genistein. A total of 66 genes are shown in this Table, 48 genes are overexpressed with respect to genistein and 18 genes are overexpressed with respect to soy alone. Table IV separates all the differentially regulated genes only affected by soy.
Selected human genes and associated data mining. Summary table displaying overexpressed genes with respect to genistein or soy and their correlative number of Medline search hits to a specific search word. A subset of differentially regulated genes are displayed along with gene data mining results from BEAR GeneInfo, a data mining tool that allows for the whole scale search of genes with associated search terms (i.e. prostate cancer, androgen, soy). The number of references found are listed.
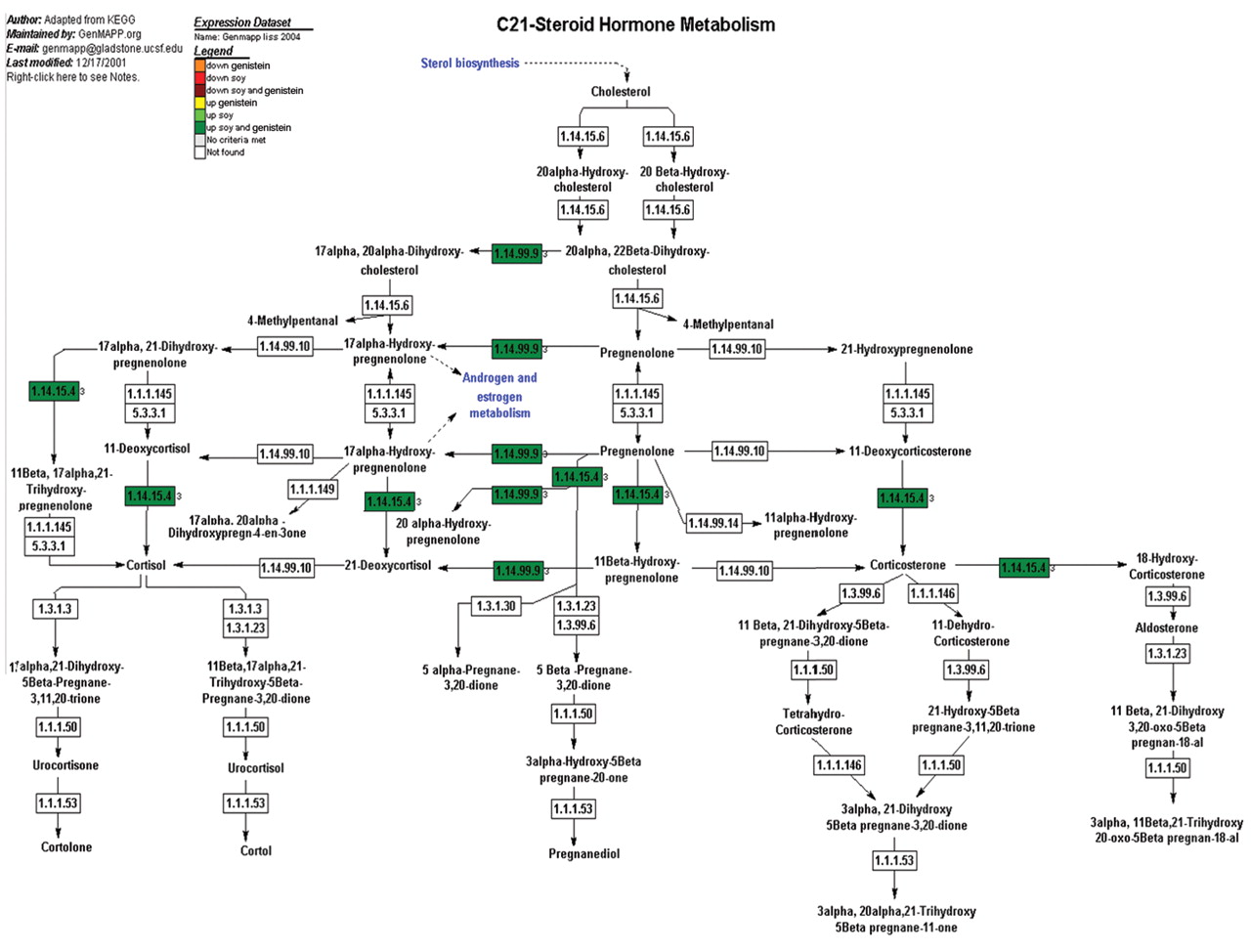
Examination of pathways affected by soy and genistein. Steroid metabolism. We used the GenMaPP analysis program and annotated cellular pathway maps from KEGG (Figure 6) to observe the influence of the phytoestrogen genistein within the steroid pathway and compare this to soy to determine any gene regulation to add to this established mechanism. Analysis of the overlaid expression data demonstrated that 12 gene were differentially regulated. All 12 genes were up-regulated with respect to both genistein and soy treatments. There were no genes regulated solely to genistein or soy.
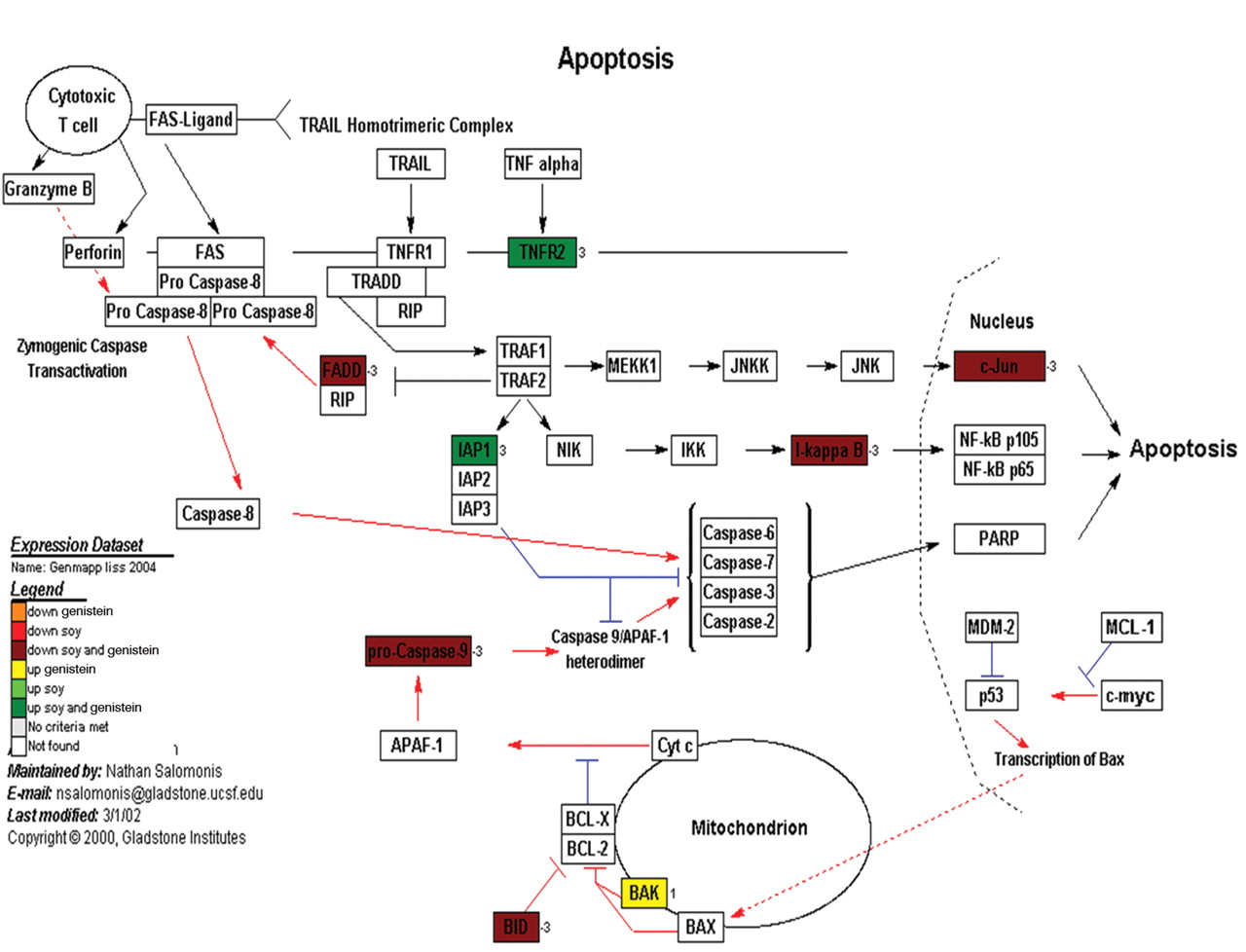
Apoptosis. As mentioned previously, apoptosis was not a major feature of the cells in cell culture exposed to the treatment of genistein or soy. The apoptosis pathway is an important regulator of cell proliferation and was explored for its importance in tumor production and proliferation. Gene expression data, with respect to known cellular pathways, again using the GenMaPP analysis program and annotated pathway maps from KEGG (Figure 7) demonstrated that 8 genes were differentially regulated in the apoptosis pathway. This includes 2 genes that were down-regulated with respect to both genistein and soy treatments and 5 genes were up-regulated. One gene was up-regulated with respect to genistein and not soy (BAK).
Human genes showing differential regulation in soy treatments, not genistein (human data only).
Cell cycle. The observed changes in cell growth curves could be caused by changes in cell cycle proteins and their regulation. For this reason we examined or gene expression data with respect to known cell cycle pathways using the GenMaPP analysis program (www.genmapp.org) and annotated cell cycle pathway maps from KEGG (Figure 8). Analysis of the overlaid expression data demonstrated that 16 cell cycle genes were differentially expressed. This included 12 genes that were down-regulated and 4 genes that were up-regulated. Of note, all genes that demonstrated differential expression were differentially expressed with both genistein and soy treatments.
Wnt signaling. Wnt signaling pathway controls the regulation of Beta-catenin, which is a well studied protein known to have nuclear effects and facilitates cell proliferation by the genes it promotes (24). As described above, the Wnt pathway has shown to be differentially regulated in prostate cancer in addition to describe beta-catenin mutations (25). In order to visualize the regulation as it pertains to soy and/or genistein, we used a KEGG file with the GenMaPP program to display the regulation patterns (Figure 9). Analysis of the overlaid expression data demonstrated that 11 genes were differentially expressed. This included 7 genes that were down-regulated and 4 genes that were up-regulated. Of note, all genes that demonstrated differential expression were differentially expressed in both genistein and soy treatments.

A representative cellular pathway map constructed by GenMaPP showing steroid hormone metabolism including colors to show the differential regulation of each gene in the pathway. The colors that correlate with the legend at the top of the Figure explain the different treatment modalities (either genistein or soy) and if the gene was up- or down-regulated.
TGF-beta. We again used the GenMaPP analysis program and annotated pathway maps from KEGG (Figure 10). Analysis of the overlaid expression data demonstrated that 11 genes were differentially expressed genes. This included 7 genes that were down-regulated and 4 genes that were up-regulated. Of note, all genes that demonstrated differential expression were differentially expressed in both genistein and soy treatments.
Nuclear receptors. The KEGG pathway maps (Figure 11) demonstrated that 9 genes were differentially expressed genes. This included 3 genes that were down-regulated and 5 genes that were up-regulated. One gene was up-regulated with respect of treatment of genistein but not soy.
Discussion
Cellular proliferation. We are interested in the clinical changes caused by soy treatment on prostate cancer development and progression. Our ability to identify changes caused by soy that correlate with clinical results of reduced tumor formation is of utmost importance. These common clinical effects have been seen in humans through epidemiologic and small early clinical studies, as well as in studies using the rat model of prostate cancer. By combining data across species and focusing on the common clinical effects and their associated gene expression changes, we hope to find specific genes that play a significant role in the clinical effects of soy on prostate cancer. Cell culture of PAIII, PC3, and LNCaP allowed visualization of growth patterns of treated soy or genistein compared to control. Genistein displayed a predictable and strong inhibitory affect on proliferation of PAIII, PC3, and LNCaP cells in cell culture (Figure 1b, 2b, 3b). The inhibiton was less dramatic when treated with soy compared to genistein. Thus it appears most likely the majority of the inhibitory affect on proliferation are due to genistein within soy. This is in agreement with the published data on genistein and the main active ingredient in soy (19, 20). While genistein is postulated to work through steroid hormone receptors, the finding of decreased cell proliferation in both androgen-sensitive LNCaP and the androgen-insensitive PC3 cells implies that the effects of genistein may occur through more than just the androgen signaling pathway. Soy protein-treated cells did not show as a dramatic inhibition on prostate cancer cells as isolated genistein, though an influence was noted. The cell proliferation of the soy treated cells showed a decreased slope; though, not significant the finding was consistent over multiple studies (Figures 1a, 2a, and 3a). The LNCaP cell line had an interesting finding in that the soy protein had a stronger affect on cell proliferation (Figure 3a). A possible explanation for this finding is that LNCaP, as an androgen-sensitive cell line, may represent an earlier, less aggressive stage of prostate cancer, and thus the component where soy may have its best chemopreventative benifit. This would suggest that soy protein acts earlier in the progression of prostate cancer and may be effective in blunting the proliferation of the tumor and slowing the course of the disease. While the main impact of both proliferation and gene expression changes appear to be the affect of genistein in the soy, other effects are identified. The differential gene expression shows that soy isolate may propagate through the genes modulated only by soy, which may affect prostate cancer cells by a mechanism other than proliferation.
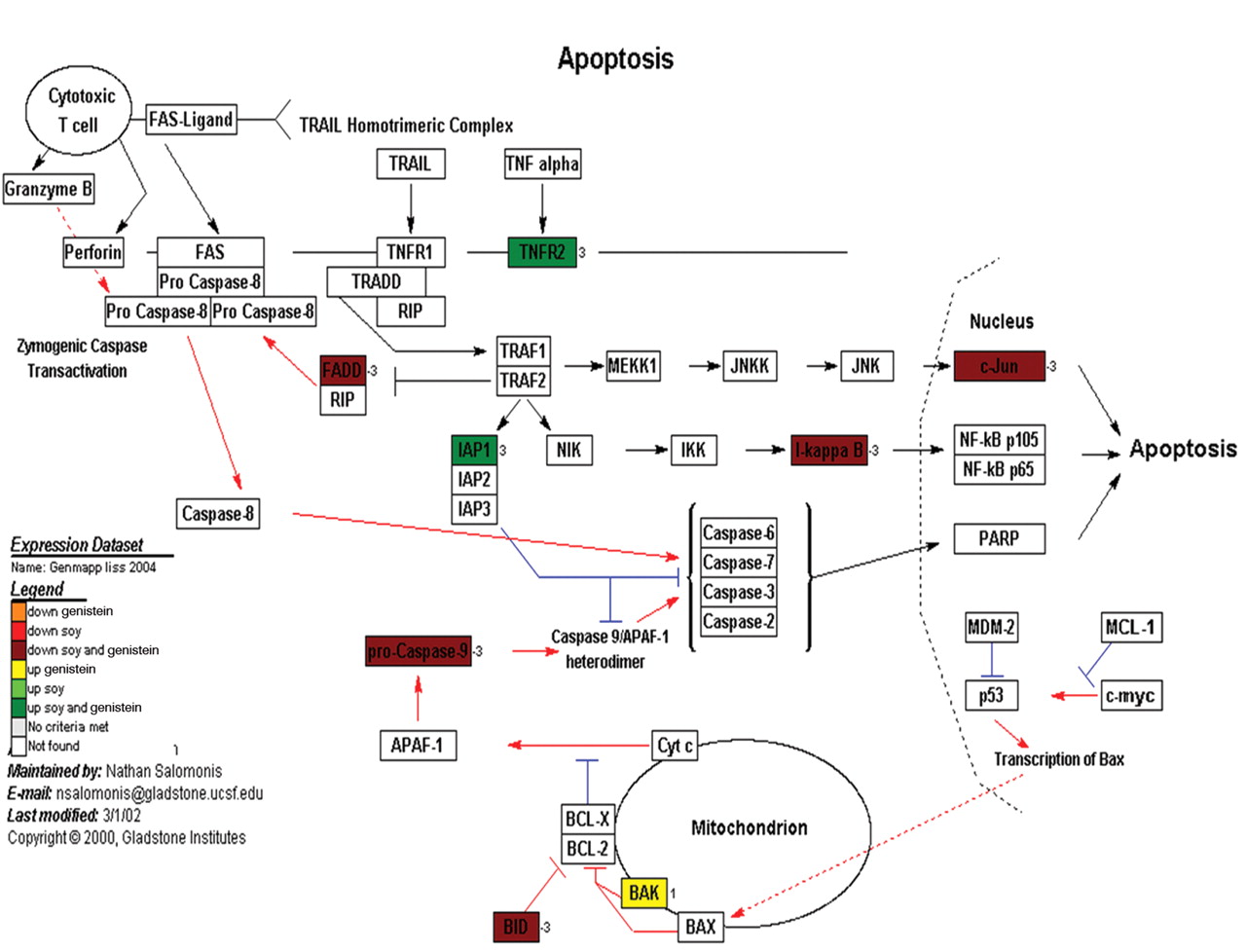
A representative cellular pathway map constructed by GenMaPP showing the apoptosis pathway including colors to show the differential regulation of each gene in the pathway. The colors that correlate with the legend at the bottom of the Figure explain the different treatment modalities (either genistein or soy) and if the gene was up or down regulated.
Examination of pathways affected by soy and genistein. Steroid metabolism. The weak oestrogenic affect of isoflavones are the most reproducible and relevant pathways that have been recognized (8, 11, 22). The interaction of steroid affects, in particular the relationship between estrogens and androgens, has been a mechanism and treatment focus for prostate cancer for over 60 years (26, 27). This association relates back to the use of orchiectomy in the early treatment of advanced prostate cancer. Hormone therapy, including medical orchiectomy through the use of steroid enzyme inhibitors and receptor blockers, is still a major treatment option for prostate cancer (28). Genistein recapitulates the affect of hormonal therapy, but on a much weaker scale and has few side-effects (23). In studying the affect of genistein on prostate cancer cells, gene microarrays have led to multiple findings that allow specific and relevant pathways to be explored. Takahashi, Lavigne, and Wang described in gene array data that the androgen receptor-mediated pathways were modulated by genistein even at the smallest concentration tested (29). They concluded that the androgen receptor mediated pathways were the most relevant mechanism to target for genistein studies. Betick et al. also described that the androgen receptors were down-regulated in LNCaP cells in response to physiologic treatments of genistein (30).
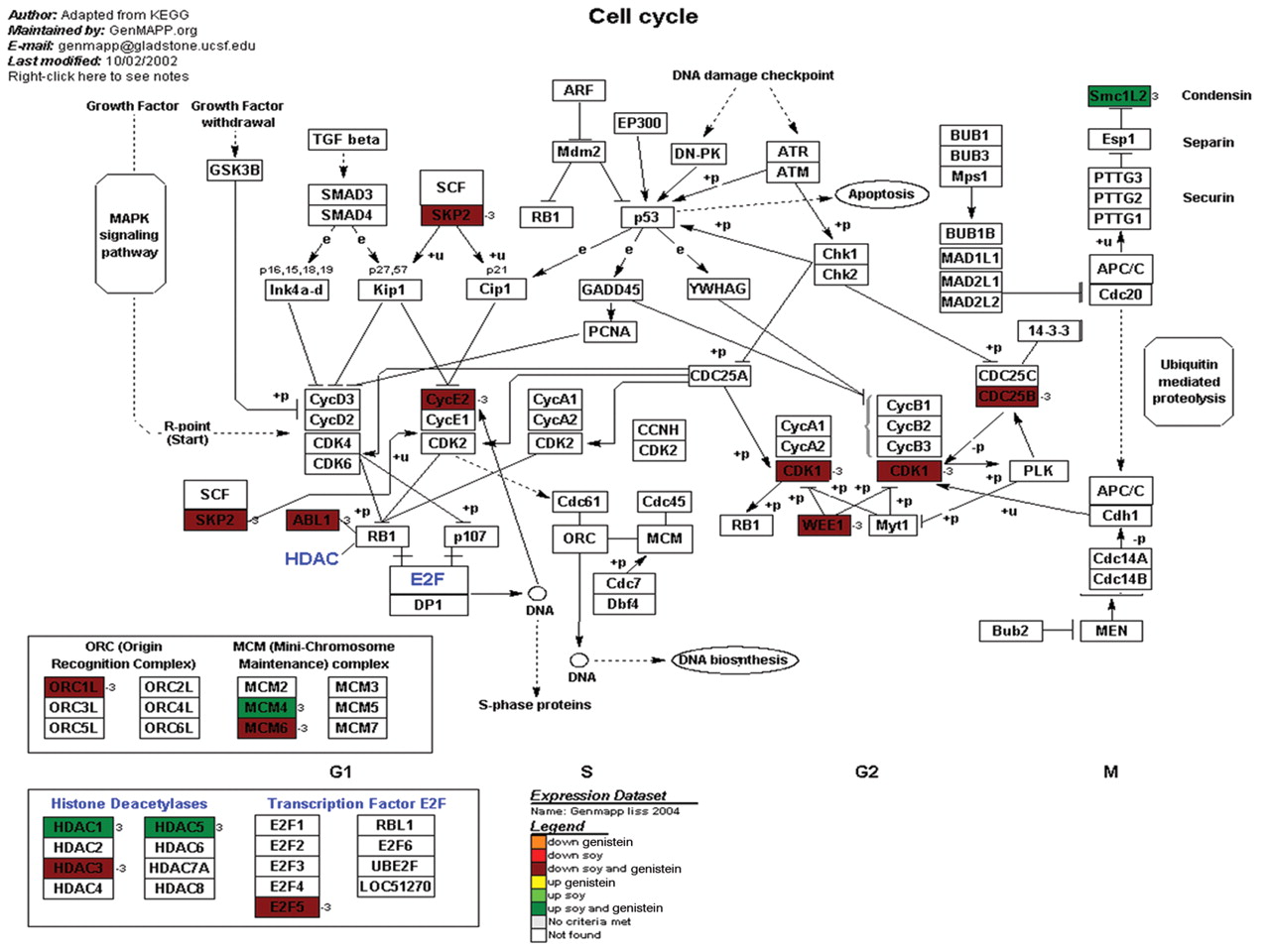
A representative cellular pathway map constructed by GenMaPP showing the cell cycle pathway including colors to show the differential regulation of each gene in the pathway. The colors that correlate with the legend at the bottom of the Figure explain the different treatment modalities (either genistein or soy) and if the gene was up- or down-regulated.
Due to these previous studies describing the effect of genistein on the steroid hormone metabolism pathway, we wanted to compare genistein and soy isolate for this pathway as well. Steroid metabolism is an important aspect of prostate cancer that may impact tumor aggressiveness and prognosis. The GenMapp pathways from this study shows that the vast majority of gene regulation changes express upreguation using soy or genistein (Figure 6). In particular, the majority of the changes in the metabolic enzymes are of up-regulation. The genistein/soy based inhibition of testosterone and DHT could cause feedback expression increases in the enzymes responsible for testosterone production. This hypothesis supports one of the most prominent models by which soy is thought to exert its chemopreventive effects, through androgen pathway inhibition (29, 31).
Apoptosis. Steroid metabolism plays a very intricate and vital role in the overall affect of soy isolates, yet the direct cytotoxic affect has only recently been evaluated. One mechanism of involves the role of proteasomes in apoptosis regulation (32). Preliminary data from Kazi et al. and others found that in the LNCaP cell line, genistein-mediated proteasome inhibition was accompanied by induction of apoptosis (8, 32). Genistein has also been implicated in the induction of caspases, in particular caspase-3, to mediate apoptosis via CPP32 (33). Another manuscript also noted that genistein may induce apoptosis through topoisomerase II (8). In addition, Davis, Singh, and Bhuiyan. described the affect of genistein on cellular proliferation and subsequent apoptosis. In their studies genistein results in the down-regulation of cyclin B and up-regulation in p21 inducing apoptosis after placing the cells in cycle arrest (34). Some studies suggest genistein does not necessarily cause apoptosis, but sensitizes the cancer cells to inhance chemotherapeutic agents (35). The effects of genistein may be dose-dependent because when comparing different concentrations of genistein, higher doses may cause apoptosis while lower doses may not.
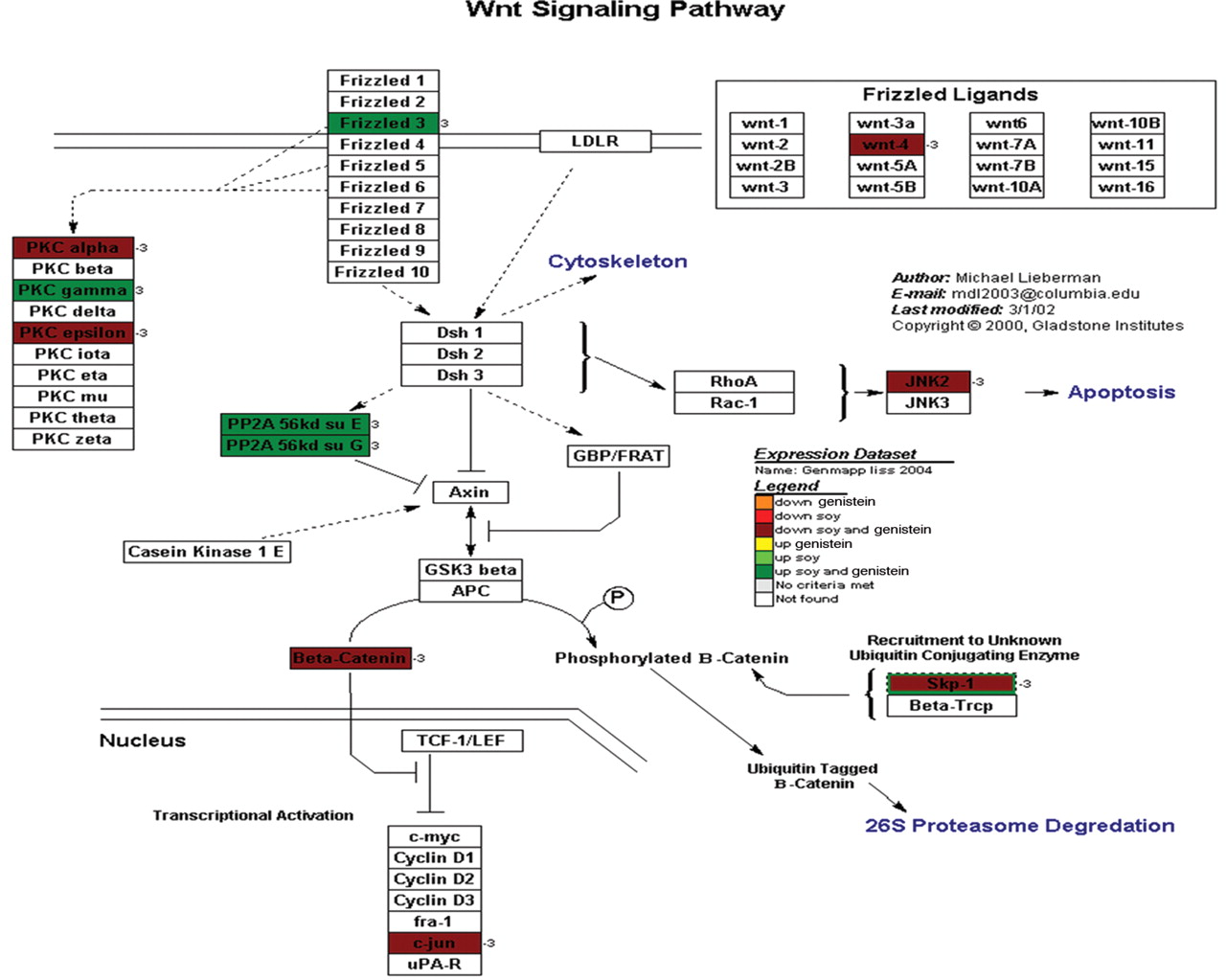
A representative cellular pathway map constructed by GenMaPP showing the Wnt signaling pathway including colors to show the differential regulation of each gene in the pathway. The colors that correlate with the legend explain the different treatment modalities (either genistein or soy) and if the gene was up- or down-regulated.
We examined soy and genistein differential expression data in PC3 and PAIII and displayed them in a GenMapp KEGG Figure (Figure 7). The majority of the apoptotic pathway components are down-regulated by both soy and genistein and was demonstrated in the cell culture where the treated cells were viable but not proliferating, possibly in G0 arrest. This suggested that the decrease in proliferation was an intrinsic function of the cells and not cell death. Thus while apoptosis is not enhanced with the use of soy or genistein alone, it may predispose the cells to G0 arrest and subsequent apoptosis by other means.
Cell cycle. Hormonal regulation by genistein also plays a role in cell cycle modulation by decreasing cellular proliferation. Wang, Clubbs, and Bomser showed that genistein alters cell proliferation via estrogen dependent kinases as well as extracellular kinases (22). Therefore, they added to the understanding that genistein has both hormonal-dependent and -independent mechanisms affecting the prostate tumor proliferation. Another venue in which soy protein and its isolates may affect cells other than androgens and direct cytotoxicity is modulation of the cell cycle by acting on the cyclins. Soy isolates can alter the cell cycle by modulating TGF-1β or cyclins directly (34, 36). The cell cycle shows diffuse down-regulation in cells when treated with soy or genistein (Figure 8). These results have also been shown in tissue culture and confirms that soy and genistein down-regulate the cell cycle and proliferation of prostate cancer cells.

A representative cellular pathway map constructed by GenMaPP showing the TGF-beta signaling pathway including colors to show the differential regulation of each gene in the pathway. The colors that correlate with the legend at the bottom of the Figure explain the different treatment modalities (either genistein or soy) and if the gene was up- or down-regulated.
WNT signaling. Wnts and growth factors have been shown to be differentially expressed in normal prostate tissue and prostate cancer (24). Various Wnt are up-regulated in cancer and have been associated with higher Gleason scores (37). Wnt inhibitory factors are frequently down-regulated possibly causing decreased active GSK3β and increased soluble β-catenin. This leads to cell proliferation in PC3 cells (38). GSK3 is a protein that will phosphorylate β-catenin, therefore, targeting the β-catenin protein for ubiqination and destruction. GSK3 inhibition leads to increased β-catenin concentration and transport into the nucleus activating genes associated with cell proliferation (24). β-Catenin is indirectly associated with the Akt pathway via GSK3 and is also involved in IGF-1-induced androgen receptor activation, thus linking androgens and the Wnt pathway (39). β-Catenin has also been implicated in prostate cancer due to various mutations in the gene noted in prostate cancer (40). These mutations may block β-catenin protein phosphorylation and ubiquitin-related destruction, leading to continued cell proliferation (25). These mutations in β-catenin have been shown in a limited amount of prostate cancer samples and may be a late finding in cancer progression. Verras and Sun concluded that 20-38 % of hormone refractory prostate cancers show β-catenin nuclear localization greater than that would be caused by β-catenin mutation alone, indicating the Wnt pathway as a possible source of this β-catenin activation (40). Soluble β-catenin is only one of the sources of β-catenin, which also has a structural cell function. This other pool of β-catenin allows the stabilization of E-cadherin and thus may be involved in tumor cell metastatic potential (30). Figure 9 shows the GeneMaPP displaying β-catenin to be down-regulated by both soy and genistein. The frizzled receptor is up-regulated by both. The up-regulation of the frizzled 3 protein is likely a feedback mechanism to increase the frizzled receptors due to down-regulation of β-catenin and decreased cell proliferation. Of note, the JNK protein, which is important in the apoptotic pathway, is down-regulated as well. This again suggests that the decrease in proliferation was an intrinsic cellular mechanism not due to apoptosis.
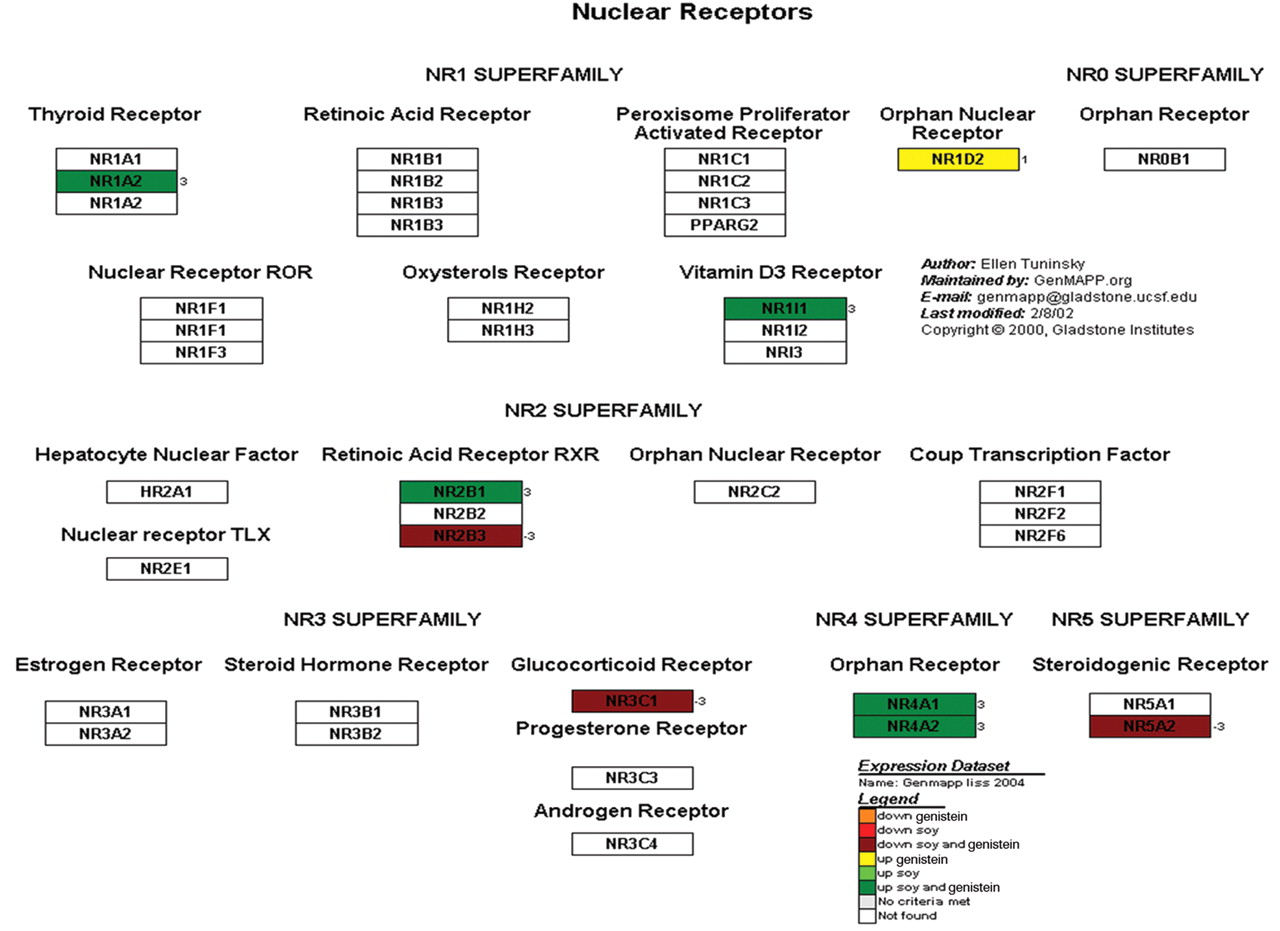
A representative cellular pathway map constructed by GenMaPP showing nuclear receptors including colors to show the differential regulation of each gene in the pathway. The colors that correlate with the legend at the bottom of the Figure explain the different treatment modalities (either genistein or soy) and if the gene was up- or down-regulated.
TGF-beta. The TGF-β signaling pathway plays a central role in epithelial cell growth and acts through multiple intracellular pathways. Upon receptor binding, Smad2/3 proteins become phosphorylated and associates with Smad4 (41, 42). This Smad complex translocates to the nucleus, binds to DNA and regulates transcription of specific genes (41). The TGF-β pathway is interconnected with the androgen receptors, VEGF, pathway, JNK/JUN pathway and Wnt pathway through β-catenin and other cofactors including the Smads proteins. Recent data from Sheehan Kallakury. and Ross described that smad4 expression correlates with grade and stage of prostate cancer (42). We have found that the majority of the the TGF-β signaling pathway are inhibited by soy and genistein. Smad 2 in particular is down-regulated with the inhibitory Smad6 being up-regulated (Figure 10). These findings describe that inhibition of the TGF-β pathway correlates with inhibition of cellular proliferation suggesting the value of further researched in this mechanism.
Nuclear receptors. Nuclear receptors play and intricate part in guiding cell proliferation and production as the result of a changing environment. By studying these receptors, insight into the action of soy and/or genistein on nuclear factors and how they regulate cell function is of great importance to explain mechanism of action. Figure 11 contains some important nuclear receptors and their function in cell signaling and proliferation. Of note, the vitamin D3 nuclear receptor is up-regulated with soy and genistein. Tokar and Weber have recently described that vitamin D3 (cholecalciferol) inhibits cellular changes associated with malignant transformation and invasion of prostatic neoplasia by up-regulating nuclear receptors (43). The connection between vitamin D and androgen receptors is controversial but may suggest an important connection between soy and vitamin D. In our study, the orphan nuclear receptor NR1D2 is the only nuclear receptor solely influenced by genistein and not soy (Figure 11). The NR1D2 receptor is up-regulated by genistein and is linked to the RARB gene, which encodes retinoic acid receptor beta (44). The retinoic acid receptors are also differentially regulated by soy and genistein. Of note the NR2B1 orphan nuclear receptor is complexed in a herterodimer with constitutive androstane receptor (CAR) causing it to be continually expressed (45). Figure 11 shows soy and genistein up-regulate this CAR/RXR heterodimer causing the retinoic acid signaling mechanism to be continually expressed, possibly decreasing cancer growth.
Conclusion
Soy protein has been shown to decrease the incidence of clinically significant prostate cancer in both human studies and in animal models of prostate cancer. We have used gene expression microarrays and cross-species gene analysis to identify gene pathways that may be influenced by soy protein.. While the majority of the features we found regarding pathway regulation were common to soy and its purified factor genistein, some genes were identified via microarray analysis possibly due to other soy protein components. The soy protein isolate is made of many proteins, in which only a few have been investigated, and more importantly it is how these proteins interact with one another that is more difficult to investigate. The use of soy and genistein play an important role in chemoprevention and pharmacology in designing treatment modalities to reduce the risk for prostate cancer.
Acknowledgments
NCI funding through Julia White, MD in the Department of Radiation Oncology at the Medical College of Wisconsin, Milwaukee, WI, USA, who had no involvement in the study design, in the collection, analysis and interpretation of data, in the writing of the manuscript, or in the decision to submit the manuscript for publication.
Mark Suckow and Morris Pollard at the Lobund Laboratory at the University of Notre Dame for supplying the PA-III rat cell line and the soy protein isolate for investigation. Paul Lindhol and Andre Kadjacsy-Balla for supplying the PC-3 human cell line.
- Received March 1, 2010.
- Accepted March 12, 2010.
- Copyright© 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.